한
한국 국 중 중부 부 서 서해 해안 안 경 경기 기만 만 일 일대 대 33개 개 간 간척 척지 지의 의 토
토양 양 염 염농 농도 도와 와 식 식생 생의 의 연 연속 속분 분포 포
Soil Salinity and Continuum Distribution of Vegetation on the
Three Reclaimed Tidal Flats of Kyonggi-Bay in the Mid-West Coast of Korea
김은규·천소을1,*·주영규2·정영상3·정형근4
Eun-Kyu Kim, Soul Chun1,*Young-K. Joo2, Yeong-Sang Jung3, and Hyeung-Gun Jung4
죽전고등학교, 1연세대학교 기초과학연구소, 2연세대학교 응용과학부 생물자원공학과,
3강원대학교 농업생명과학대학, 4연세대학교 환경공학과
Juk jeon high school, Yongin 449-548, Korea
1Institute of Basic Science, Yonsei University, Wonju 220-710, Korea
2Department of Bioresources and Technology, Yonsei University, Wonju 220-710, Korea
3Department of Biological Environment, Kangwon National University, Chuncheon 200-701, Korea
4Department of Environmental Engineering, Yonsei University, Wonju 220-710, Korea
Assessing for flora distribution is necessary for land management and environmental research in reclaimed tidal flats. This study was conducted to find out the relationship between vegetation distribution and soil salinity on the three reclaimed tidal flats of Kyonggi-bay in the mid-west coast of Korea. We investigated the soil salinity and identified the vegetation at continuum distribution zones, and describe the characteristics of continuum distribution. On the reclaimed tidal flats, spatial variation of vegetation formed partially, however continuum distribution formed as the result of connection for each spatial variation along with the soil salinity. Continuum distribution was overlapped with edaphic gradient therefore high salt tolerance species occurred at high saline zones and non-salt tolerance species occurred at low saline zones. Continuum distribution was various and also clearly distinguished on the aged reclaimed tidal flats but it was not clear on the early stage of reclamation. The continuum distribution type distinguished sequential and non-sequential type. Sequential type started from high saline zone and connected to low saline zone, gradually. On the sequential type, vegetation changed from pioneer halophyte to facultative halophyte and glycophyte along with the soil salinity gradient. Non-sequential type formed by non-sequential change of soil salinity. On the non-sequential type, vegetation distribution was non-regular form because it has not changed gradually. In the aged reclaimed tidal flats, there were vegetation wilted zone where high soil salinity, and continuum distribution formed started from this zone with bare soil.
Key words: Reclaimed tidal flat, Soil salinity, Continuum distribution, Halophyte
서 언
염류토양(salt-affected soils)인 간척지는 높은 토양 염농도로 인해 초기에는 염생식물(halophyte)이 발생 하나 시간 경과와 함께 탈염이 이루어지면서 우점종 이 바뀌게 된다(Kwon et al., 1983; Kang and Shim, 1998). 간척지 내에서 식생출현 유무와 출현종이 달라 지는 현상은 토양환경에 대한 반응의 결과로(Lee et
al., 1983; Lee et al., 2000), 장소에 따라 토양특성이 다른 현상인 토양공간변이(soil spatial variation)로 설 명된다(Burrough, 1983a, b; Armstrong, 1986; Eghabll et al., 1993; Kravchenko et al., 1999). 토양환경의 공 간변이는 식생의 공간변이로 이어지게 되며(De Jong and Burrough, 1995), 염습지의 공간변이는 수분 구배 와 지형(Sanderson et al., 2000), 토양 염농도 (Culberson, 2001), 식물의 염저항성 정도(Clarke and Hannon, 1970; Mahall and Park, 1976a; Callaway et al., 1990), 침수조건(Silvestri et al., 2005), 생물학적 요소(Cooper, 1982; Mahall and Park, 1976b), 종간의 접 수 : 2007. 12. 28 수 리 : 2008. 1. 20
*연락저자 : Phone: +82337602812, E-mail: [email protected]
경쟁(Metcalfe et al., 1986; Bertness, 1991a, b) 및 침 수조건·토양 염농도·종간의 경쟁(Penninos and Callaway, 1992)등 생물학적 조건과 물리적 조건의 복 합적 요인에 의하는 것으로 알려져있다. 간척지 식생 의 경우, 염습지와 유사하여 탈염의 정도가 장소에 따 라 다른 토양공간변이(Jung et al., 2003) 현상에 의해 식물종의 출현유무와 출현종의 변화가 쉽게 구분된 다. 간척지 내에 나타나는 식생의 공간변이는 부분적 으로 이루어지지만, 비슷한 환경내성을 갖는 종들이 비슷한 환경조건 하에서 반복될 경우 하나의 유형으 로 분류되는 점(Patten and Ellis, 1995)에 근거할 때 간척지 내에 출현하는 유사한 식생 유형은 어떤 환경 요인의 영향을 받고 있음을 시사한다.
식생의 분포와 토양환경 간의 관계에 대하여 Park(1969)은 주안 해안에서 염생식물의 연속적인 분 포가 해안가로부터의 이격거리에 따른 토양의 NaCl 함량이라고 하였고, Kim(1971)은 목포지방의 간척지 에서 식생의 출현을 결정하는 것은 조사기점으로부터 내륙을 향한 이격거리에 따른 염소이온(Cl-) 농도의 차이라고 설명하였다. Kim and Lee(1983a, b)는 염분 농도에 따라 칠면초(Suaeda japonica), 갯개미취 (Aster tripolium) 및 갈대(Phragmites communis) 군 집이 연속적인 분포를 하며, 염분농도에 따라 고농도 에서 저농도 순으로 퉁퉁마디(Salicornia europaea), 칠 면초, 갈대, 갯개미취, 매자기(Scirpus fluviatilis) 및 갯 질경(Limenium tetragonum)이 연속적인 분포를 한다 고 하여 토양환경과 식물종의 연속분포 관계를 제시 하였다.
국내 염습지 또는 간척지에서 식물종의 분포에 관 한 선행 연구는 다수 이루어졌음에도 불구하고, 분포 현상을 하나의 속성으로 표현하는 연속분포(Austin, 1985; Austin and Smith, 1989)에 대한 연구는 많지 않다. 본 연구는 간척지 내에서 발생하는 식생의 분포 현상과 토양특성 간의 관계를 규명하고자 하였으며, 중부 서해안 경기만 일대의 남양, 석문 및 시화 간척 지를 대상으로 하였다.
재료 및 방법
연구대상지 연구대상지 3곳은 모두 한반도 중부 서해안의 경기만 내에 분포하는 간척지이다. 남양 간 척지는 경기도 화성군 남양면 장덕1통 앞(동경 126˚
40′∼126˚50′, 북위 37˚00′∼37˚10′)에, 석문 간척 지는 당진군 석문면(동경 126˚30′∼126˚40′, 북위 36˚50′∼37˚00′)에, 시화 간척지는 경기도 안산시 시화호(동경 126˚40′∼126˚50′, 북위 37˚10′∼37˚
20′)에 인접하여 각각 위치한다(Fig. 1). 간척지 조성 기간은 남양 간척지의 경우, 1991년 9월부터 2002년 3
월, 석문 간척지는 1987년 8월부터 1995년 12월, 시화 간척지의 경우 1987년 6월부터 1993년 12월까지이다.
연구지역의 4월부터 11월까지 평균 기온은 16.7℃이고 총강수량은 1,144∼1,314 mm 이다.
연속분포 현상 기록 한국 중부 서해안 경기만 일 대의 남양, 석문 및 시화 간척지에서 2002년 4월부터 11월까지 현장관찰에 의하여 연속분포를 보이는 지역 에서 연속분포 현상과 출현하는 식물종을 기록하였고 토양 염농도를 측정하였다. 식물종 동정과 식물명은 대한식물도감(Lee, 1999)과 원색 한국식물도감(Lee, 2002)에 준하였고, 귀화식물원색도감 보유편(Park, 2001), 한국의 외래·귀화식물(Park, 2001)을 참조하였 다. 염생식물과 중성식물에 대한 구분은 국내에서 기 존에 연구된 자료(Kim, 1983; Ihm, 2001)와 외국의 연 구된 자료(Zhao et al., 2002; USDA, 1999)를 참고로 하였고, 간척 초기에 다른 식물이 정착하지 않은 상태 에서 개척자로 간척지에 출현하는 종은 선구종 염생식 물군으로 분류하였고, 선구종 염생식물군 출현 이후에 출현하는 염생식물은 저염생식물군으로 분류하였다.
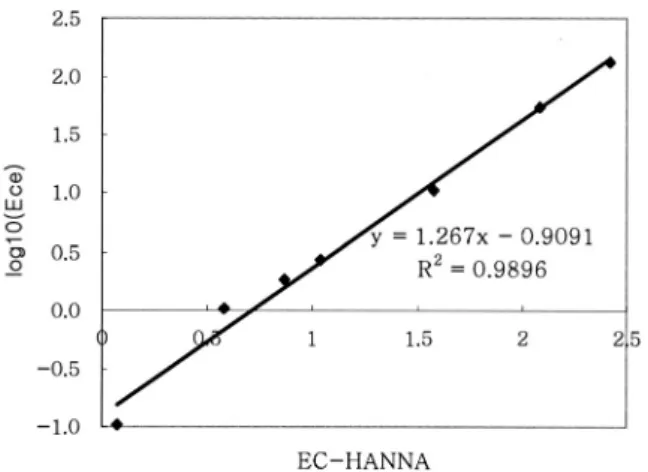
토양 염농도 측정 및 토양 시료 분석 토양 염농 도의 척도로 전기 전도도(electric conductivity, ECe) 를 활용하였고(USSL, 1954), 토양 염농도 측정은 일 정한 연속분포양식을 띠고 있는 곳을 선택하여 측정 하였다. 연속분포양식은 식생이 없는 지점을 기점으 로 식물종이 순서적으로 발생하여 연속분포의 구분이 뚜렷한 곳을 대상으로 하였다. 측정은 토양 염도계 (HANNA: HI 7031) 탐침을 식생 분포지 근권부 표 Fig. 1. Study sites(Namyang, Seukmun and Sihwa) of Kyonggi-bay in the mid-west coast of Korea.
층(깊이 0∼10 cm)에 삽입하여 현장에서 실시하였고, EC meter 값으로 보정하였다. 보정한 관계는 Fig. 2 와 같다. 측정횟수는 동일 특성을 가진 분포지에서 3 지점 이상 반복 측정하였으며, 한 개의 측정 지점에서 는 다시 짧은 거리 간격을 두고 최소 3회 이상 반복 하여 측정하였다. 간척지 토양의 특성을 분석하기 위 하여 식생분포지에서 0∼10, 10∼30 cm 깊이에서 토 양시료를 채취하여 풍건한 후 2 mm 토양체로 조제 한 후 분석에 사용하였다. 토양분석은 pH(1:5)는 초 자전극법, 양이온은 Mehlich Ⅲ 추출액으로 추출하여 atomic absorption spectrophotometer(AAS, Perkin- Elmer 2380)로 분석하였고(Tran and Simard, 1993), 토성은 pipetting 법으로 분석하였다(Sheldrick and Wang, 1993).
결 과
3개 간척지 토양의 물리·화학적 특성 3개 간척지 토양에서 식생 분포지의 물리·화학적 특성을 Table 1로 나타냈다. 3개 간척지 모두 미사질 함량이 많은 미사질양토로 나타났다. pH는 3개 간척지가 7.3∼7.9 의 범위를 보였고, 간척 년 수가 가장 짧은 남양에서 간척 후 경과 년 수가 오래 된 석문과 시화 보다 높 았다. Ca2+는 시화에서, Mg2+와 K+는 남양에서, Na+ 는 석문에서 각각 높았다.
남양 간척지 남양 간척지의 식생발생 유형을 식 생이 없는 곳을 기점으로 하여 출현한 식생 순서대로 토양 염농도를 측정하여 Fig. 3으로 나타냈다. 간척 초기 지역인 남양의 식생분포는 식생이 없는 곳과 선 구종 염생식물이 발생한 곳으로 나타났으나 선구종 염생식물인 칠면초, 퉁퉁마디, 해홍나물(Suaeda maritima) 및 나문재(Suaeda glauca)가 대부분 괴상 으로 분포하여 연속분포 유형이 뚜렷하지 않았다.
TypeⅠ은 식생 비분포지를 기점으로 칠면초, 해홍나 물 및 퉁퉁마디가 분포한 유형으로, 식생 비분포지에 비하여 식생 분포지의 토양 염농도가 높은 수준이나 식생 분포지 간의 토양 염농도는 뚜렷한 차이가 없었 다. TypeⅡ는 식생 비분포지를 기점으로 해홍나물, 나문재 및 해홍나물이 분포한 유형으로, 식생 비분포 지의 토양 염농도가 식생 분포지에 비하여 높으나 식 생 분포지간에는 큰 차이가 없었다. TypeⅢ은 식생 비분포지를 기점으로 칠면초와 해홍나물로 이어지는 유형으로, 식생 비분포지와 분포지 간에 토양 염농도 가 뚜렷한 차이를 보이지 않았다. TypeⅣ는 식생 비 분포지를 기점으로 칠면초, 해홍나물 및 퉁퉁마디가 연속분포한 유형으로, 퉁퉁마디 분포지의 토양 염농 도가 식생 비분포지보다 높았고, 해홍나물의 분포지 가 가장 낮았다. TypeⅠ∼Ⅳ의 전체 유형에서 토양 염농도는 식생 비분포지가 분포지에 비하여 높지 않 으며, 4종의 식물종(해홍나물, 나문재, 칠면초 및 퉁퉁 마디)간에도 토양 염농도의 변화가 일정하지 않아, 식 생출현 순서와 토양 염농도의 순서가 일치하지 않고 있어, 연속적인 분포로 해석하기 어렵다.
석문 간척지 석문 간척지는 경지 정리로 인해 1차 식생이 파괴되고 2차 식생이 출현한 곳이다. 남양 간 척지에 출현한 칠면초, 해홍나물, 퉁퉁마디 및 나문재 와 같은 선구종 식생이 감소하여 선구종의 단일군락 이 매우 드물었다. 현장 관찰에 의할 때 드문드문 남 아있는 교란되지 않은 곳이 연속분포와 공간적 변이 를 보인 지역이다. 이곳의 식생 발생 순서 및 토양 염 농도는 Fig. 4의 typeⅠ∼Ⅴ와 같다. 식생 분포지 중 한곳은 침수의 흔적이 있는 습지역(typeⅤ)으로 이곳 의 출현 식생은 일반 지역(typeⅠ∼Ⅳ)과 다르게 나 타났다. TypeⅠ은 식생 비분포지를 기점으로, 저염생 Fig. 2. Relationship between ECe and EC-HANNA value.
Namyang Seukmun Sihwa
1.2~46.9 17.8~68.4 21.3~48.2
37.6~78.3 28.0~74.1 36.7~64.3
9.7~31.0 3.6~11.0 5.2~11.9
7.9 7.3 7.7
9.6 11.6 8.1
1.2 3.2 6.2
1.1 0.4 0.5
22.4 4.9 1.4 Mg2+
K+ Ca2+
Na+ pH
clay silt
sand
--- cmol kg-1--- --- % ---
Table 1. Physiochemical properties of the soils taken at the vegetation occurred spots on the 3 study sites.
식물 혼생지에서 저염생식물(갈대) 분포지로 이어지 는 분포유형이다. 분포 순서에 따라 토양 염농도가 뚜 렷하게 구분되며, 토양 염농도는 저염생식물 혼생지 보다 갈대의 단일 분포지가 높았다. TypeⅡ는 식생 비분포지를 기점으로 갈대와 목본식물인 위성류 (Tamarix chinensis)를 지나 저염생식물 + 중성식물 의 혼생지(갯꾸러미풀, Puccinellia nipponica + 큰옥매 듭풀, Polygonum bellardi + 돌피, Echinochloa crusgalli) 및 중성식물 혼생지(빗자루국화, Aster subulatus + 방동사니대가리, Cyperus sanguinolentus) 로 이어지는 유형이다. 토양 염농도가 순차적으로 감 소하여 식생의 연속분포 순서와 토양 염농도의 순서 가 일치하였다. TypeⅢ은 식생 비분포지를 기점으로 하여 염생식물인 해홍나물, 갯개미취, 중성식물인 돌
피와 차풀(Cassia nomame), 저염생식물과 중성식물의 혼생지(갈대 + 빗자루국화 + 방동사니대가리) 및 저 염생식물인 갈대군집으로 이어지는 유형이다. 이 유 형에서 토양 염농도는 선구종 염생식물인 해홍나물 지점에서 높았고, 저염생식물과 중성식물의 혼생지까 지 순차적으로 감소하고, 갈대 분포지에서 다시 높아 졌다. TypeⅣ는 식생 비분포지를 기점으로 선구종 염 생식물, 저염생식물, 중성식물 + 저염생식물 혼생지 (돌피 + 사데풀, Sonchus brachyotus + 강아지풀, Setaria viridis + 빗자루국화 + 차풀 + 망초, Erigeron canadensis + 갯댑싸리, Kochia scoparia var.
littorea)를 거쳐 저염생식물인 갈대 분포지로 이어지 는 유형이다. 출현순서에 따라 선구종 염생식물에서 저염생식물 분포지까지 토양 염농도가 감소한 후 중 Fig. 3. The relationship between plant distribution order and soil salinity was not clear on Namyang: soil salinity gradient lacks consistency among pioneer halophytes and bare zone.
성식물과 저염생식물 혼생지에서 다시 증가하여 갈대 분포지에서 높아졌다. TypeⅤ는 습지역에 출현한 연 속분포 유형이다. 식생 비분포지를 중심으로 2개의 방 향으로 나뉘어지는 바, 하나는 갯꾸러미풀을 지나 토 양 염농도가 높은 새섬매자기(Scirpus planiculmis)로 이어지고, 다른 하나는 큰옥매듭풀을 지나 토양 염농
도가 낮은 큰고랭이(Scirpus tabernaemontani)로 이어 졌다. 석문 간척지에서 갈대 분포지는 저염생식물 혼 생지 및 중성식물 분포지 보다 토양 염농도가 높은 곳에 형성되어, 갈대의 내염성 범위가 넓음을 보였고 (typeⅠ,Ⅱ,Ⅲ, Ⅳ), 습지역에서는 새섬매자기가 토양 염농도가 높은 곳에 분포하여 내염성 범위가 넓음을 Fig. 4. The relationship between plant distribution order and soil salinity was clear on Seukmun: soil salinity gradient has difference and consistency among vegetation occurred zones and bare zone. Continuum distribution order of vegetation explains the soil salinity gradient order as follow: bare soil > facultative halophyte > mixed with facultative halophyte > glycophyte. These results imply that continuum distribution formed by soil salinity gradient.
보였다(typeⅤ). 일반지역과 습지역을 종합한 식생의 연속분포 유형은 토양 염농도 구배에 따라 식생 비분 포지 > 선구종 염생식물 분포지> 저염생식물∼저염생 + 중성식물 혼생지 > 중성식물 분포지의 순으로 나
타나, 토양 염농도 조건에 근거할 때 식생의 연속분포 는 토양 염농도에 의하여 결정되었다.
시화 간척지 시화 간척지는 교란되지 않은 곳으
Fig. 5. The relationship between plant distribution order and soil salinity was clear on Sihwa: soil salinity gradient has difference and consistency among vegetation occurred zones and bare zone or vegetation wilted zone. Continuum distribution order of vegetation explains that the soil salinity gradient as follow: bare soil or vegetation wilted zone > pioneer halophyte > facultative halophyte~mixed with facultative halophyte > glycophyte. These results imply that continuum distribution formed by soil salinity gradient and continuum distribution type differed from spatial zone.
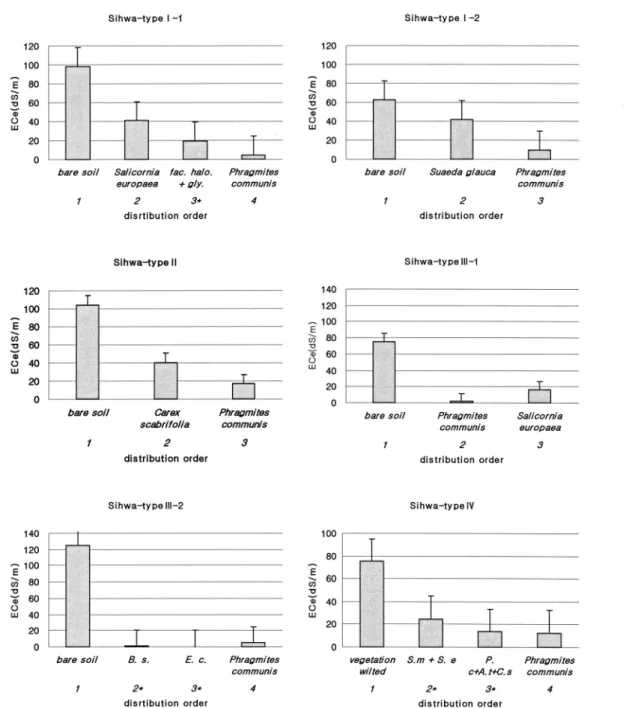
로 석문에 비하여 연속분포 유형이 더 뚜렷하게 구분 되었다. 시화 간척지의 연속분포 유형은 Fig. 5의 typeⅠ∼Ⅳ와 같다. 연속분포 순서와 토양 염농도의 순서 및 식물종의 내염성 크기가 일치하고 있어, 출현 식물종이 장소에 따라 다르며 토양 염농도의 영향을 받고 있음을 알 수 있다. TypeⅠ(Ⅰ-1, 2)은 토양 염 농도가 높은 식생 비분포지를 기점으로 토양 염농도 가 낮아지면서 선구종 염생식물, 저염생식물 순으로 이어지는 유형이다. TypeⅠ-1은 식생 비분포지를 기 점으로 퉁퉁마디, 저염생식물군+중성식물군 혼생지 (갯개미취 + 가는갯는쟁이, Atriplex gmelini + 붉은 서나물, Erechitites hieracifolia + 갯댑싸리 + 빗자루 국화 + 갈대) 및 갈대의 순으로 이어지는 유형이며, TypeⅠ-2도 식생 비분포지를 기점으로 선구종 염생 식물, 저염생식물의 순으로 이어져 연속분포 순서와 토양 염농도의 순서가 일치하고 있다. TypeⅡ는 토양 염농도가 높은 식생 비분포지를 기점으로 토양 염농 도가 낮아지면서 저염생식물 중 토양 염농도가 높은 곳에 출현한 천일사초(Carex scabrifolia)를 지나 갈대 군으로 이어지는 유형이다. TypeⅢ(Ⅲ-1, 2) 유형은 비순차적 유형으로, typeⅠ및 Ⅱ와 다르게 식생의 연 속분포에 따른 토양 염농도의 변화가 순차적이지 않 음을 보이고 있다. TypeⅢ-1은 토양 염농도가 높은 식생 비분포지에서 토양 염농도가 낮은 갈대군을 지 나 토양 염농도가 높은 선구종 염생식물 분포지로 이 어지는 유형이다. TypeⅢ-2는 토양 염농도가 높은 식 생 비분포지를 기점으로 토양 염농도가 매우 낮은 중
성식물(개피와 돌피)을 지나, 토양 염농도가 중성식물 분포지보다 약간 높은 갈대 분포지로 이어지는 유형 이다. TypeⅢ(Ⅲ-1, 2) 유형은 토양 염농도의 변화가 순차적이지 않지만 출현하는 식물종의 내염성은 토양 염농도와 일치하는 양상을 보인다. TypeⅣ는 토양 염 농도가 높은 식생 고사지점을 기점으로 토양 염농도가 순차적으로 낮아지면서 선구종 염생식물 혼생지(해홍 나물 + 퉁퉁마디), 저염생식물 혼생지(갈대 + 갯개미 취 + 천일사초) 및 갈대 분포지로 이어지는 유형이다.
시화 간척지에서 연속분포 유형은 토양 염농도 구배에 따라 식생 비분포지 및 식생 고사지 > 선구종 염생식 물 분포지 > 저염생식물 분포지> 저염생식물 혼생지 >
중성식물 분포지 순으로 나타나, 토양 염농도 조건에 근거할 때 식생의 연속분포는 토양 염농도에 의하여 결정되었다.
연속분포 유형 간척지에 출현하는 식생의 연속분 포는 간척 경과 년 수 및 토양 염농도와 밀접한 관계 를 보이며, Fig. 6, 7 및 8과 같은 3가지 유형으로 표 현된다. 이들 3가지의 연속분포 유형은 연속성의 현상 에 따라 순차적 유형(Seukmun typeⅡ, Sihwa type
Ⅰ-1, Ⅰ-2, Ⅱ 및 Ⅳ)과 비순차적 유형(Seukmun typeⅠ, Ⅲ, Ⅳ, Ⅴ, Sihwa typeⅢ-1 및 Ⅲ-2)의 2가지 로 구분된다. 순차적 유형은 간척초기를 거쳐 간척 경 과 년 수가 오래된 지역에서 나타나는 현상으로, 식생 비분포지를 기점으로 토양 염농도의 단계에 따라 내 염성이 있는 식물들이 단계적으로 분포하며 2가지로
Fig. 6. Sequential type for continuum distribution of vegetation was influenced by soil salinity and reclaimed age. Bare zone formed at high salinity zone on the year of reclamation, pioneer halophyte patch formed at high salinity zone on the early stage of reclamation. Otherwise glycophyte patch formed at low soil salinity zone on the aged site. Continuum distribution started from pioneer halophyte [ended with]
glycophyte along with soil salinity and age after reclamation
Fig. 7. Another sequential type for continuum distribution of vegetation figure out that vegetation wilted zone formed aged reclaimed tidal flats. However, bare zone and vegetation wilted zone formed at high soil salinity area. Continuum distribution started from mixed with pioneer halophyte [ended with]
glycophyte along with soil salinity and age after reclamation.
구분된다(Fig. 6과 7). 그 하나는, 간척 초기에는 토양 염농도가 가장 높은 식생 비분포지와 선구종 염생식 물 분포지가 공존하나 시간경과와 함께 점진적인 탈 염이 이루어지고 이에 적응하는 식생이 순차적으로 출현하는 유형이다(선구종 염생식물 분포지 → 저염 생식물 분포지 → 저염생식물의 혼생 분포지 → 중성 식물 분포지)(Fig. 6). 다른 하나는 간척 경과 년 수 가 오래 되어 탈염이 진행 된 것으로 예상되는 곳에 서 토양 염농도가 높은 식생 비분포지와 식생 고사지 역이 나타나고 이들 지역을 기점으로 선구종 염생식 물 혼생 분포지, 저염생식물 혼생 분포지 및 중성식물 분포지의 순으로 연속분포가 이어지는 유형이다(Fig.
7).
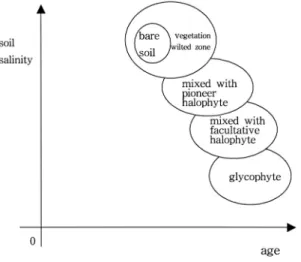
비순차적 연속분포 유형(Fig. 8)은 간척 경과 년 수 가 오래된 지역에서 나타나나, 토양 염농도와 내염성 식생의 출현이 단계적 변화를 보이지 않는다. 즉 토양 염농도가 높은 식생 비분포지를 기점으로 할 경우 다 음 단계인 선구종 염생식물로 이어지지 않고, 토양 염 농도가 낮은 저염생식물 또는 중성식물로 이어진 후 토양 염농도가 비교적 높은 선구종 염생식물 또는 저 염생식물로 이어져, 순차적인 연속성을 보이지 않는 다(Fig. 8). 그러나 출현하는 식물종의 내염성은 토양 염농도와 일치하는 양상을 보인다. 식생의 분포는 순 차적 유형 및 비순차적 유형 모두에서 토양 염농도가
높은 곳에는 내염성 식물이, 낮은 곳에는 비내염성 식 물이 출현하므로 식물종의 분포는 토양 염농도의 공 간적 변화에 기인하며, 연속분포는 토양 염농도의 연 속성에 기인함을 알 수 있다.
고 찰
토양 염농도에 따른 식생의 연속분포 현상은 간척 경과 년 수가 오래된 석문과 시화 간척지에서 뚜렷하 게 나타났으나, 간척 초기 지역인 남양에서는 일정한 유형이 형성되지 않았다. 연속분포 유형은 식생이 없 거나 고사한 지점을 기점으로 하여 선구종 염생식물, 저염생식물 및 중성식물 분포지의 순으로 나타나며, 토양 염농도는 식생이 없거나 고사한 지점 > 선구종 염생식물 분포지 > 저염생식물 분포지 > 중성식물 분 포지의 순으로 나타나 식생의 연속분포 순서와 토양 염농도의 순서가 일치하는 연속분포 유형을 보였다.
따라서 식생의 연속분포 원인은 토양속성 중의 하나 인 토양 염농도이다(Bouzille et al., 2001). 토양공간변 이는 탈염의 정도 차에 따라 발생하고, 각각의 토양공 간변이 지점에는 탈염의 정도에 따라 내염성을 달리 하는 식물종(Lee et al., 1999)이 출현 하여, 토양 염 농도의 순서와 식물종의 내염성 크기가 일치하는 연 속분포가 발생한다. 즉 토양속성은 염농도가 높은 지 점에서 낮은 지점으로 단계적으로 이어지며, 식물종 Fig. 8. Non-sequential type(continuum distribution typeⅢⅢ, ⅣⅣ) for continuum distribution was formed by spatial variation of soil salinity in the same age and the same site. Each bare zone connected to facultative halophyte/glycophyte, and its next was pioneer halophyte/facultative halophyte, respectively.
′
도 내염성이 큰 것부터 작은 것으로 단계적으로 이어 져, 토양속성과 식생이 연속분포 양상을 보인다. 이와 같은 연구의 결과는 염류토양에서 토양 염농도에 적 응할 수 있는 능력이 식물종의 분포에 생태적으로 중 요한 영향을 끼치고 있음을 나타낸다(Deeter, 2002).
간척지 내에서 중성식물이 출현하여 식물종이 변화 하는 현상은 천이현상(Kim, 1971; Joenje, 1979; Min, 1985)으로 해석되었다. 그러나 북유럽의 갯벌(bare mudflats)은 지형적 특성과 방치로 인해(Bonis et al., 2005) 간척 후 염생식물과 저염생식물(sub halophytic species)이 수십 년 이내에 사라지며(Gravesen, 1972;
Joenje, 1974; Joenje and During, 1977; Westhoff and Sykora, 1979), 방목이 이루어진 800∼1,000년이 넘은 오래된 간척지의 토양 염농도가 가장 높은 곳에서는 20년이 된 간척지에서 출현한 것과 같은 종의 다년생 염생식물이 출현하여 한번 생성된 염류토양이 전체적 으로 중성식물이 분포할 수 있는 조건으로 바뀌는 것 은 아님을 보였다(Bonis et al., 2005). 이는 간척지 전 체가 균일하게 탈염이 이루어진 조건에서 식생이 변 화해 가는 것이 아니라, 부분적으로 형성되는 탈염지 역에 선호하는 식물종이 선택적으로 발생하고 있기 때문이다. 따라서 식물종이 토양환경에 따라 선택적 으로 발생하는 선택적 공간적응은 간척 후 탈염에 의 해 염류토양의 공간변이 현상인 미소환경변화(micro environmental variation)가 지속적으로 발생하기 때문 에 나타나는 것임을 알 수 있다. 간척지에 출현하는 식물종은 간척 후 시간 경과와 함께 탈염과 같은 미 소환경변화가 심화 될수록 더 다양해질 뿐 아니라, 염 생식물이 감소하고 중성식물이 증가하는 일련의 방향 으로 전개되어 천이현상으로 해석된다(Min, 1985).
간척초기인 남양은 식생이 없는 곳과 선구종 염생 식물 분포지 간에, 그리고 선구종 염생식물종 분포지 간에 뚜렷한 연속분포 유형을 인정하기 어려웠는데, 식생의 분포 순서와 토양 염농도가 일치하지 않고 있 기 때문이다. 연속분포가 이루어지지 않은 이유는 식 생 분포지 간에 유사한 환경을 보였기 때문으로 사료 된다. 즉 간척초기이므로 교란, 탈염 또는 염분집적과 같은 공간적 변이가 없었다는 점과, 간척초기의 강한 염류토양에 이곳에서 생존이 가능한 선구종 염생식물 들만 출현하였기 때문으로 해석된다. 따라서 간척초 기 지역에서 선구종 염생식물과 식생이 없는 곳의 공 간적 변이 및 분포차이는 토양 염농도 조건에 의한 것보다는 식물 종자의 흩어짐과 관계가 있을 것으로 추정된다(Ellison, 1987; Noy-Meir and Maarel 1987).
그 이유로는 현장관찰에 의한 결과 간척초기에는 총 생으로 분포하던 염생식물이 경과 년 수가 지나면서 한 개체가 분포했던 장소를 중심으로 밀생하여, 종자
의 산포와 관계있음을 보이기 때문이다. 따라서 간척 후 1 년 차 미만인 남양에서 염생식물 간의 공간적 분포 차이는 토양환경적 요인의 영향이 적으며, 오히 려 종자의 흩어짐과 발아 등과 같은 식물 발생적 요 인이 더 큰 것으로 보이며(Shumway and Bertness, 1992; Bertness and Yeh, 1994) 토양환경 조건인 토양 염농도에 의해 분포지를 달리하는 것으로는 보이지 않는다. 이에 근거 할 때 간척 초기인 남양 전체에서 식생의 분포는 연속적인 분포를 형성하기 보다는 간 척 초기의 자연발생적인 식생 발생유형으로 괴상분포 에 더 가깝다고 보는 것이 타당한 것으로 해석된다.
석문과 시화 간척지에서는 식생의 연속분포 순서와 토양 염농도의 순서가 일치하여, 뚜렷한 연속분포 유 형을 나타내(Fig. 4와 5), 간척 경과 년 수가 길수록 식생과 분포지의 환경이 뚜렷하게 구분되는 경향이 강하였다. 식생의 분포는 토양속성과 밀접한 관계가 있는바(Adams, 1963; Vince and Snow, 1984;
Armstrong, 1985; Olff et al., 1997), 연속분포는 토양 속성의 연속성에 기인하며, 간척지가 염류토양임을 고려할 때 간척지내에서 식생의 연속분포는 탈염이 중요한 역할을 하는 것으로 판단된다. 탈염의 원인은 자연적인 세탈, 지하수위의 깊이나 토성(Min and Kim, 1997), 지형(Noordwijk et al., 1979) 등과 같은 물리적 성질에 기인하는데 동일 간척지 내에서 토성 과 지형 및 지하수위의 깊이가 같음에 근거할 때 탈 염의 원인은 이들 조건 외에, 식물에 의한 제염(Hong et al., 1969; Lee et al., 2007)과 같은 생물학적 현상 도 작용한다.
연속분포 현상은 간척 초기 지역에 비하여 간척 경 과 년 수가 오래된 지역일수록 유형이 다양하며 뚜렷 하게 나타나고 있다(Fig. 4와 5). 본 연구에서 토양 염농도 조건을 기준으로 연속분포를 분석한 결과 연 속분포에 출현하는 식물종과 토양 염농도가 일치하여 간척지에서 식물종의 연속분포는 토양 염농도의 영향 이 지배적이며, 토양 염농도에 대한 식물종의 반응으 로 해석된다.
요 약
간척지에서 식생의 분포에 대한 평가는 토양관리 및 환경적 연구를 위해 필요하다. 본 연구는 한국 중 부 서해안 경기만 내의 3개 간척지에서 식생의 분포 와 토양 염농도 간의 관계를 규명하기 위하여 실시하 였다. 식생이 연속분포를 보이는 지점에서 토양 염농 도를 측정하고 출현식물종을 분류하여 연속분포의 특 성을 기술하였다.
간척지 내에 나타나는 식생의 공간변이는 부분적으 로 이루어지지만 각각의 공간변이를 토양 염농도를
기 준 으 로 연 결 한 결 과 연 속 분 포 (continuum distribution)를 보였다. 연속분포는 토양 염농도 구배 와 일치하여 토양 염농도가 높은 곳은 내염성 식물종 이, 낮은 곳은 비내염성 식물종이 분포하였다. 연속분 포는 간척 경과 년 수가 오래된 곳에서 유형이 다양 하고 명확하게 구분되나, 간척 초기 지역에서는 연속 분포 현상이 뚜렷하지 않았다. 연속분포 유형은 순차 적 유형과 비순차적 유형으로 구분되었다. 순차적 유 형은 토양 염농도가 높은 곳을 기점으로 할 경우 토 양 염농도가 단계적으로 낮아지는 방향으로 이어지며 식 물 종 도 이 에 따 라 선 구 종 염 생 식 물 (pioneer halophyte)에서 저염생식물(facultative halophyte), 중 성식물(glycophyte)의 순으로 변화하였다. 비순차적 유형은 비연속적 토양 염농도의 변화에 의해 형성되 며, 식생의 분포도 단계적 변화를 보이지 않고 비정형 적이다. 간척 경과 년 수가 오래된 지역에서는 토양 염농도가 높은 식생 고사지역이 있고 식생 비분포지 와 함께 이 지점을 기점으로 식생의 연속분포가 형성 되었다.
인 용 문 헌
Adams, D. A. 1963. Factors influencing vascular plant zonation in North Carolina salt marshes. Ecol. 44(3): 445-456.
Armstrong, W., E. J. Wright, S. Lythe, and T. J. Gaynard. 1985.
Plant zonation and effects of the spring-neap tidal cycle on soil aeration in a Humber salt marsh. J. Ecol. 73: 323-339.
Armstrong, A. C. 1986. On the fractal dimension of some transient soil properties. Soil. Sci. Soc. Am. J. 37: 641-652.
Austin, M. P. 1985. Continuum concept, ordination methods and niche theory. Ann. Rev. Ecol. Syste. 16: 39-61.
Austin, M. P. and T. M. Smith. 1989. A new model for the continuum concept. Vegetatio 83: 35-47.
Bertness, M. D. 1991a. Interspecific interactions among high marsh perennials in a New England salt marsh. Ecol. 72: 125-137.
Bertness, M. D. 1991b. Zonation of Spartina patens and Spartina alterniflora in a New England salt marsh. Ecol. 72: 138-148.
Bertness, M. D. and S. M. Yeh. 1994. Cooperative and competitive interactions in the recruitment on marsh elders. Ecol. 75(8): 2416- 2429.
Bonis, A., J. B. Bouzille, B. Amiaud, and G. Loucougaray. 2005.
Plant community patterns in old embanked grasslands and the survival of halophytic flora. Flora 200: 74-87.
Bouzillé, J. B., E. Kerneis, A. Bonis, and B. Touzard. 2001.
Vegetation and ecological gradients in abandoned salt pans in western France. J. Vege. Sci. 12: 269-278.
Burrough, P. A. 1983a. Multiscale sources of spatial variation in soil: Ⅰ. The application of fractal concepts to nested levels of soil variation. Soil. Sci. Soc. Am. J. 34: 577-597.
Burrough, P. A. 1983b. Multiscale sources of spatial variation in soil: Ⅱ. A non-Brownian fractal model and its application in soil.
Soil. Sci. Soc. Am. J. 34: 599-620.
Callaway, R. M., S. Jones, W. R. Ferren, Jr., and A. Parikh. 1990.
Ecology of a mediterranean-climate estuarine wet-land at Carpinteria, California: plant distributions and soil salinity in the upper marsh. Can. J. Botany 69: 1139-1146.
Clarke, L. D., and N. J. Hannon. 1970. The mangrove swamp and salt marsh communities of the Sydney district. Ⅲ. Plant growth in relation to salinity and waterlogging. J. Ecol. 58: 351-369.
Cooper, A. 1982. The effects of salinity and waterlogging on the growth and cation uptake of salt marsh plants. New Phytologist 90: 263-275.
Culberson, S. D. 2001. The interaction of physical and biological determinants producing vegetation zonation in tidal mashes of the San Francisco Bay Estuary, California, USA. Ph.D. dissertation, University of California, Ecology.
De Jong, S. M. and P. A. Burrough. 1995. A fractal approach to the classification of Mediterranean vegetation types in remotely sensed images. Photogramm Engineeging Remote Senssing 61:
1041-1053.
Eghabll, B., L. N. Mielke, G. A. Calvo, and W. W. Wilhelm. 1993.
Fractal description of soil fragmentation for various tillage methods and crop sequences. Soil. Sci. Soc. Am. J. 57: 1337-1341.
Ellison, A. M. 1987. Effects of competition, disturbance, and herbivory on Salicornia europaea. Ecol. 68: 576-586.
Deeter, L. M. 2002. Sodium chloride tolerance of selected herbaceous perennials and the effects of sodium chloride on osmotic adjustment and ionic uptake in three species of herbaceous perennials. p. 28. Ph. D. dissertation, Ohio State University.
Gravesen, P. 1972. Plant communities of salt-marsh origin at Tipperne, Western Jutland. Botanisk Tidsskrift 67: 1-32.
Hong, S. W., Y. C. Hah, and Y. K. Choi. 1969. Biologcal improvement of reclaimed tidal land(Ⅰ), Desalination effects of saline soil by the growth of certain halophytes. Korean J. Botany 12(1): 7-14.
Ihm, B. S. 2001. Distribution of halophytes on the south coast of Korea. Nature Conservation 116: 9-14.
Joenje, W. 1974. Production and structure in the early stages of vegetation development in the Lauwerszee-poder. Vegetatio 29:
101-108.
Jeonje, W., and H. J. During. 1977. Colonisation of a desalination Waddenpolder by byrophyte. Vegetatio 35: 177-185.
Joenje, W. 1979. Plant succession and nature conservation of newly embanked tidal flats in the Lawerszeepolder. p. 617-634. In Jefferies, R. L. and A. J. Davy. eds. Ecological process in coastal environments. Blackwell, Oxford.
Jung, Y. S., W. H. Lee, J. H. Joo, I. H. Yu, W. S. Shin, Y. Ahn, and S. H. Yoo. 2003. Use of electromagnetic inductance for salinity measurement in reclaimed saline land. Korean J. Soil Sci. Fert.
36(2): 57-65.
Kang, B. H., and S. I. Shim. 1998. Screening of tolerant plants and development of biological monitoring technique for saline stress.
Ⅰ. Survey of vegetation in saline region and determination of saline tolerance of the plant species of the region. Korean J.
Environ. Agri. 17(1): 26-33.
Kim, C. S. 1971. An ecological study on the process of plant community formation in tidal land. Korean J. Botany 14(4): 27-33.
Kim, C. S. 1983. Distribution of halophyte community. Nature conservation 41: 31-36.
Kim, D. Y. and J. S. Lee. 1983a. Ecological studies on the halophyte community of the coast.Ⅱ. On the salt marsh of Cheongha. p.
409-416. Gunsan National University.
Kim, D. Y. and J. S. Lee. 1983b. Ecological studies on the halophyte community of the coast.Ⅱ. On the reclaimed tidal flat land of Naichodo Ri. p. 399-407. Gunsan National University.
Kravchenko, A. N., C. W. Boast, and D. G. Bullock. 1999.
Multifractal analysis of soil spatial variability. Agro. J. 91: 1033- 1041.
Kwon, H. J., W. I. Chung, and J. Y. Cho. 1983. Studies on the variation of vegetation and rice root formation accompanied with the desaltation at the reclaimed tidal fields. Korean J. Crop Sci.
28(3): 305-309.
Lee, B. M., S. I. Shim, S. G. Lee, B. H. Kang, I. M. Chung, and K.
H. Kim . 1999. Physiological response on saline tolerance between halophytes and glycophytes. Korean J. Environ. Agri. 18(1): 61- 65.
Lee, J. Y., J. O. Guh, H. S. Chang, and S. H. Bae. 1983. Weed distribution and its plant sociological aspects on the polder land.
Korean J. Weed Sci. 4(2): 135-142.
Lee, K. B., J. G. Kang, J. Li, D. B. Lee, C. W. Park, and J. D. Kim.
2007. Evaluation of salt-tolerance plant for improving saline soil of reclaimed land. Korean J. Soil Sci. Fert. 40(3): 173-180.
Lee, S. H, Y. An, S. H. Yoo, and S. M. Lee. 2000. Changes in early stage vegetation succession as affected desalinization process in Dae-Ho reclaimed land. Korean J. Environ Agri. 19(4): 364-369.
Lee, T. B. 1999. Illustrated flora of Korea. Hayng-Moon Publishing Co., Seoul.
Lee, Y. N. 2002. Flora of Korea. Kyo-Hak Publishing Co., Ltd., Seoul.
Mahall, B. E. and R. B. Park. 1976a. The ecotone between Spartina foliosa Trin. and Salicornia virginica L. in salt marshes of northern San Francisco Bay. Ⅱ. Soil water and salinity. J. Ecol. 64: 793- 809.
Mahall, B. E. and R. B. Park. 1976b. The ecotone between Spartina foliosa Trin. and Salicornia virginica L. in salt marshes of northern San Francisco Bay. Ⅲ. Soil aeration and tidal immersion. J. Ecol.
64: 811-819.
Metcalfe, W. S., A. M. Ellison, and M. D. Brtness. 1986.
Survivorship and spatial development of Spartina alterniflora Loisel. (Gramineae) seedlings in a New England salt marsh. Ann.
Botany 58: 249-258.
Min, B. M. 1985. Changes of soil and vegetation in costal reclaimed lands, west coast of Korea. Seoul National University, Ph. D.
dissertation, Department of Botany.
Min, B. M. and J. H. Kim. 1997. Soil texture and desalination after land reclamation on the west coast of Korea. Korean J. Ecol.
20(2): 133-143.
Noordwijk-Puijk, K. V., W. G. Beeftink, and P. Hogeweg. 1979.
Vegetation development on salt-marsh flats after disappearance of the tidal factor. Vegetatio 39: 1-13.
Noy-Meir, I. and E. Van der Maarel. 1987. Relations between community theory and community analysis in vegetation science:
some historical perspectives. Vegetatio 69: 5-15.
Olff, H., J. De Leeuw, J. P. Bakker, R. J. Platerink, H. J. Van Wijnen, and W. De Munck. 1997. Vegetation succession and herbivory in a salt marsh: changes induced by sea level rise and silt deposition along an elevational gradient. J. Ecol. 85: 799-814.
Park, I. K. 1969. A study of continuum of the salt plant communities in the Juan coastal area. Graduate School of Education, Seoul National University, Master thesis, Department of Biology Education.
Park, S. H. 2001. Foreign Naturalized plant of Korea. Dae-Won Publishing Co., Seoul.
Park, S. H. 2001. Colored Illustrations of Naturalized Plants of Korea. Ilchokak Co., Seoul.
Patten, R. S. and J. E. Ellis. 1995. Patterns of species and community distributions related to environmental gradients in an arid tropical ecosystem. Vegetatio 117: 69-79.
Penninos, S. C. and R. M. Callaway. 1992. Salt marsh plant zonation: The relative importance and physical factors. Ecol.
73(2): 681-690.
Sanderson, E. W., S. L. Ustin, and T. C. Foin. 2000. The influence of tidal channels on the distribution of salt marsh plant species in Petaluma Marsh, CA, USA. Plant Ecol. 146: 29-41.
Sheldrick, B. H. and C. Wang. 1993. Particle Size Distribution. p.
499-511. In Carter, M. R. ed. Soil sampling and methods of analysis. Lewis Publishers, London.
Shumway, S. W. and M. D. Bertness. 1992. Salt stress limitation of seedling recruitment in a salt marsh plant community. Oecologia 92: 490-497.
Silvestri, S., A. Defina, and M. Marani. 2005. Tidal regime, salinity and salt marsh plant zonation. Estuarine Coastal and Shelf Science 62: 119-130.
Tran, T. S. and R. R. Simard. 1993. Mehlich Ⅲ-Extractable Elements. p. 43-49. In Carter, M. R. ed. Soil sampling and methods of analysis. Lewis Publishers, London.
US Salinity Laboratory Staff. 1954. Diagnosis and improvement of Saline and alkali soils. USDA Handbook No. 60.
USDA-ARS George E. Brown, Jr. Salinity Lab. 1999. Halophyte database salt-tolerance plants and their uses. USDA ARS by N.P.
Yensen
http://www.ussl.ars.usda.gov/pls/caliche/halophyte.preface Vince, S. W. and A. A. Snow. 1984. Plant zonation in an Alaskan
slat marsh. Ⅰ. Distribution, abundance, and environmental factors.
J. Ecol. 72: 651-667.
Westhoff, V. and K. V. Sykora. 1979. A study of the influence of desalination of the Juncetum gerardii. Acta Botanica Neerlandica 28: 505-512.
Zhao, K., F. Hai, and I. A. Ungar. 2002. Survey of halophyte species in China. Plant Sci. 163(3): 491-498.