DOI 10.17480/psk.2018.62.2.118
인간 기도 상피세포에서 MUC5AC 뮤신 유전자 발현 및 생성에 대한 코니페린 등 흰털오가피 유래 천연물의 영향
이 현 재*
삼육대학교 교양대학 및 대학원 중독과학과
(Received March 18, 2018; Revised April 18, 2018; Accepted April 19, 2018)
Effects of natural products derived from Acanthopanax divaricatus var. albeofructus including coniferin on gene expression and production of MUC5AC
mucin from human airway epithelial cells
Hyun Jae Lee*
Smith Liberal Arts College and Department of Addiction Science, Graduate School, Sahmyook University, 815 Hwarang-ro, Nowon-gu, Seoul 01795, Korea
Abstract — We investigated whether coniferin, tortoside A, eleutheroside E or salvadoraside derived from Acanthopanax divaricatus var. albeofructus affects airway MUC5AC mucin gene expression and production induced by epidermal growth factor(EGF) from NCI-H292 cells. Cells were pretreated with each agent for 30 min and then stimulated with EGF for 24h. The four compounds inhibited the gene expression and production of MUC5AC mucin from NCI-H292 cells. This result suggests that the four compounds can inhibit the production and gene expression of mucin induced by EGF, through directly acting on airway epithelial cells.
Keywords airway, mucin, coniferin)
서론 (Introduction)
인간의 기도 내부에 존재하는 점액(mucus)은 상피세포층을 덮 고 있으며 점액섬모 청소(mucociliary clearance) 기능을 통하여 비산 화학물질, 미세분진 입자, 병원성 미생물 등 인체 외부에서 유입되는 유해인자들에 대한 생리적 방어작용을 수행함으로써 기도 내부를 독성 자극으로부터 보호하고 있다. 호흡기에 존재 하는 점액의 이와 같은 인체 방어작용은 점액의 생화학적 주 구 성요소인 뮤신의 물리화학적 특성인 점탄성(viscoelasticity)에 기 인한다. 그러나, 정상 범위를 벗어난 뮤신의 양과 질의 이상은 인
체의 방어작용에 영향을 주어 심각한 병리 현상을 유발할 수 있 다. 즉, 천식, 만성 기관지염, 폐기종, 기관지 확장증, 낭포성 섬 유증 등의 기도 질환에서 관찰되는 점액(객담)의 과다 생성 및 분비는 이러한 질환의 예후를 악화시키는 주된 요인으로 알려져 있다.1-4)과다 생성 및 분비된 점액을 기도로부터 제거하는 데는 두 가지 방법이 있을 수 있다. 첫째, 물리적 방법에 의한 점액의 제거, 즉 점액의 점도를 낮춘 뒤 흡인해 내는 방법이고, 둘째는, 점액 생성 자체를 억제할 수 있는 약물을 투여하는 방법이다. 물 리적 방법은 기도 내부의 자극을 유발하고, 반사기전에 의해 점 액 분비를 오히려 자극하게 된다. 마취 하에서 그런 방법이 시도 된다고 해도, 점액의 제거는 feedback mechanism을 통해 점액 의 생성과 분비를 더욱더 자극하게 된다. 따라서, 점액에 점성을 부여하는 주 구성요소인 뮤신의 생성 자체를 조절하거나 혹은 분 비를 조절하기 위한 약물학적 접근은 호흡기 질환의 치료에 있 어 중요한 전략이 될 수 있다.5)이러한 전략에 근거하여, 항염증, 항산화, 항암 효능을 발현하는 것으로 알려진 다수의 천연물 중에
#
Corresponding Author Hyun Jae Lee
Smith Liberal Arts College and Department of Addiction Sci- ence, Graduate School Sahmyook University, 815 Hwarang-ro, Nowon-gu, Seoul 01795, Korea
Tel.: 010-2506-1297 Fax.: 02-3399-1909 E-mail: [email protected]
Short Report
종설서 호흡기 염증성 질환에서 관찰되는 기도 뮤신의 생성 및 과다분 비를 조절할 가능성이 있는 후보물질의 탐색은 유효 적절한 접 근 방법이라 할 수 있을 것이다. 전통의학 문헌과 다수의 연구보 고에 의하면, 오가피(Acanthopanax divaricatus var. albeofructus) 는 경험적으로 강심, 강장, 거풍습 효능을 포함하여 다양한 염증 성 질환의 조절을 위해 사용되어 왔으며,6)오가피에 함유된 코 니페린, 토르토사이드 A, 엘류쎄로사이드 E등의 천연물이 항염 증, 항스트레스 등의 생리활성을 발현함이 알려져 있다. 즉, 코 니페린 등의 phenylpropanoid glycosides와 토르토사이드 A와 같 은 리그난 등은 항염증 작용을 나타내며, 엘류쎄로사이드 E 는 관 절염 등의 병리적 상황에서 조절 작용을 발현하고, 설치류의 수 면박탈에 의한 스트레스 모델에서 항스트레스 작용을 나타냄이 보고되어 있다.7-10)그러나, 현재까지 인간 기도 상피세포에서 뮤 신 유전자의 발현 및 생성에 대해 오가피에서 유래한 코니페린, 토르토사이드 A, 엘류쎄로사이드 E, 살바도라사이드 등의 천연 물이 어떠한 영향을 미치는 지 여부는 연구된 바가 없었다. 따라 서, 본 연구에서는 코니페린, 토르토사이드 A, 엘류쎄로사이드 E, 살바도라사이드 등이 인간 기도 상피세포에서 상피세포 성장 인자인 EGF에 의해 증가된 MUC5AC 뮤신의 유전자 발현 및 생성에 대해 어떠한 영향을 나타내는 지 탐색함으로써, 후속 연 구를 통한 효과적인 기도점액 과다생성(분비) 조절 신약의 개발 을 위한 단초를 제시하고자 하였다.
실험 방법 (Experimental Methods)
세포주 및 시약
NCI-H292 세포는 American Type Culture Collection 사 (Manassas, VA, U.S.A.)에서 구입하였다. 생약 전문가에 의해 감 별 완료된 흰털오가피 수피 6 kg을 잘게 절단한 후 MeOH (10 L) 을 이용하여 3회 반복 환류 추출한 후 감압 농축하여 MeOH 추 출물 200 g을 얻었다. MeOH 추출물을 물에 현탁시킨 후 hexane (0.8 L X 3)으로 분획하고, 농축, 건조하여 hexane 분획 추출물 (79.0 g)을 얻었다. 잔류물을 농축, 건조시켜 다시 증류수에 현탁 시킨 후 ethyl acetate(0.8 L X 3)를 이용하여 ethyl acetate 분 획(36.0 g)과 물 분획(50.0 g)을 얻었다. Ethyl acetate 분획과 물 분획을 다시 각종 컬럼 크로마토그래피를 수행하여 coniferin, totrtoside A, eleutheroside E, salvadoraside 등의 물질을 분리, 정제하였는데, 제반 과정은 충남대학교 약학대학 생약학교실 김 영호 교수 연구실에서 수행되었다(Fig. 1). Protease inhibitor cocktail은 Roche사(Indianapolis, IN, U.S.A.)에서, mouse anti- MUC5AC clone 45M1 및 HRP-Goat Anti-Mouse IgG Conjugate 은 NeoMarkers사(Freemont, CA, U.S.A.)에서, trypsin-EDTA, epidermal growth factor(EGF), Tween 20, bovine serum albumin(BSA), HEPES, dimethyl sulfoxide(DMSO), 3,3',5,5'-
tetramethyl- benzidine peroxide solution(TMB), NP-40, EDTA, EGTA, HEPES, ethidium bromide, diethylpyrocarbonate(DEPC), 등은 Sigma-aldrich(St. Louis, MO, U.S.A.)사에서, Easy-Blue RNA extraction kit는 INTRON biotechnology(Kyunggi-do, Korea) 사에서, Accuprep RT premix kit는 Bioneer(Daejeon, Korea) 사에서, 그리고 PCR Master Mix는 ABgene(Rochester, NY, U.S.A.) 사에서, penicillin-G, streptomycin, fetal bovine serum(FBS), RPMI 1640은 GIBCO-BRL(Grand Island, New York, U.S.A.) 사에서 구입하여 사용하였다.
인간 기도상피 세포(NCI-H292) 배양 및 약물 처리
세포는 습도가 충분히 유지되며 95% 공기, 5% CO2를 함유하
Fig. 1 − Chemical structure of coniferin, totrtoside A, eleutheroside
E and salvadoraside.
는 37oC 배양기 내에서 HEPES(25 mM), penicillin G(100 U/
ml), streptomycin(100μg/ml), FBS(10%, V/V) 등이 첨가된 RPMI 1640 배양액에서 배양하였는데, 1주에 2회 빈도로 subculture하였고 배양액은 2일마다 1회씩 교체하여 주었다. 뮤 신 생성 및 그 유전자 발현에 대한 약물의 작용을 검증하기 위 하여, 뮤신 생성량 검증을 위해서는 24 well culture plate 를 기 준으로, well 당 2.0 × 104 cells/well의 밀도로, 뮤신 유전자 발현 정도의 검증을 위해서는 6 well culture plate 를 기준으로, well 당 5.0 × 104 cells/well의 밀도로 각각 세포를 도포하고 배양하였 다. 세포가 각 well의 70-80% 정도를 차지할 정도로 자라면, FBS 의 농도를 0.2%로 감소시킨 배양액을 주고 24 시간 동안 배양 하고, serum을 첨가하지 않은 배양액(serum-free medium) 으로 세포를 세척한 후 약물 1-100 μM을 함유하는 배양액 200 μl(24 well plate 기준)를 well마다 가하였다. 30분이 경과한 후 EGF (25 ng/ml)를 세포에 투여한 후 37oC에서 추가로 24시간 동안 배 양하였다.11-13)
인간 기도상피 세포 내에 존재하는 total RNA의 분리 24 시간 동안 각 약물을 처리한 세포를 냉각된 PBS 로 2회 세척하였다. 세포에 trypsin-EDTA 용액을 처리하여 배양 용기 바닥으로부터 분리하고, 세포들의 혼합물을 1.5 ml 용량의 microtube에 옮겨 원심 분리함으로써 세포들만 수거하였다. 이 어서, total RNA를 분리하고자 INTRON biotechnology 사의 Easy-Blue RNA extraction kit(total RNA isolation reagent) 를 이용해(0.5 ml/4 × 105 cells) 세포를 lysis 시키고, 상온에서 5 분간 방치하였다. 5분 후 즉시, microtube 에 chloroform을 첨가, 15초간 vortexing 하고 상온에 2-3분간 방치한 후 4oC, 13,000 rpm(Hanil centrifuge, MICRO 17 R)에서 10분간 원심 분리하여 얻은 상층액 400 μl를 새 microtube에 옮겼다. 상층액 에 동량의 isopropanol을 첨가하여 잘 혼합한 후 상온에서 10분 간 방치하고 다시 4oC, 13,000 rpm에서 10분간 원심 분리하여 RNA 침전물을 얻었다. 이 침전물에 diethylpyrocarbonate(DEPC) 가 함유된 75% ethanol 을 가하고 4oC, 10,000 rpm에서 10분간 원심 분리함으로써 세척하였다. 수거된 RNA 침전물을 5분간 대 기 중에서 건조시킨 후, 20 μl 의 RNase-free water로 부유시키 고, spectrophotometer(Beckman, DU-650)를 사용하여 260 nm 파장에서 흡광도를 측정함으로써 RNA 의 농도를 계량한 후 실 험에 사용하였다.14)
PCR(Polymerase Chain Reaction)을 위한 primer 제조 PCR에 사용된 primer는 전문 제조회사인 Genotec(주) (Daejeon, Korea) 에 주문, 합성하였다. NCI-H292 세포에서의 human MUC5AC 유전자 합성을 위해 사용한 sense primer 의 염기서열은 5‘-TGA TCA TCC AGC AGC AGG GCT-3',
antisense primer 의 염기서열은 5’-CCG AGC TCA GAG GAC ATA TGG G-3' 이었다. 정량적 대조 유전자로 사용된 Rig/S15 유전자 primer의 염기서열은 (sense primer) 5'-TTC CGC AAG TTC ACC TAC C-3' 및 (antisense primer) 5'-CGG GCC GGC CAT GCT TTA CG-3'이었다.
RNA의 역전사 반응 및 중합효소 연쇄반응(RT-PCR) 수거된 total RNA를 이용, 역전사 반응(Reverse Transcription) 으로 cDNA 를 만들고, 이를 중합효소 연쇄반응(PCR) 으로 증폭 시켰다. 즉, 얻어진 total RNA 1 μg을 75oC에서 5분간 가열함으 로써 denaturation 시키고, 이를 얼음에 담가 급냉시킨 후 RT premix kit의 사용자 설명서에 따라 역전사 반응을 진행시켰다.
MUC5AC 유전자에 대한 PCR 은, 각각의 역전사 반응에서 얻 은 cDNA 산물 2 μl를 PCR premix kit 의 사용자 설명서에 따 라 진행시켰다. 증폭반응을 위하여, PCR을 40회 실시(PCR thermal cycler, Takara MP-300, Japan) 하였으며, denaturation 은 94oC에서 30초, annealing은 60oC에서 30초, extension은 72oC에서 30초간 각각 시행하였다.
전기영동에 의한 중합효소 연쇄반응 산물의 확인
RNA의 역전사 반응 및 중합효소 연쇄반응으로 증폭된 cDNA 산물들을 전기영동으로 분리함으로써 MUC5AC 유전자 발현 변 동여부를 관찰하였다. 즉, 증폭된 PCR 산물 10 μl를 10×gel loading buffer(0.25% bromphenol blue, 0.25% xylene cyanol FF, 50% glycerol)와 잘 혼합한 다음, Tris-acetate-EDTA buffer (40 mM Tris-acetate, 1 mM EDTA) 용액 및 1 μg/ml의 ethidium bromide가 포함된 1.0% agarose gel 에서 전기 영동하였다. Gel 상에서 이동된 각각의 DNA band 는 자외선 투사기(ultraviolet transilluminator)를 이용하여 관찰하고, 사진 촬영하였다.
인간 기도상피 세포에서의 MUC5AC 뮤신 생성량 측정 각 약물의 처리 기간이 종료된 후, 배상세포 내 생성되어 저장 되어 있는 뮤신을 정량하기 위하여, 세포 용해용 완충액(20 mM Tris, 0.5% NP-40, 250 mM NaCl, 3 mM EDTA, 3 mM EGTA, protease inhibitor cocktail)을 가하여 세포 내에 존재하는 MUC5AC 뮤신을 추출하였다. 즉시로, 효소연계 면역흡착법 (enzyme-linked immunosorbent assay, ELISA) 을 이용하여 뮤 신의 생성량을 다음과 같이 측정하였다. 뮤신을 함유하고 있는 배양 상층액 및 수거된 cell lysate를 각각 PBS로 1/10배 희석하 고, 희석된 각 sample을 ELISA 전용의 96-well plate에 각각 100μl씩 분포시킨 후 42oC에서 완전히 건조시켰다. 그 후 PBS- Tween 20(0.05%, PBS-T) 용액 200 μl/well을 이용, 각 well 당 3회씩 세척하였다. 세척 후 PBS-T에 용해된 2% BSA 용액 200μl를 각 well당 가하고 다시 1시간 동안 incubation하였다. 1
시간 후 PBS-T 200 μl로 3회 세척하고 MUC5AC에 대한 monoclonal antibody인 mouse anti-MUC5AC clone 45M1을 2% BSA에 1 : 200의 비율로 희석한 후에, 각 well당 100 μl씩 첨가하고1시간 동안 incubation하였다. 1시간 후 PBS-T로 3회 세척하고 2차 항체인 Horse radish peroxidase(HRP)-Goat Anti-Mouse IgG Conjugate를 2% BSA에 1 : 3,000의 비율로 희석한 후, 각 well당 100 μl씩 첨가하고 1시간 동안 incubation 하였다. PBS-T로 다시 3회 세척 후 3,3',5,5'- tetramethyl- benzidine peroxide(TMB) 용액 100 μl를 각 well에 첨가하고 5 분 후 1N H2SO4 50lμl를 첨가하여 반응을 정지시켰다. 450 nm 에서 각 well의 흡광도를 측정함으로써 대조군과 약물 처리군에 존재하는 MUC5AC 뮤신을 정량하였다.11-13)
통계처리
모든 측정 결과는 Mean±S.E.M. 으로 환산한 후, 약물 처리군 의 측정치는 대조군 측정치의 백분율로 나타냈다. 통계처리는 one- way ANOVA 및 post-hoc test 로서 Holm-Sidak test를 이용하였 으며 p<0.05인 경우 통계적으로 유의한 것으로 판정하였다.
실험 결과 (Results)
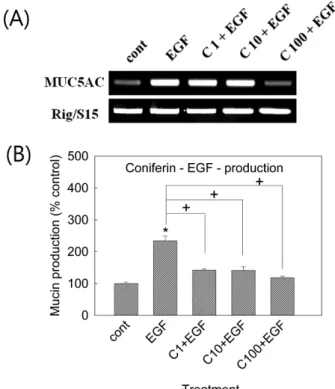
코니페린이 EGF로 자극된 MUC5AC 기도 뮤신 유전자 발현 과 뮤신의 생성에 미치는 영향
코니페린은 EGF처리로 증가된 MUC5AC 유전자 발현을 억 제하는 경향을 보여주었다. 동시에, 코니페린은 25 ng/ml의 EGF 처리로 증가된 MUC5AC 기도뮤신의 생성을 억제하였다. 각 처 리 농도별 뮤신의 양은, 대조군, EGF 25 ng/ml단독 처리군, 코니 페린 10-6 M + EGF, 코니페린 10-5 M + EGF, 코니페린 10-4 M + EGF처리군에서 각각 100±4%, 234±15%, 142±4%, 141±
11%, 118±5% 이었다(Fig. 2(A), (B)).
토르토사이드 A가 EGF로 자극된 MUC5AC 기도 뮤신 유전 자 발현과 뮤신의 생성에 미치는 영향
토르토사이드 A는 EGF처리로 증가된 MUC5AC 유전자 발현 을 억제하는 경향을 보여주었다. 또한, 토르토사이드 A는 25 ng/
ml의 EGF처리로 증가된 MUC5AC 기도뮤신의 생성도 억제하 였다. 각 처리 농도별 뮤신의 양은, 대조군, EGF 25 ng/ml단독 처리군, 토르토사이드 A 10-6 M + EGF, 토르토사이드 A 10-5 M + EGF, 토르토사이드 A 10-4 M + EGF처리군에서 각각 100±4%, 234±15%, 180±3%, 173±6%, 134±7% 이었다(Fig.
3(A), (B)).
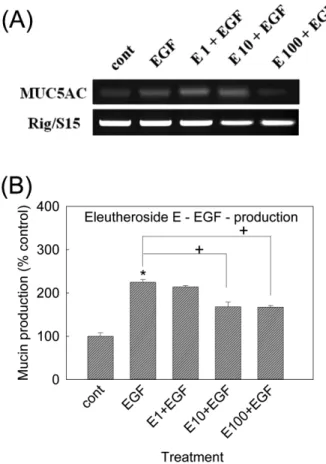
엘류쎄로사이드 E가 EGF로 자극된 MUC5AC 기도뮤신 유 전자 발현과 뮤신의 생성에 미치는 영향
엘류쎄로사이드 E역시 EGF처리로 증가된 MUC5AC 유전자 발현을 억제하는 경향을 보여주었다. 동시에, 엘류쎄로사이드 E 는 25 ng/ml의 EGF처리로 증가된 MUC5AC 기도뮤신의 생성도 억제하였다. 각 처리 농도별 뮤신의 양은, 대조군, EGF 25 ng/ml 단독 처리군, 엘류쎄로사이드 E 10-6 M + EGF, 엘류쎄로사이드 E 10-5 M + EGF, 엘류쎄로사이드 E 10-4 M + EGF처리군에서 각각 100±8%, 225±6%, 214±3%, 168±11%, 167±4% 이었다 (Fig. 4(A), (B)).
살바도라사이드가 EGF로 자극된 MUC5AC 기도 뮤신 유전 자 발현과 뮤신의 생성에 미치는 영향
살바도라사이드도 EGF처리로 증가된 MUC5AC 유전자 발현 을 억제하는 경향을 보여주었다. 또한, 살바도라사이드는 25 ng/
Fig. 2 − Effect of coniferin on EGF-induced MUC5AC gene expression and mucin production from NCI-H292 cells.
NCI-H292 cells were pretreated with varying concentrations of coniferin for 30 min and then stimulated with EGF(25 ng/ml) for 24 h. MUC5AC gene expression was measured by RT-PCR.
As a quantitative control, primers for Rig/S15 rRNA, which encodes a small ribosomal subunit protein, a housekeeping gene that was constitutively expressed, were used(A). Cell lysates were collected for measurement of MUC5AC mucin production by ELISA. Each bar represents a mean±S.E.M. of 3 culture wells in comparison with that of control set at 100%
(B). Three independent experiments were performed and the representative data were shown.
* significantly different from control(p<0.05).
+ significantly different from EGF alone(p<0.05).
(cont: control; C: coniferin; Concentration unit is µM.)
ml의 EGF처리로 증가된 MUC5AC 기도뮤신의 생성을 억제하 였다. 각 처리 농도별 뮤신의 양은, 대조군, EGF 25 ng/ml단독 처리군, 살바도라사이드 10-6 M + EGF, 살바도라사이드 10-5 M + EGF, 살바도라사이드 10-4 M + EGF처리군에서 각각 100±8%, 225±6%, 164±2%, 163±3%, 150±4% 이었다(Fig.
5(A), (B)).
고 찰 (Discussion)
뮤신이란 serine 과 threonine 의 아미노산 반복서열 구조를 가 지는 고분자량의 점액성 당단백질(mucous glycoprotein) 로서, 전
체 점액(mucus) 중량의 2% 정도를 차지하는 물질이지만 점액 고유의 물리화학적 성질인 점탄성(viscoelasticity) 을 나타나게 하는 주요한 생화학적 구성요소인데, 선상의 펩티드 서열 (apomucin) 에 탄수화물 가지가 부착되어 이루어진 실 모양의 분 자로서 아포뮤신은 현재까지 동정된 MUC 1, 2, 3A, 3B, 4, 5AC, 5B, 6, 7, 8, 9, 11, 12, 13, 16, 17, 18, 19, 20 등 약 20여 종의 특정 뮤신 유전자(MUC) 에 의해 그 염기 서열이 결정된다. 이 가운데 MUC5AC 와 MUC5B 유전자 산물만이 기도에서 분비 되는 겔 형성 뮤신(gel-forming mucin)으로 알려져 있다. 또한,
Fig. 3 − Effect of tortoside A on EGF-induced MUC5AC gene
expression and mucin production from NCI-H292 cells NCI-H292 cells were pretreated with varying concentrations of tortoside A for 30 min and then stimulated with EGF(25 ng/ml) for 24 h. MUC5AC gene expression was measured by RT-PCR. As a quantitative control, primers for Rig/S15 rRNA, which encodes a small ribosomal subunit protein, a housekeeping gene that was constitutively expressed, were used(A). Cell lysates were collected for measurement of MUC5AC mucin production by ELISA.
Each bar represents a mean±S.E.M. of 3 culture wells in comparison with that of control set at 100%(B). Three independent experiments were performed and the representative data were shown.
* significantly different from control(p<0.05).
+ significantly different from EGF alone(p<0.05).
(cont: control; T: tortoside A; Concentration unit is µM.)
Fig. 4 − Effect of eleutheroside E on EGF-induced MUC5AC gene expression and mucin production from NCI-H292 cells.
NCI-H292 cells were pretreated with varying concentrations of eleutheroside E for 30 min and then stimulated with EGF(25 ng/ml) for 24 h. MUC5AC gene expression was measured by RT-PCR. As a quantitative control, primers for Rig/S15 rRNA, which encodes a small ribosomal subunit protein, a housekeeping gene that was constitutively expressed, were used(A). Cell lysates were collected for measurement of MUC5AC mucin production by ELISA.
Each bar represents a mean±S.E.M. of 3 culture wells in comparison with that of control set at 100%(B). Three independent experiments were performed and the representative data were shown.
* significantly different from control(p<0.05).
+ significantly different from EGF alone(p<0.05).
(cont: control; E: eleutheroside E; Concentration unit is
µM.)
객담(sputum) 또는 기도 점액 (mucus)은 침, 혈청 단백질 삼출 물, 박리된 상피세포들과 뮤신 등의 혼합물, 즉 수분, 지질, 당단 백질, 단백질, 염류, 효소, 항효소 물질, 산화성 물질, 항산화성 물질, 외인성 세균, 내인성 항균물질, DNA 등 세포 잔해, 히스 타민 등 세포 및 혈장 유래 염증매개물질과 뮤신의 혼합물을 의 미한다.15)
다수의 연구에 의하여, 호흡기 세포 및 조직의 발생, 성장, 자 극, 복구 등의 과정을 조절하는 신호들이 점액성 세포로의 화생 (metastasis) 과정에 관여한다는 사실을 알게 되었다. 이들은 염 증성 신호전달 경로와 평행으로 혹은 순차적으로 작용하거나 몇 몇 경우에서는 염증과 독립적으로 작용하기도 한다. 여기에 해
당하는 신호전달 경로로는, 상피세포 성장인자 수용체(EGFR), transforming growth factor (TGF)-β, Wnt/β-catenin, HIF-1 신 호전달 경로 등이 있다. 이들 중 다수는, 염증성 자극이나 세포 손상에 반응하여 활성화되며 현재 이런 경로들 간의 중요한 상 호작용을 규명 중에 있다. 그 중에서 EGFR 신호전달 경로를 중 심으로 살펴보면 다음과 같다. 점액성 세포로 화생의 진행 과정 에서 EGFR의 역할은, 항원 노출 흰쥐에서 EGFR tyrosine kinase 의 활성 억제가 점액성 세포로의 화생을 차단할 수 있음 을 보여준 연구에 의해 제시되었다. 흥미로운 점은, 항원에 노출 되지 않은 동물에서는 EGFR ligand 인 TGF-β의 직접적 투여에 의해서 점액성 세포로의 화생이 야기되지 않았다는 점이다. 동 물실험 및 잘 분화된 기도 상피세포에서, EGFR 은 기저부 세포 막(basolateral plasma membrane)에 국한되는 양상을 보인다. 그 러나, 치밀 결합(tight junction)에 대한 기계적 손상이 일어나면 EGFR 이 첨부 세포막(apical plasma membrane)으로 전위하고 그 위치에서 리간드의 자극을 받을 수 있게 된다 Sendai virus, cigarette smoke, chemical toxicants, proteases와 같은 매우 다 양한 종류의 신호들에 반응하여 유발되는 in vitro 점액성 세포 로의 화생 유도에 있어서도 EGFR 활성화가 요구되며, 천식, 낭 포성 섬유증, COPD 환자의 기도 상피에서 EGFR 의 발현이 증 가되는 것으로 알려져 있다. EGFR 활성화는 항원, 바이러스, 호 중성구, 흡연 등에 반응하여 나타나는 인간 기도상피 세포에서 의 MUC5AC 발현 증가와, 동물모델에서의 Muc5ac 유도와 점 액성 세포로의 화생에 있어 중요한 역할을 한다는 사실이 알려 져 있다.16-18)동시에, 인체 호흡기 뮤신의 생성 및 유전자 발현 조절을 연구하는 실험모델로 자주 사용되는 인간 기도 상피세포 인 NCI-H292 세포에 대해, 상피세포 성장인자인 EGF는 호흡기 계에 존재하는 MUC5AC 뮤신의 유전자 발현을 증가시킬 수 있 는데, EGF 수용체에 결합 후 mitogen-activated protein kinase (MAPK) cascade를 활성화시키는 작용기전을 나타낸다.19-21)
임상적으로 기도점액의 과다 생성 및 분비를 유의하게 조절할 수 있는 유망한 약물은 당질 코르티코이드계(glucocorticoids) 약 물로 알려져 있으나 동반되는 광범위한 부작용이 치료 약물로서 의 효용성을 제한하고 있는 실정이다.5)동시에, 호흡기 질환의 임상에서 과다 분비된 점액의 효율적 제거를 목적으로 다수의 점 액용해제 및 거담제 등이 사용되고 있으나 그 작용 및 작용 기 전이 불명확하며 약물 투여 및 점액의 물리적 제거에 따르는 반 사적 과다분비 현상 등으로 인하여 점액 과다분비 질환의 효율 적 조절은 용이하지 않은 것으로 알려져 있다.1, 5, 15, 22) 이러한 기존의 보고에 근거하여 수행된 본 연구의 결과에서 볼 수 있는 바, 흰털오가피에서 유래한 단일성분인 코니페린, 토르토사이드 A, 엘류쎄로사이드 E, 살바도라사이드는 공통적으로 EGF 로 증 가(자극)된 MUC5AC 뮤신의 유전자 발현을 억제하였다. 동시에, 네 물질은 EGF로 증가(자극)된 MUC5AC 뮤신 당단백질의 생
Fig. 5 − Effect of salvadoraside on EGF-induced MUC5AC gene
expression and mucin production from NCI-H292 cells.
NCI-H292 cells were pretreated with varying concentrations of salvadoraside for 30 min and then stimulated with EGF(25 ng/ml) for 24 h. MUC5AC gene expression was measured by RT-PCR. As a quantitative control, primers for Rig/S15 rRNA, which encodes a small ribosomal subunit protein, a housekeeping gene that was constitutively expressed, were used(A). Cell lysates were collected for measurement of MUC5AC mucin production by ELISA. Each bar represents a mean±S.E.M. of 3 culture wells in comparison with that of control set at 100%(B). Three independent experiments were performed and the representative data were shown.
* significantly different from control(p<0.05).
+ significantly different from EGF alone(p<0.05).
(cont: control; S: salvadoraside; Concentration unit is µM.)
성(production)도 억제하였다(Fig. 2, 3, 4, 5). 이러한 약리학적 작용을 보이는 네 종류 천연물의 분자 수준에서의 작용기전은 아 직 자세히 알려진 바 없으나, EGF 수용체에 결합 후 mitogen- activated protein kinase(MAPK) cascade 관련 신호전달 경로의 특정 단계에서 억제적 작용을 나타낼 것으로 예측되는 바, 이러 한 과정에 미치는 네 약물의 작용을 탐색함으로써 분자 수준에 서의 약리작용 기전을 일부 제시할 수 있을 것으로 추정된다.
결 론 (Conclusion)
흰털오가피에서 유래한 코니페린, 토르토사이드 A, 엘류쎄로 사이드 E, 살바도라사이드는 기도 상피세포층에 존재하는 배상 세포에서의 뮤신 유전자 발현 및 생성 과정에 걸쳐 일관된 억제 작용을 나타냄으로써, 유망한 뮤신 과다생성 조절약물로 개발될 가능성을 제시하고 있다. 본 연구에서 얻어진 이러한 지견들은 호흡기 점액의 과다생성 및 분비를 보이는 천식, 만성 기관지염 등 다양한 호흡기 염증성 질환의 진행 과정에서 기도 뮤신의 과 다한 생성 및 분비 조절에 초점을 둔, 점액 조절용 신약후보물질 개발 과정에 단초를 제시함에 있어 일부 기여할 수 있을 것으로 판단된다.
감사의 말씀 (Acknowledgement)
본 연구는 한국연구재단 이공분야 기초연구사업 중 신진연구 자 지원사업에 의거하여 수행되었으며, 이에 감사 드립니다. (NRF- 2017R1C1B1005126)
References
1) Voynow, J. A. and Rubin, B. K. : Mucins, mucus, and sputum.
Chest 135(2), 505 (2009).
2) Kim, K. C., Rearick, J. I., Nettesheim, P. and Jetten, A. M. : Biochemical characterization of mucous glycoproteins synthesized and secreted by hamster tracheal epithelial cells in primary culture. J. Biol. Chem. 260, 4021 (1985).
3) Ko, K. H., Lee, C. J., Shin, C. Y., Jo, M.-J. and Kim, K. C. : Inhibition of mucin release from airway goblet cells by polycationic peptides. Am. J. Physiol. 277(21), L811 (1999).
4) Kim, K. C., Opaskar-Hincman, H. and Bhaskar, K. R. : Secretions from primary hamster tracheal surface epithelial cells in culture: Mucin-like glycoproteins, proteoglycans, and lipids. Exp. Lung Res. 15, 299 (1989).
5) Mutschler, E. and Derendorf, H. : Drug actions. CRC press, Inc., Boca Raton, Florida, p. 410 (1995).
6) Jang, I. M. : Treatise on asian herbal medicines. Haksul-pyunsu-
kwan in Research institute of natural products of Seoul National University, Seoul (2003).
7) Díaz Lanza, A.M., Abad Martínez, M.J., Fernández Matellano, L., Recuero Carretero, C., Villaescusa Castillo, L., Silván Sen, A.M. and Bermejo Benito, P. : Lignan and phenylpropanoid glycosides from Phillyrea latifolia and their in vitro anti- inflammatory activity. Planta Med. 67(3), 219 (2001).
8) Cai, X.F., Lee, I.S., Dat, N.T., Shen, G., Kang, J.S., Kim, D.H.
and Kim, Y.H. : Inhibitory lignans against NFAT transcription factor from Acanthopanax koreanum. Arch. Pharm. Res. 27(7), 738 (2004).
9) He, C., Chen, X., Zhao, C., Qie, Y., Yan, Z. and Zhu, X. : Eleutheroside E ameliorates arthritis severity in collagen- induced arthritis mice model by suppressing inflammatory cytokine release. Inflammation. 37(5), 1533 (2014).
10) Huang, L.Z., Wei, L., Zhao, H.F., Huang, B.K., Rahman, K. and Qin, L.P. : The effect of Eleutheroside E on behavioral alterations in murine sleep deprivation stress model. Eur. J.
Pharmacol. 658(2-3), 150 (2011).
11) Shao, M. X., Ueki, I. F. and Nadel, J. A. : TNF-alpha converting enzyme mediated MUC5AC mucin expression in cultured human airway epithelial cells. Proc. Natl. Acad. Sci. USA.
100(20), 11618 (2003).
12) Heo, H. J., Lee, H. J., Kim, Y. S., Kang, S. S., Son, K. H., Seok, J. H., Seo, U. K. and Lee, C. J. : Effects of baicalin and wogonin on mucin release from cultured airway epithelial cells. Phy- tother. Res. 21(12), 1130 (2007).
13) Song, K. S., Lee, W. J., Chung, K. C., Koo, J. S., Yang, E. J., Choi, J. Y. and Yoon, J. H. : IL-1beta and TNF-alpha induced MUC5AC overexpression through a mechanism involving ERK/p38 mitogen-activated protein kinase-MSK1-CREB activation in human airway epithelial cells. J. Biol. Chem.
278(26), 23243 (2003).
14) Karlinsey, J., Stamatoyannopoulos, G. and Enver, T. : Simultaneous purification of DNA and RNA from small numbers of eukaryotic cells. Anal. Biochem. 180 (2), 303 (1989).
15) Rogers, D. F. and Barnes, P. J. : Treatment of airway mucus hypersecretion. Ann. Med. 38(2), 116 (2006).
16) Deshmukh, H.S., Case, L.M., Wesselkamper, S.C., Borchers, M.T., Martin, L.D. and Shertzer, H.G. : Metalloproteinases mediate mucin 5AC expression by epidermal growth factor receptor activation. Am. J. Respir. Crit. Care Med. 171, 305 (2005).
17) Wen, F.Q., Kohyama, T., Liu, X., Zhu, Y.K., Wang, H. and Kim, H.J. : Interleukin-4-and interleukin-13-enhanced transforming growth factor-beta2 production in cultured human bronchial epithelial cells is attenuated by interferon-gamma. Am. J.
Respir. Cell Mol. Biol. 26, 484 (2002).
18) Vermeer, P.D., Einwalter, L.A., Moninger, T.O., Rokhlina, T., Kern, J.A. and Zabner, J. : Segregation of receptor and ligand regulates activation of epithelial growth factor receptor.
Nature. 422, 322 (2003).
19) Takeyama, K., Dabbagh, K., Lee, H., Agusti, C., Lausier, J. A., Ueki, I. F., Grattan, K. M. and Nadel, J. A. : Epidermal growth factor system regulates mucin production in airways. Proc.
Natl. Acad. Sci. USA. 6, 3081 (1999).
20) Takeyama, K., Dabbagh, K., Shim, J. J., Dao-Pick, T., Ueki, I. F.
and Nadel, J.A. : Oxidative stress causes mucin synthesis via transactivation of epidermal growth factor receptor: role of
neutrophils. J. Immunol. 164(3), 1546 (200).
21) Kim, J. H., Park, D.K., Lee, C. H. and Yoon, D. Y. : A new iso- flavone glycitein 7-O-beta-D-glucoside 4''-O-methylate, iso- lated from Cordyceps militaris grown on germinated soybeans extract, inhibits EGF-induced mucus hypersecretion in the human lung mucoepidermoid cells. Phytother. Res. 26(12), 1807 (2012).
22) Kim, K. D., Lee, H. J., Lim, S. P., Sikder, A., Lee, S. Y. and Lee, C. J. : Silibinin regulates gene expression, production and secretion of mucin from cultured airway epithelial cells.
Phytother. Res. 26 (9), 1301 (2012).