Alendronate 농도와 처리시간에 따른 MC3T3-E1와 C2C12 세포의 생존율과 Alkaline Phosphatase 활성도
경희대학교 의학전문대학원 경희의료원 산부인과학교실, 경희대학교 의학전문대학원 동서신의학병원 내과학교실1, 조선대학교 의과대학 산부인과학교실2, 고신대학교 의과대학 산부인과학교실3, 고려대학교 의과대학 산부인과학교실4
정민형․어완규1․정 혁2․이태화3․김흥열3․김 탁4
ALP Activity and Percentage of Viable Cells in C2C12 and MC3T3-E1 Cells according to Alendronate Concentration and Treatment Time
Min Hyung Jung, Wan Kyu Eo1, Hyuk Jung2, Tae Hwa Lee3, Heung Yeol Kim3, Tak Kim4 Department of Obstetrics and Gynecology, Kyung Hee Medical Center,
Department of Internal Medicine, East-West Neo Medical Center, School of Medicine, Kyung Hee University1, Seoul, Department of Obstetrics & Gynecology, College of Medicine, Chosun University2, Gwangju,
Department of Obstetrics & Gynecology, College of Medicine, Kosin University3, Busan, Department of Obstetrics & Gynecology, College of Medicine, Korea University4, Seoul, Korea
Objectives: To investigate the effect of alendronate on osteoblast proliferation and differentiation using mosue preosteoblast cell line, MC3T3-E1 and myoblastic cell line, C2C12.
Methods: C2C12 and MC3T3-E1 cells were cultured in DMEM media with various concentrations (10-9 M, 10-8 M, 10-7 M, 10-6 M, 10-5 M, 10-4 M) of alendronate for one, two, or three days. After the indicated culture times, MTT assay was performed and alkaline phosphatase activity was examined.
Results: Alendronate had no cytotoxicity regardless of the concentrations used. ALP activity in C2C12 cells was increased in alendronate treated group compared to the control group before 24 hours of treatment, but decreased rapidly after 24 hours of treatment. In MC3T3-E1, it was increased greatly at 10-8 M before 48 hours of treatment and decreased rapidly at more than 10-8 M concentration regardless of times of treatment.
Conclusion: This study shows that alendronate has a dose-dependent effect in osteoblast differentiation.
The effect may be associated with the increased expression of ALP.
Key Words: Alendronate, Osteoblast differentiation, C2C12 cell, MC3T3-E1 cell
논문접수일: 2009년 1월 10일, 심사일: 2009년 2월 13일, 게재확정일: 2009년 3월 6일
책임저자: 김흥열, 고신대학교 복음병원 산부인과학교실 Tel: 051)990-6226, Fax: 051)990-3300 E-mail: [email protected]
골 형성과 흡수는 뼈의 발달과 성장의 과정 (mode- ling)이나 오래된 뼈가 새로운 뼈로 치환 전환되는
과정 (remodeling)으로 진행된다. 일반적으로 성인의 골 형성은 modeling 과정이 끝나고 골 흡수가 re- modeling 과정에 의해서만 일어나게 된다. 결국 이러 한 골 remodeling 과정은 골을 형성하는 조골세포 (osteoblast)와 골을 흡수하는 파골세포 (osteoclast)의 세밀한 균형에 의존한다. 골다공증은 조골세포의 골
형성 기능보다 파골세포의 골 흡수 기능이 초과하여 유발된다.
비스포스포네이트는 크게 아미노기 함유 여부에 따라 비아미노 (nonamino)-와 아미노 (amino)-비스포스 포네이트의 두 종류로 구분되는데 이들의 작용기전 은 각각 다르다. 비아미노-비스포스포네이트는 파골 세포에 의해 독성 ATP로 대사되어 파골세포내 축적 되고 이들이 세포내에서 미토콘드리아의 ADP/ATP 전달체 (translocator)를 억제함으로서 파골세포의 세 포고사 (apoptosis)를 야기하는 것으로 알려져 있다.
이에 반해 아미노-비스포스포네이트는 콜레스테롤 합성경로인 메발로네이트 경로 (mevalonate pathway) 에서 farnesyl pyrophosphate (FPP) synthase 효소를 억 제함으로써 세포골격근 조절인자인 Rab, Rac, Ras, Rho, Cdc42와 같은 GTP-결합 단백질들의 prenylation 에 필요한 geranylgeranyl diphosphate (GGPP) 합성을 저해한다1. 이러한 세포 골격근 조절인자들은 파골 세포의 형성과 기능을 억제함으로 비스포스포네이 트는 결국 파골세포의 형성과 기능에 영향을 주는 것으로 알려져 있다2. 현재 골 관련 질환에서 사용되 는 약제는 이러한 아미노-비스포스포네이트이다3. Alendronate (4-amino-1-hydorxybutylidne bisphospho- nate sodium salt)는 대표적인 제3세대 아미노-비스포 스토네이트로서 골 형성에는 손상을 주지 않으면서 골 흡수 억제 효과가 100배 정도 강화된 약제이다3. 그 결과 aldendronate는 사람의 골다공증 예방과 치 료에 널리 이용되고 있다4.
이와 같이 비록 비스포스포네이트의 일차적인 작 용이 파골세포의 골 흡수 억제라 할지라도, 최근 비 스포스포네이트가 조골세포에도 영향을 준다는 연 구들이 제시되고 있다. 일부 연구는 조골세포의 증 식과 분화를 증진시키고5, 세포 고사를 감소시킨다 고 보고하고 있다6. 또한 비스포스포네이트가 섬유 모 집락형성 (fibroblastic colony formation)을 증가시 키며7, 비스포스포네이트 치료 후 피질골의 두께가 증가되었다는 보고도 있다8,9. 이러한 결과들은 비스 포스포네이트가 파골세포의 억제능력 외에도 조골 세포의 동화작용과 나아가 골 형성 촉진 효과를 가 지고 있음을 강력하게 시사하고 있다. 그러나 이러 한 비스포스포네이트가 조골세포의 형성 및 분화에
미치는 정확한 기전에 대한 분자생물학적 수준에서 의 연구는 매우 제한적이다.
그러므로 본 연구는 조골세포 분화 유도 세포주 인 C2C12와 MC3T3-E1세포를 이용하여 alendronate 농도와 처리시간에 따른 세포의 생존율과 alkaline phosphatase의 활성도에 대해 조사하였다.
대상 및 방법
1. 세포배양 및 분화
C2C12와 MC3T3-E1 세포주를 우태아혈청 (FBS, GIBCO, NE, USA)가 포함된 Dulbecco’s Modified Eagle’s Medium (DMEM, WelGENE, Daejeon, Korea) 을 사용해 37oC, 5% CO2 세포 배양기에서 배양하였 으며 계대배양을 위해 매 2~3일 간격으로 신선한 배지로 교환하였다.
동일 수 세대 세포를 다양한 농도 (10-9 M, 10-8 M, 10-7 M, 10-6 M, 10-5 M, 10-4 M)의 alendronate를 함유 한 DMEM 배지에 37oC에서 1일, 2일, 3일 동안 배양 하였다. 이때 배지는 매일 동일 농도의 alendronate를 함유한 배지로 교환하였다.
2. MTT 분석
C2C12와 MC3T3-E1 세포주를 24-well plate의 각 well당 2×104개의 세포가 들어가도록 분주한 후 세 포들이 부착할 수 있도록 1일간 5% CO2, 37oC 배양 기에서 배양한 후 부착되지 않은 세포를 제거하기 위해 배지를 교환하고, 실험군에는 alendronate를 각 각 농도별로 첨가하여 1, 2 및 3일 동안 배양하였다.
각각의 시간이 경과된 후 다양한 alendronate 농도로 배양된 세포를 5 mg/mL의 농도의 MTT (3-(4,5-dime- thylthiazol-2-yl)-2,5- diphenyl tetrazolium bromide;
Sigma, USA) 100μL씩을 각각의 well에 용액을 첨가 하고, 1시간 동안 37oC 인큐베이터에서 배양하였다.
보라색 결정이 생성되면 배지를 제거하고 100μL의 DMSO (Dimethyl sulfoxide, Sigma, USA)를 첨가하여 형성된 formazan 결정을 용해시킨 후 용해된 용액을 96-will plate에 100μL씩 분주하고 ELISA 분석기 (Spectra MAX 250, Molecular Devices Co., USA)로 540 nm에서 흡광도를 측정하였다.
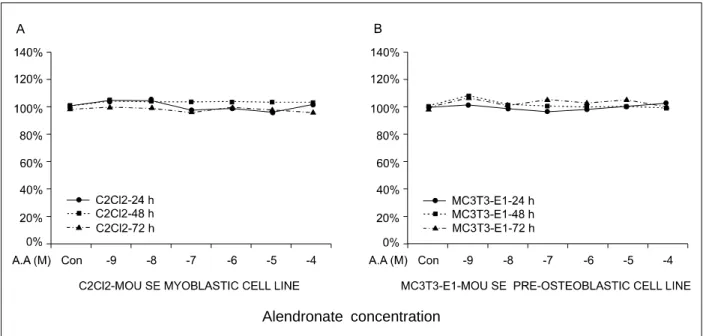
Alendronate concentration
Fig. 1. Effects of different alendronate concentrations on the percentage (%) of viable cells in C2Cl2 and MC3T3-E1 cells.
매 실험마다 세포 활성도를 대조군에 대한 백분율 로 산출하고, 각각의 실험은 3회 반복 시행하였다.
3. Alkaline phosphatase (ALP) 분석
골세포로 분화하였는지 확인하기 위하여 조골세 포에 특이적으로 발현하는 ALP 활성도를 측정하였 다. 다양한 alendronate농도로 1, 2 및 3일 동안 배양 하였다. 배양된 세포의 배지를 제거하고 PBS로 세척 한 후 0.1% Triton-X를 각각 100μL씩 넣고 시편들을 반복된 동결과 융해과정을 통하여 세포를 용해시켰 다. 20μL씩 96-well plate 상으로 옮기고, 여기에 pNPP 용액을 60μL씩 넣은 다음, 1시간 동안 배양하 고 노란색으로 발색한 것을 확인한 후, 405 nm 파장 으로 흡광도를 분석하였다.
매 실험마다 ALP 활성도는 대조군에 대한 백분율 로 산출하였고, 각각의 실험은 3회 반복 시행하였다.
4. 통계
자료는 평균±표준편차로 표시하고, 실험군 사이 의 비교는 Student's t-test로 하였으며 통계학적 유의 수준은 P값이 0.05 미만으로 하였다.
결 과
1. Alendronate의 농도와 처리시간에 따른 MC3T3-E1와 C2C12 세포의 생존율 Alendronate의 농도 (10-9 M, 10-8 M, 10-7 M, 10-6 M, 10-5 M, 10-4 M)와 시간 (24시간, 48시간, 72시간)을 변화시키면서 MTT assay로 C2C12와 MC3T3-E1의 생존도를 측정하였다. C2C12와 MC3T3-E1 세포주 모두 고농도와 시간 경과에 따른 세포 독성은 나타 내지 않았다 (Fig. 1).
2. Alendronate 농도와 처리시간에 따른 MC3T3- E1와 C2C12 세포의 ALP 활성도
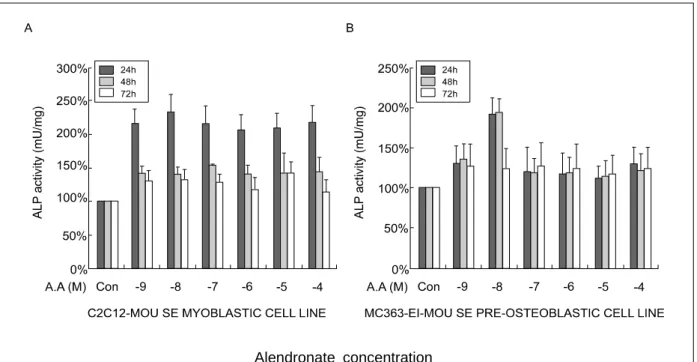
Alendronate의 농도 및 시간에 따른 골세포 분화를 확인하기 위해 조골세포에서 나타나는 특징인 ALP 활성을 측정하였다. C2C12 세포주는 alendronate 처 리 후 24시간 후 10-9 M에서 10-4 M까지 모든 농도에 서 대조군에 비해 ALP 활성이 2배 이상 증가되었다.
그러나 alendronate 처리 24시간 이후 시간이 경과함 에 따라 ALP 활성이 급격히 감소되어, 고농도 및 장 기간 동안의 alendronate 처리는 ALP 활성이 감소됨 을 알 수 있었다 (Fig. 2A).
Alendronate concentration
Fig. 2. Effects of different alendronate concentrations on ALP activity in C2Cl2 and MC3T3-E1 cells.
C2C12 세포에 비하여 MC3T3-E1에서는 10-8 M에 서 48시간까지 ALP 활성이 급격하게 증가되나 10-8 M 이상의 농도에서는 시간경과에 상관없이 ALP 활 성이 급격히 감소되었다 (Fig. 2B).
고 찰
비스포스포네이트에 의한 파골세포의 골 흡수 작 용 억제 효과는 잘 알려져 있지만, 그 기전은 명확하 게 밝혀져 있지 않다. 그러나 지금까지 이러한 작용 을 설명하는데 2가지 모델이 제시되고 있다. 첫째는 비스포스포네이트가 직접적으로 파골세포의 활성도 를 감소시킨다는 것이다10. 둘째는 조골세포를 통해 간접적으로 파골세포를 억제하는 것으로, 비스포스 포네이트는 조골세포를 자극하여 파골세포의 활성 인자 생성을 억제하거나 파골세포의 억제자를 생성 하게 한다11. 후자의 경우 비스포스포네이트는 골 형 성 작용은 손상시키지 않으면서 골 흡수 작용을 억 제하는 것으로 알려져 있다12.
파골세포는 조혈모세포로부터 기원하는 반면 조골 세포의 기원은 중간엽 줄기세포이다. 최근 아미노-
비스포스포네이트가 중간엽 줄기세포의 증식과 조골 세포로의 분화를 모두 항진시킨다고 보고하였다13. 이러한 결과들은 중간엽 줄기세포로부터 조골세포로 의 분화 촉진 기전이 아미노-비스포스포네이트가 조 골세포에 영향을 주는 방식에 있어서 중요한 기전으 로 작용할 수 있음을 시사하고 있다. 본 연구 결과는 이러한 가능성을 충분히 뒷받침하고 있다.
생체 내에서 조골세포가 bisphosphonate에 노출되 는 약리적 농도 범위는 확실하지 않다. 따라서 생리 적 상태와 흡사한 in vitro 농도를 결정하기는 어려우 나, alendronate 투여 후 흡수 공간에서 높게는 10-3 M 농도로 축적되는 것으로 보고되고 있다14-18. 이런 이 유로 본 연구에서는 alendronate를 10-9 M에서 10-4 M 농도까지 측정하였으며, alendronate가 조골세포로의 분화에 미치는 효과는 농도와 처리 시간에 따라 다 양하였다.
조골세포 분화의 초기 현상을 조사하기 위해 조골 세포 분화에 대한 3가지 모델이 주로 비교된다. 첫 번째는 생쥐유래의 전조골세포 (preosteoblast) 세포주 MC3T3-E1이고, 두 번째는 생쥐의 미분화근육세포 (myoblastic) 유래 세포주 C2C12이며, 세 번째는 두
개관의 1차 조직 유래의 조골세포이다. 현재까지 비 스포스포네이트 작용 중 특히 조골세포에 대한 작용 기전이 뚜렷이 구명된 바 없으며, 실험에 이용된 모 델마다 다소 상이한 결과가 얻어지고 있다18. 그래서 본 연구는 1차 조직 유래의 조골세포를 제외한 MC3T3-E1과 C2C12의 2가지 세포주를 이용하여 alendronate의 조골세포에 미치는 작용기전을 비교하 였다. 본 연구에서도 alendronate가 조골세포의 분화 에 미치는 효과가 세포주에 따라 약간의 차이가 발 견되었다. 그러나 이러한 미세한 차이는 alendronate 를 저농도로 단기간 처리 시 조골세포의 분화를 유 도하는 반면, 고농도로 장기간 처리 시 분화를 감소 시킨다는 기본적인 개념을 벗어나지는 않았다.
체외 alendronate가 ≥10-4mol/L 농도에서는 정상 사람의 조골세포의 생존과 증식에 영향을 미치나,
≤10-5mol/L 농도에서는 영향을 미치는 않는다고 보 고하였다19. 이러한 결과는 우리의 연구결과와 다소 상이하였다. 본 연구는 세포의 생존력은 10-9 M에서 10-4 M까지 처리한 alendronate의 농도에 영향을 받지 않았다. 이러한 결과의 차이는 실험에 이용된 세포 의 종류 차이에 기인하는 것으로 사료된다. 본 연구 에 사용된 세포는 생쥐 유래의 전조골세포 (preosteo- blast) 세포주 MC3T3-E1과 생쥐의 미분화근육세포유 래 세포주 C2C12인데 반해, 퇴행성 관절성 질환으 로 수술을 받는 환자의 수술 표본으로부터 채취한 정상적인 골세포를 이용하였다19. 이러한 골세포는 본 연구에서 사용된 세포주가 미분화된 조골세포인 데 반해, 분화가 거의 완성된 세포이기 때문에 alen- dronate에 의한 세포 독성 효과가 다르게 작용하였을 것으로 사료된다.
Alendronate가 조골세포의 분화 표지인자의 발현 에 미치는 효과 또한 표지자의 종류에 따라 처리 농 도와 시간 및 세포의 종류에 따라 다소 다른 결과를 보였다. 이는 골 형성 인자마다 서로 다른 분화시기 에 있는 조골세포의 활동성을 반영하기 때문이다20. 예를 들어, proteoglycan type I 생성과 ALP 활동성은 골기질의 형성과 관련되며, OCN은 골기질의 석회화 와 관련된다21.
비스포스포네이트가 조골세포의 증식과 분화에 작용하는 기전에 대해서는 명확하지 않다. 그러나
크게 2가지 경로를 통해 작용하는 것으로 추론되고 있다. 하나는 비스포스포네이트가 b-fobroblast growth factor (FGF)를 촉진함으로써 조골세포의 동화작용에 영향을 미친다는 것이다22.
골세포분화에 관여하는 중요한 신호전달체계는 TGF/BMP, Wnt/β-catenin, FGF, Hedgehog 및 Notch 등이 있으며23-26, 서로 상호작용을 하며 골 형성을 조절한다고 알려져 있다. 따라서 alendronate가 조골 세포의 분화 촉진 작용의 기전을 보다 명확하게 이 해하기 위해서는 향후 이러한 신호전달체계에 대한 연구가 추진되어야 할 것이다.
결론적으로 본 연구는 alendronate가 농도에 의존 적으로 조골세포의 분화를 촉진시키고 있음을 보여 주고 있으며, 이러한 alendronate에 의한 분화 촉진 효 과는 ALP증가를 야기시키고 있음을 시사하고 있다.
결 론
Alendronate는 대표적인 제3세대 아미노-비스포스 토네이트로서 일차적인 작용이 파골세포의 골 흡수 억제라 할지라도, 최근 비스포스포네이트가 조골세 포에도 영향을 준다는 연구들이 제시되고 있다.
C2C12와 MC3T3-E1세포를 FBS가 포함된 DMEM, 배양액에 배양하면서 alendronate를 다양한 농도 (10-9 M, 10-8 M, 10-7 M, 10-6 M, 10-5 M, 10-4 M)로 첨 가하고 1, 2, 3일 동안 배양한 후 MTT 분석, alkaline phosphatase (ALP) 활성도의 조골세포와 파골세포 분 화 관련 을 조사하였으며 Alendronate의 농도에 따른 세포 독성은 없었다. ALP 활성은 C2C12 세포주에서 는 alendronate 처리 후 24시간까지는 모든 농도에서 대조군에 비해 2배 이상 증가하였으나 24시간 이후 에는 시간이 경과함에 따라 급격히 감소되었다.
MC3T3-E1에서는 ALP 활성이 10-8 M에서 48시간까 지 급격하게 증가되나 10-8 M 이상의 농도에서는 시 간경과에 상관없이 급격히 감소되었다.
결론적으로 본 연구는 alendronate가 농도에 의존 적으로 조골세포의 분화를 촉진시키고 있음을 보여 주고 있으며, 이러한 alendronate에 의한 분화 촉진 효과는 ALP를 야기시키고 있음을 시사하고 있으며 앞으로 더 많은 연구가 필요할 것으로 사료된다.
국 문 초 록
목적: 본 연구는 조골세포 분화 유도 세포주인 C2C12와 MC3T3-E1세포를 이용하여 조골세포의 증 식과 분화과정 동안의 alendronate의 작용기전에 대 해 조사하고자 한다.
방법: C2C12 세포와 MC3T3-E1 세포를 DMEM 배 양액에 배양하면서 alendronate를 다양한 농도 (10-9 M, 10-8 M, 10-7 M, 10-6 M, 10-5 M, 10-4 M)로 첨가하 고 1, 2, 3일 동안 배양한 후 MTT 분석 및 alkaline phosphatase (ALP) 활성도를 측정하였다.
결과: Alendronate의 농도에 따른 세포 독성은 없 었다. ALP 활성은 C2C12 세포주에서는 alendronate 처리 후 24시간까지는 모든 농도에서 대조군에 비해 2배 이상 증가하였으나 24시간 이후에는 시간이 경 과함에 따라 급격히 감소되었다. MC3T3-E1에서는 ALP 활성이 10-8 M에서 48시간까지 급격하게 증가 되나 10-8 M 이상의 농도에서는 시간경과에 상관없 이 급격히 감소되었다.
결론: Alendronate가 농도에 의존적으로 조골세포 의 분화를 촉진시키고 있음을 보여주고 있으며, 이 러한 alendronate에 의한 분화 촉진 효과는 ALP의 발 현 증가와 연관되는 것으로 추정된다.
중심단어: 알렌드로네이트, 골아세포 분화, C2c12 세포, MC3T3-E1 세포
참 고 문 헌
1. Fisher JE, Rogers MJ, Halasy JM, Luckman SP, Hughes DE, Masarachia PJ, et al. Alendronate mechanism of action: geranylgeraniol, an interme- diate in the mevalonate pathway, prevents inhi- bition of osteoclast formation, bone resorption, and kinase activation in vitro. Proc Natl Acad Sci USA 1999;96:133-8.
2. Rodan GA, Martin TJ. Therapeutic approaches to bone diseases. Science 2000;289:1508-14.
3. Millett PJ, Allen MJ, Bostrom MP. Effects of alendronate on particle-induced osteolysis in a rat model. J Bone Joint Surg Am 2002;84-A:236-49.
4. Russell RG, Rogers MJ. Bisphosphonates: from the laboratory to the clinic and back again. Bone 1999;25:97-106.
5. Im GI, Qureshi SA, Kenney J, Rubash HE, Shanbhag AS. Osteoblast proliferation and matu- ration by bisphosphonates. Biomaterials 2004;25:
4105-15.
6. Aubin JE. Bone stem cells. J Cell Biochem Suppl 1998;30-31:73-82.
7. Giuliani N, Girasole G, Pedrazzoni M, Passeri G, Gatti C, Passeri M. Alendronate stimulates b-FGF productin and mineralized nodule formation in hman osteoblastic cells and osteoblastogenesis in human bone marrow cultures. J Bone Miner Res 1995;10:S171.
8. Canvassieux PM, Arlot ME, Reda C, Wei L, Yates AJ, Meunier PJ. Histomorphometric assessment of the long-term effects of alendronate on bone quality and remodeling in patients with osteo- porosis. J Clin Invest 1997;100:1475-80.
9. Lian JB, Stein GS, Aubin JE. Bone formation:
Maturation and functional activities of osteoblast lineage cells. In primers on the metabolic bone diseases and disorders of mineral metabolism.
Favus MJ. ed; American Society for Bone and Mineral Research: Washington DC, 2003, chapter 3.
10. Alakangas A, Selander K, Mulari M, Halleen J, Lehenkari P, Monkkonen J, et al. Alendronate disturbs vesicular trafficking in osteoclasts. Calcif Tissue Int 2002;70:40-7.
11. Schoppet M, Preissner KT, Hofbauer LC. RANK ligand and osteoprotegerin: paracrine regulators of bone metabolism and vascular function. Arterioscler Thromb Vasc Biol 2002;22:549-53.
12. Van Beek ER, Lowik CW, Papapoulos SE. Bis- phosphonates suppress bone resorption by a direct effect on early osteoclast precursors without affec- ting the osteoclastogenic capacity of osteogenic cells: the role of protein geranylgeranylation in the
action of nitrogen-containing bisphosphonates on osteoclast precursors. Bone 2002;30:64-70.
13. Von Knoch F, Jaquiery C, Kowalsky M, Schaeren S, Alabre C, Martin I, et al. Effects of bisphospho- nates on proliferation and osteoblast differentiation of human bone marrow stromal cells. Biomaterials 2005;26:6941-9.
14. Sato M, Grasser W, Endo N, Akins R, Simmons H, Thompson DD, et al. Bisphosphonate action.
Alendronate localization in rat bone and effects on osteoclast ultrastructure. J Clin Invest 1991;88:
2095-105.
15. Sama AA, Khan SN, Myers ER, Huang RC, Cammisa FP Jr, Sandhu HS, Lane JM. High-dose alendronate uncouples osteoclast and osteoblast function: a study in a rat spine pseudarthrosis model. Clin Orthop Relat Res 2004;425:135-42.
16. Reinholz GG, Getz B, Pederson L, Sanders EL, Subramaniam M, Ingle JN, Spelsberg TC. Bisphos- phonates directly regulate cell proliferation, diffe- rentiation, and gene expression in human osteo- blasts. Cancer Res 2000;60:6001-7.
17. Sahni M, Guenther HL, Fleisch H, Collin P, Martin TJ. Bisphosphonates act on rat bone resorption through the mediation of osteoblasts. J Clin Invest 1993;91:2004-11.
18. Mackie PS, Fisher JL, Zhou H, Choong PF.
Bisphosphonates regulate cell growth and gene expression in the UMR 106-01 clonal rat osteo- sarcoma cell line. Br J Cancer 2001;84:951-8.
19. Garcia-Moreno C, Serrano S, Nacher M, Farre M, Diez A, Marinoso ML, et al. Effect of alendronate on cultured normal human osteoblasts. Bone 1998;
22:233-9.
20. Kushida K, Takahashi M, Kawana K, Inoue T.
Comparison of markers for bone formation and resorption in premenopausal and postmenopausal subjects, and osteoporosis patients. J Clin Endo- crinol Metab 1995;80:2447-50.
21. Stein GS, Lian JB. Molecular mechanisms media- ting proliferation/differentiation interrelationships during progressive development of the osteoblast phenotype. Endocr Rev 1993;14:424-42.
22. Giuliani N, Pedrazzoni M, Negri G, Passeri G, Impicciatore M, Girasole G. Bisphosphonates sti- mulate formation of osteoblast precursors and mineralized nodules in murine and human bone marrow cultures in vitro and promote early steoblastogenesis in young and aged mice in vivo.
Bone 1998;22:455-61.
23. Shi Y, Massague J. Mechanisms of TGF-beta signaling from cell membrane to the nucleus. Cell 2003;113:685-700.
24. Westendorf JJ, Kahler RA, Schroeder TM. Wnt signaling in osteoblasts and bone diseases. Gene 2004;341:19-39.
25. Rapraeger AC, Krufka A, Olwin BB. Requirement of heparan sulfate for bFGF-mediated fibroblast growth and myoblast differentiation. Science 1991;
252:1705-8.
26. Nobta M, Tsukazaki T, Shibata Y, Xin C, Moriishi T, Sakano S, et al. Critical regulation of bone morphogenetic protein-induced osteoblastic differen- tiation by Delta1/Jagged1- activated Notch1 signa- ling. J Biol Chem 2005;280:15842-8.