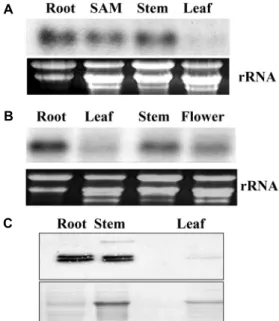
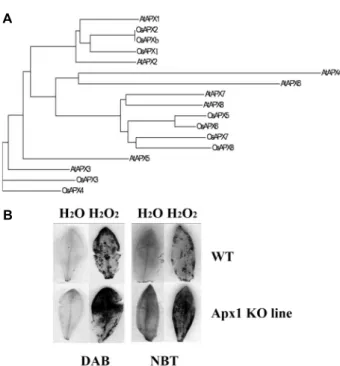
관련 문서
And based on these results, in order to utilize the witty stories appeared in the novels of Korean Pansori as actual educational texts, the brief analysis
In conclusion, it is suggested that: (a) there be educational programs for developing personality on the part of juveniles in order to prevent and
In order to defeat these problems and enhance patient’s convenience, we designed fluidal lipid matrix (FLM) for long-acting injection that can maintain the long efficacy
Conclusions : The mesiodens and the 3rd molar teeth are very similar to the inorganic component. These results provide reasonable rationale that mesiodens can be used
The proposed BMS in this thesis aimed to secure stability and increase the speed of balancing by minimizing the heat problem occurring in the cell
The analysis was done on the school section that needs to be improved first and is the most basic out of the different issued related to the high school equalization such
Secondly, in the results of the analysis as to the influence of the Taekowndo leader's leadership on the stress of a Taekowndo player, it appeared that in
From these results, we suggested that it is necessary to invigorate nature-experience in education program for alternative school so that self-esteem and