Raw 264.7 대식세포에서 Chrysophanol의 LPS로 유도된 전염증성 사이토카인 억제 효과
반연예1*․박의성2*․홍근혜1․임양이1․박건영1
1차의과학대학교 식품생명공학과
2연세대학교 식품영양학과
Chrysophanol Suppressed the Level of Pro-Inflammatory Cytokines Induced by LPS in Raw 264.7 Macrophage Cells
Yanni Pan1*, Eui-Seong Park2*, Geun Hye Hong1, Yaung-Iee Lim1, and Kun-Young Park1
1Department of Food Science and Biotechnology, Cha University
2Department of Food and Nutrition, Yonsei University
ABSTRACT The level of pro-inflammatory cytokines was markedly suppressed by chrysophanol in lipopolysaccharide (LPS)-treated Raw 264.7 macrophage cells. Chrysophanol had no toxic effect on the Raw 264.7 cells at the treatment concentrations ranging from 30 to 60 μM. Sixty μM (High) chrysophanol exhibited the highest protection against the LPS effect on the Raw 264.7 cells, as was determined by MTT assay. Thirty μM (Low) and 60 μM concentrations of chrysophanol significantly decreased the production of the pro-inflammatory cytokines TNF-α, IL-1β, IL-6, and IL-10 in the LPS-treated Raw 264.7 cells as compared to that of LPS treatment only. LPS+Low and LPS+High also significantly suppressed the mRNA expressions of the pro-inflammatory cytokines TNF-α, IL-1β, INF-γ, and COX-2 compared to that of LPS treatment only. Moreover, LPS+Low and LPS+High significantly decreased the protein expressions of the pro-inflammatory cytokines TNF-α, IL-1β, and IL-6, and also the inflammation related proteins of iNOS and COX-2 compared to that of LPS treatment only in the cells. According to our results, chrysophanol, which is derived from plants and especially curly dock, showed suppressive effects by inhibiting the inflammatory effects induced by LPS in Raw 264.7 cells by regulating pro-inflammatory cytokines and inflammation related enzymes.
These results indicated that chrysophanol may possibly be used to treat inflammatory diseases and it is perhaps a marker of anti-inflammatory functions in plants.
Key words: chrysophanol, anti-inflammation, LPS, pro-inflammatory cytokines, Raw 264.7 cells
Received 1 October 2019; Accepted 26 November 2019 Corresponding author: Kun-Young Park, Department of Food Sci- ence and Biotechnology, Cha University, Seongnam, Gyeonggi 13488, Korea
E-mail: [email protected], Phone: +82-31-881-7159
Author information: Yanni Pan (Graduate student), Eui-Seong Park (Graduate student), Geun Hye Hong (Graduate student), Yaung-Iee Lim (Researcher), Kun-Young Park (Professor)
서 론
염증반응(inflammatory response)은 비만, 심혈관계 질 환, 당뇨, 치매, 암, 천식과 같은 질병이 발생했을 때 과도하 게 발생하며, 이로 인해 열, 통증 등의 악영향을 우리 몸에 주는 것으로 알려져 있다(Hotamisligil, 2017; Watkins 등, 1995). Lipopolysaccharide(LPS)는 지금까지 알려진 다양 한 염증 유발 관련 인자 중 하나로 우리 몸에 세포를 자극하 고 toll-like receptor-4(TLR4)와 LPS binding protein (LBP)에 영향을 미친다(Gioannini와 Weiss, 2007; Poltorak
등, 1998). LPS는 TLR4를 활성화하고, TLR4는 nuclear factor κB(NF-κB)를 활성화하면 NF-κB에서 분리된 p50, p65가 핵으로 이동하여 전사인자(transcription factor)로 작용하게 된다. 핵에서 전사인자가 활성화되면 다양한 염증 인자의 방출을 촉진하는데, 이 중 전염증성 사이토카인인 interleukin(IL)-1β, IL-6, tumor necrosis factor-α(TNF -α), IL-10과 type 1 interferon(IFN) 등을 활성화하며, 염 증관련 효소인 nitric oxide synthase(iNOS)와 cyclooxy- genase-2(COX-2)를 활성화한다(Lu 등, 2008; Xie 등, 1994; Lee 등, 2004). 특히 전염증성 사이토카인은 질병의 마커로 활용된다(Watkins 등, 1995). 또한 다양한 염증반응 은 체내 nitric oxide(NO)의 생성을 증진시키고, 이로 인해 iNOS가 활성화되며, 이러한 염증반응의 활성을 통해 re- active oxygen species(ROS)를 생성한다(Guzik 등, 2003).
따라서 LPS로 유도된 전염증성 사이토카인과 염증관련 효 소를 조절할 수 있는 다양한 물질을 찾아 질병의 염증반응을 조절하여 치료하는 것이 필요하다.
Chrysophanol은 anthraquinone family 중 하나로 균류 (Qian 등, 2011)나 식물(Yang 등, 2007)의 추출물에서 발견 된다. 연구에 의하면 chrysophanol은 염증성 질병을 억제 하는 효과가 있는 것으로 알려져 있다(Han 등, 2019; Jeong 등, 2018; Han 등, 2018). Chrysophanol은 human mast cell-1(HMC-1) 세포에서 PMACI(phorbol 12-myristate 13-acetate and calcium ionophore A233187)로 유도된 caspase 1과 NF-κB를 억제하고, IL-1β, IL-6, TNF-α, TSLP(thymic stromal lymphopoietin)의 발현을 억제한 다. 그리고 chrysophanol이 HaCaT 세포에서도 TSLP와 caspase 1의 활성을 낮추는 효과가 있다. 한편 HMC-1 세 포에서는 p53 단백질 발현을 증가시켜 증식을 억제하는 효 과를 보였다. 특히 본 연구진의 선행연구에서 법제한 소리쟁 이가 일반적인 소리쟁이보다 LPS로 유도된 전염증성 사이 토카인을 더 억제하는 것을 확인했으며, 이는 법제를 통해 chrysophanol이 증가한 결과임을 확인하였다(Park 등, 2018a; 2018b).
따라서 본 연구는 소리쟁이(법제를 하면 1.7배로 증가함) 의 활성 성분인 chrysophanol의 Raw 264.7 세포에서 세포 독성, LPS로 유도된 NO, 전염증성 사이토카인인 TNF-α, IL-1β, IL-6, IL-10, IFN-γ와 염증관련 인자인 iNOS, COX-2를 enzyme-linked immunosorbent assay(ELISA), real time-quantitative polymerase chain reaction(RT- qPCR), western blot 방법을 이용하여 항염증 효과를 확인 하였다.
재료 및 방법
Raw 264.7 대식세포 배양
Raw 264.7(a murine monocyte/macrophage cell line) 세포는 한국세포주은행(KCLB, Seoul, Korea)으로부터 분 양받아 실험에 사용하였다. Raw 264.7 세포는 10% heat inactivated fetal bovine serum(FBS, GIBCO, Grand Is- land, NY, USA)과 1% penicillin-streptomycin solution (Welgene, Gyeongsan, Korea)이 포함된 DMEM(Dulbe- cco’s modified Eagle’s media, GIBCO)을 사용하여 37°C, 5% CO2 incubator에서 배양하였으며 일주일에 2~3번씩 계대 배양하였다.
MTT assay
Cell counter(Luna automated cell counter, Logos Bio- systems, Gyeonggi-do, Korea)를 이용하여 Raw 264.7 세포의 수를 측정하고, 96-well plate에 well 당 2×105 cells/mL 농도로 37°C, 5% CO2 incubator에서 24시간 동 안 배양하였다. 그 후 배지만 제거하고 30~80 μM 농도의 chrysophanol(Sigma-Aldrich Co., St. Louis, MO, USA) 및 LPS(2 μg/mL, Sigma-Aldrich Co.)를 배지로 희석해 각 well 당 100 μL씩 첨가한 후 48시간 동안 배양하였다. 그
후 배지를 제거하고 500 μg/mL 농도의 MTT solution을 배지로 희석해 각 well에 100 μL씩 분주하고 4시간 동안 배양하였다. 생성된 formazan 결정을 dimethyl sulfoxide 에 용해시키고, Wallac Victor3 1420 Multilabel Counter (Perkin-Elmer, Wellesley, MA, USA)를 사용하여 550 nm에서 흡광도를 측정하였다.
NO 생성능 측정
NO 생성능은 Griess 시약(Sigma-Aldrich Co.)을 이용 하여 측정하였다. Cell counter(Luna automated cell count- er, Logos Biosystems)를 이용하여 Raw 264.7 세포의 수 를 측정하고, 6-well plate에 2×105 cells/mL의 농도로 2 mL의 배지를 첨가한 후 24시간 동안 37°C, 5% CO2 in- cubator에서 배양하였다. 배양된 Raw 264.7 세포에 30 (Low) 및 60(High) μM 농도의 chrysophanol과 LPS(2 μg/
mL)를 48시간 동안 처리한 후 배지에 Griess 시약을 처리하 고, Wallac Victor3 1420 Multilabel Counter(Perkin El- mer)를 이용하여 550 nm에서 흡광도를 측정하였다.
Enzyme-linked immunosorbent (ELISA) assay를 이용 한 전염증성 사이토카인 측정
6-well plate에 well 당 2×105 cells/mL 농도로 37°C, 5% CO2 incubator에서 24시간 동안 배양하였다. 배지를 제거한 후 30(Low) 및 60(High) μM의 chrysophanol과 LPS(2 μg/mL)를 각 well에 처리하고, 이를 48시간 동안 배양한 후 ELISA kit(BioLegend, San Diego, CA, USA)을 사용하여 TNF-α, IL-1β, IL-6, IL-10의 농도를 측정하였 다. 구체적인 실험방법은 제조사가 제공한 방법에 따라 수행 하였다.
Real time-quantitative polymerase chain reaction (RT- qPCR)을 이용한 전염증성 사이토카인, 염증관련 효소 유전 자의 mRNA level 측정
6-well plate에 well 당 세포를 2×105 cells/mL 농도로 24시간 동안 37°C, 5% CO2 incubator에서 배양하였다. 배 지를 제거한 후 30(Low) 및 60(High) μM의 chrysophanol 과 LPS(2 μg/mL)를 배지로 희석해 각 well에 2 mL씩 첨가 하였다. 48시간 후 배지를 제거한 다음 Trizol(Invitrogen, Carlsbad, CA, USA)을 샘플당 1 mL씩 처리하여 RNA를 분 리하였다. RNA 정량은 Nano Drop ND-1000(NanoDrop Technologies Inc., Wilmington, DE, USA)을 이용하였으 며, 정량된 RNA는 Superscript Ⅱ reverse transcriptase (Invitrogen)를 이용하여 cDNA로 합성하였다. 합성된 cDNA 는 thermal cycler BioRad CFX-96 real time system(Bio -Rad, Hercules, CA, USA)을 이용하여 유전자 발현을 분 석하였다. 유전자로는 18S rRNA, TNF-α, IL-1β, IFN-γ 와 COX-2를 사용하였고, 사용한 primer 서열은 다음과 같 다: 18S rRNA forward 5’-TCGAGGCCCTGTAATTG
Chrysophanol
c d ab ab
a ab
0 20 40 60 80 100 120
30 40 50 60 70 80
Concentration (μM)
Cell viability (%) .
A
LPS+chrysophanol
d c a
b
c
e f 0 10 20 30 40 50 60 70 80
LPS 30 40 50 60 70 80
Concentration (μM)
Cell viability (%) .
B
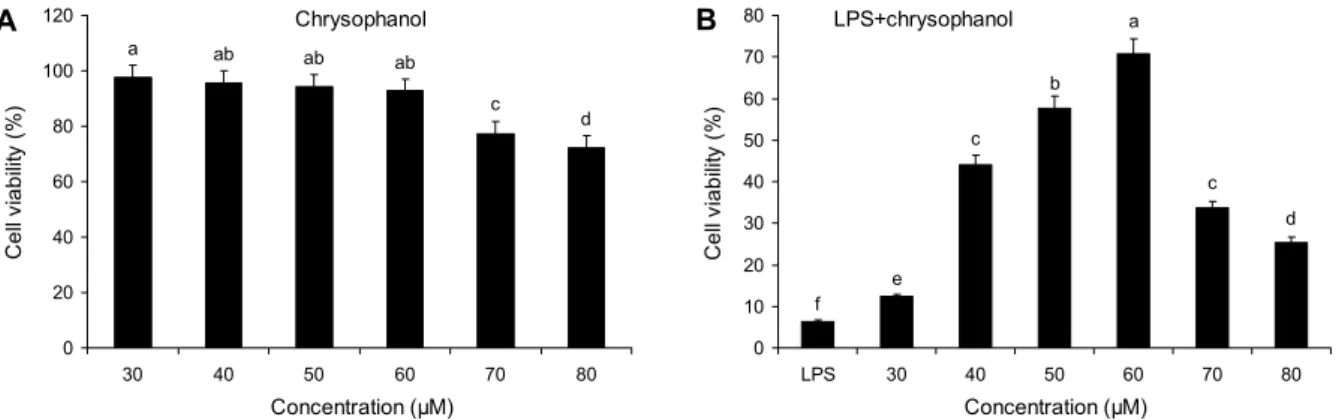
Fig. 1. Viability of Raw 264.7 cells by the treatment of chrysophonal and LPS. The cells were induced with various concentrations (30, 40, 50, 60, 70, 80 μM) for 48 hours. Means with different letters (a-f) above the bars are significantly different (P<0.05) by Duncan multiple range test.
GAA-3’, reverse 5’-CCCTCCAATGGATCCTCGTT-3’, TNF-α forward 5’-CAGGCGGTGCCTATGTCTC-3’, re- verse 5’-CGATCACCCCGAAGTTCAGTAG-3’, IL-1β forward 5’-AAGGGCTGCTTCCAAAC-3’, reverse 5’-C TCCACAGCCACAATGA-3’, IFN-γ forward 5’-GCTTT GCAGCTCTTCCTCAT-3’, reverse 5’-GTCACCATCC TTTTGCCAGT-3’, COX-2 forward 5’-GGTGCCTGGT CTGATGATG-3’, reverse 5’-TGCTGGTTTGGAATAG TTGCT-3’.
Western blot을 이용한 전염증성 사이토카인, 염증관련 효 소 유전자의 단백질 발현 측정
세포의 배양 및 샘플 처리는 mRNA 측정과 동일한 방식을 사용하였다. 샘플 처리 후 세포의 배지를 제거한 다음 ra- dio-immunoprecipitation assay(RIPA, Invitrogen) buf- fer를 이용하여 단백질을 분리하였다. Bradford assay 방법 을 이용하여 단백질의 농도를 정량하였다(Bradford, 1976).
추출된 단백질을 SDS-polyacrylamide gel 전기영동으로 분리하고 polyvinylidene fluoride(PVDF, Bio-Rad) mem- brane으로 이동시킨 후, PBS-T를 함유한 5% skim milk로 비특이적인 단백질을 blocking 하였다. Blocking 후 PVDF membrane을 PBS-T 3회, PBS 1회로 세척한 다음 1차 항 체를 4°C에서 overnight 하면서 반응시켰다. 그 이후 PBS- T 3회, PBS 1회로 세척한 다음 2차 항체를 실온에서 2시간 동안 처리하였다. IL-6, IL-1β, TNF-α, iNOS, COX-2 및 β-action(Santa Cruz, Dallas, TX, USA) 1차 항체를 사용 하였고, 각각에 맞는 2차 항체를 사용한 후 LAS-4000(Fuji- film Life Science, Tokyo, Japan)을 이용하여 확인하였다.
통계처리
RT-qPCR 데이터는 평균±표준오차(standard error, SE)로 나타내었고, 이를 제외한 모든 실험의 데이터는 평균
±표준편차(standard deviation, SD)로 나타냈다. One- way analysis of variance(ANOVA)를 실시하고 Duncan’s
multiple range test를 이용하여 각 처리 시료(n=3) 간의 유의성을 검증하였다. P<0.05일 때 유의성이 있다고 간주하 였고, SPSS v18 statistical software package(SPSS Inc., Westlands, Hong Kong)를 이용하여 모든 실험을 분석하였 다.
결과 및 고찰
Chrysophanol 및 LPS+chrysophanol의 cell viability 측정
30~60 μM 범위에서 chrysophanol의 농도가 30 μM일 때 86.6±1.6%, 60 μM일 때는 91.3±2.7%로 Raw 264.7 세포의 성장에 큰 영향을 미치지 않지만, 70 μM 이상의 농 도에서는 세포독성이 확인되었다(Fig. 1A). 또한 LPS만 처 리하면 6.4±1.1%의 생존율을 보이지만 LPS와 60 μM의 chrysophanol을 함께 처리했을 때 70.7±10.1%로 LPS로 인한 세포독성을 억제하는 것으로 확인되었다(Fig. 1B).
Han 등(2019)의 연구에 의하면 chrysophanol은 HMC-1 세포에 60 ng/mL 농도로 처리했을 때 세포독성을 관찰하지 못하였고, Raw 264.7 세포에서 1, 10, 100 μM 농도로 chrysophanol을 처리했을 때 세포독성을 관찰하지 못하였 다(Rim 등, 2013). 따라서 본 실험의 결과를 바탕으로 30 μM(Low), 60 μM(High)의 chrysophanol 농도를 선정하여 이후 실험을 진행하였다.
Chrysophanol의 LPS로 유도된 NO 생성능 억제 효과 LPS가 처리된 Raw 246.7 세포에서 NO 생성능은 대조군 과 비교하여 유의적으로 증가하였다(Table 1)(P<0.05). LPS +Low(40.60±4.31 μM)와 LPS+High(14.68±0.17 μM)는 LPS(59.68±0.68 μM)군보다 NO 수준이 유의적으로 낮아 졌다(P<0.05). LPS를 처리하지 않은 Low(4.84±0.09 μM) 와 High(5.24±0.09 μM)에서는 대조군(7.89±0.25 μM)과 비교하여 NO 생성능이 유의적으로 낮아졌지만, Low와 High 군 간의 유의적인 차이는 없었다. Chrysophanol은 BV2 mu-
Table 1. Nitric oxide production in Raw 264.7 cells by treatment of LPS and chrysophanol
NO production (μM) Control
Low High LPS LPS+Low LPS+High
7.89±0.25d 4.84±0.09e 5.24±0.09e 59.68±0.68a 40.60±4.31b 14.68±0.17c
Low: chrysophanol (30 μM), High: chrysophanol (60 μM), LPS:
lipopolysaccharide (2 μg/mL), LPS+Low: lipopolysaccharide+
chrysophanol (30 μM), LPS+High: lipopolysaccharide+chrys- ophanol (60 μM).
Means with different letters (a-e) within a column are signifi- cantly different (P<0.05) by Duncan’s multiple range test.
c b a
d c ab
240 245 250 255 260 265 270 275 280 285 290 295
Contorl Low High LPS LPS+Low LPS+High
TNF-α (pg/mL) .
b b a
d c d
0 5 10 15 20 25 30 35 40 45 50
Contorl Low High LPS LPS+Low LPS+High
IL-1β (pg/mL) .
b a ab
e d c
0 100 200 300 400 500 600
Contorl Low High LPS LPS+Low LPS+High
IL-6 (pg/mL) .
c b a
e d cd
0 2000 4000 6000 8000 10000 12000
Contorl Low High LPS LPS+Low LPS+High
IL-10 (pg/mL) .
Fig. 2. Pro-inflammatory cytokines TNF-α, IL-1β, IL-6, and IL-10 concentrations in Raw 264.7 cell line by the treatment of LPS and chrysophanol in ELISA. Low: chrysophanol (30 μM), High: chrysophanol (60 μM), LPS: lipopolysaccharide (2 μg/mL), LPS+Low:
lipopolysaccharide+chrysophanol (30 μM), LPS+High: lipopolysaccharide+chrysophanol (60 μM). Means with different letters (a-e) above the bars are significantly different (P<0.05) by Duncan’s multiple range test.
rine microglia 세포에서 자유라디칼로 유도된 산화적 손상 에 의한 NO 생성을 억제하였고(Lin 등, 2015), Raw 264.7 세포에서 LPS에 의한 NO 생성도 억제하였다(Rim 등, 2013).
NO의 생성은 nitric oxide synthase(NOS)에 의해 촉매 되어 L-arginine으로부터 합성된다(Korhonen 등, 2005). NOS 는 다양한 형태의 자극에 의해 활성화되고 그로 인해 NO의 생성을 증가시키며, 다양한 유도물질 중 LPS는 전염증성 사이토카인과 iNOS를 활성화해 NO의 생성을 증가시킨다 (Korhonen 등, 2005). 따라서 실제로 chrysophanol이 LPS
로 유도한 전염증성 사이토카인을 조절하는지에 관해 연구 를 진행하였다.
Chrysophanol의 LPS로 유도된 전염증성 사이토카인, 염 증관련 효소 억제 효과
ELISA: Raw 264.7 세포에 LPS를 처리했을 때 처리하지 않은 군과 비교하여 전염증성 사이토카인인 TNF-α, IL-1 β, IL-6 및 IL-10 생성을 증가시켰다(Fig. 2). LPS와 함께 chrysophanol을 Low, High 농도로 Raw 264.7 세포에 처 리하면 LPS만 처리한 군과 비교해 TNF-α(LPS: 286.8±3.8 pg/mL, LPS+Low: 280.5±6.1 pg/mL, LPS+High: 273.0
±3.8 pg/mL), IL-1β(LPS: 43.5±3.8 pg/mL, LPS+Low:
37.3±2.3 pg/mL, LPS+High: 34.3±4.7 pg/mL), IL-6 (LPS: 551.1±7.1 pg/mL, LPS+Low: 530.6±8.4 pg/mL, LPS+High: 512.6±4.2 pg/mL) 및 IL-10(LPS: 9,571.9±
1,316.8 pg/mL, LPS+Low: 7,947.5±453.9 pg/mL, LPS+
High: 5,971.1±631.5 pg/mL)의 발현 수준으로 유의적으로 감소하였다(P<0.05). 따라서 chrysophanol은 Raw 264.7 세포에서 LPS로 유도된 전염증성 사이토카인을 감소시켰 다고 하겠다.
RT-qPCR: Raw 264.7 세포에서 TNF-α, IL-1β, IFN- γ, COX-2의 mRNA 발현은 LPS군이 미처리군에 비해 유의
b b a
c c c
0 1 2 3 4 5 6 7 8 9
Contorl Low High LPS LPS+Low LPS+High
TNF-α mRNA expression .
d c a
d d b
0 0.5 1 1.5 2 2.5
Contorl Low High LPS LPS+Low LPS+High
IL-1β mRNA expression .
cd b a
cd c c
0 0.5 1 1.5 2 2.5 3 3.5
Contorl Low High LPS LPS+Low LPS+High
IFN-γ mRNA expression .
c b a
bc bc b
0 0.2 0.4 0.6 0.8 1 1.2 1.4 1.6 1.8
Contorl Low High LPS LPS+Low LPS+High
COX-2 mRNA expression .
Fig. 3. mRNA expression of pro-inflammatory cytokines TNF-α, IL-1β, IFN-γ, and an inflammation related gene COX-2 in Raw 264.7 cells by the treatment of LPS and chrysophanol. Low: chrysophanol (30 μM), High: chrysophanol (60 μM), LPS: lipopoly- saccharide (2 μg/mL), LPS+Low: lipopolysaccharide+chrysophanol (30 μM), LPS+High: lipopolysaccharide+chrysophanol (60 μM). Means with different letters (a-d) above the bars are significantly different (P<0.05) by Duncan’s multiple range test.
적으로 높았다(P<0.05)(Fig. 3). LPS와 chrysophanol을 함 께 처리했을 때, 전염증성 사이토카인인 TNF-α의 mRNA 발현 수준은 LPS+Low군이 2.3배, LPS+High군이 3.2배 LPS군보다 감소하였다. 또한 IL-1β의 mRNA 발현 수준은 LPS+Low군이 3.2배, LPS+High군이 8.4배 LPS군보다 감소하였고, IFN-γ는 LPS+Low군이 1.7배, LPS+High군 이 3.5배 감소하였다. 염증관련 효소인 COX-2의 mRNA 발현 수준은 LPS+Low군이 1.5배, LPS+High군이 4.1배 LPS군보다 감소하여 전반적으로 chrysophanol을 처리했 을 때 전염증성 사이토카인, 염증관련 효소의 mRNA 발현 수준이 유의적으로 감소했으며(P<0.05), 농도가 높아(High) 졌을 때 낮은 농도(Low)보다 더욱 현저히 감소시켰다.
Western blot: 전염증성 사이토카인인 TNF-α, IL-1β 와 IL-6, 염증관련 효소인 iNOS와 COX-2의 단백질 발현은 LPS를 처리했을 때 미처리 군에 비해 유의적으로 증가하였 다(P<0.05)(Fig. 4). TNF-α의 단백질 발현 수준은 LPS+
Low군이 20%, LPS+High군이 60% LPS군보다 감소하였 다. 또한 IL-1β의 단백질 발현 수준은 LPS+Low군이 30%, LPS+High군이 50% LPS군보다 감소하였고, IL-6는 LPS +High군이 20% 감소하였다. iNOS의 단백질 발현 수준은 LPS+Low군, LPS+High군 모두 LPS군보다 감소하였고, COX-2도 LPS+Low군, LPS+High군이 모두 유의적으로
감소하였다(P<0.05). 이러한 결과는 ELISA에서 확인된 전 염증성 사이토카인의 생성과 mRNA 발현과 마찬가지로 단 백질 발현에서도 전염증성 사이토카인인 TNF-α, IL-1β와 IL-6, 염증관련 효소인 iNOS와 COX-2가 유의적으로 감소 하여 LPS로 증가한 전염증성 사이토카인과 염증관련 효소 를 chrysophanol이 효과적으로 억제하는 것으로 나타났다.
전염증성 사이토카인은 각각의 다른 사이토카인이 Naive T cell을 자극하여 T helper cell(Th)로 분류시키는 역할을 하며(Vinuesa 등, 2005), Th는 종류와 역할에 따라 Th1, Th2, Th17, Treg로 분류된다(Zou와 Restifo, 2010). 이러 한 전염증성 사이토카인은 전사인자인 NF-κB에 의해 활성 화되고(Xie 등, 1994), 인체의 염증반응에 관여한다(Lu 등, 2008). Raw 264.7 세포에 LPS를 처리하면 전염증성 사이토 카인과 염증관련 효소가 급격히 증가하는데(Song 등, 2017), 이는 LPS 자극으로 염증반응이 활발히 일어나 전반적인 염 증관련 인자가 증가하는 것이다.
Jeong 등(2018)의 연구에 의하면 PMACI를 처리한 HMC -1 세포에서 chrysophanol을 처리하면 NF-κB를 억제하 고, TNF-α, IL-1β, IL-6를 억제한다고 하였다. 또한 DSS 로 유도된 colitis 마우스와 LPS를 처리한 마우스 복막 대식 세포에서 chrysophanol을 처리하면 NF-κB(p65)와 IL-6, COX-2의 발현이 감소하였다(Kim 등, 2010). 따라서 chry-
TNF-α IL-1β IL-6 β-Actin
iNOS COX-2 β-Actin Control Low High LPS LPS+ LPS+
Low High
TNF-α
c b a
e d c
0 0.2 0.4 0.6 0.8 1 1.2 1.4
Folds of control .
Control Low High LPS LPS+ LPS+
Low High
IL-1β
d cd d
a b
c
0 0.2 0.4 0.6 0.8 1 1.2 1.4
Folds of control .
Control Low High LPS LPS+ LPS+
Low High
IL-6
b ab a
d d c
0 0.2 0.4 0.6 0.8 1 1.2 1.4
Folds of control .
Control Low High LPS LPS+ LPS+
Low High iNOS
c b a
e d cd
0 0.2 0.4 0.6 0.8 1 1.2 1.4
Folds of control .
Control Low High LPS LPS+ LPS+
Low High
COX-2
c b a
e d d
0 0.2 0.4 0.6 0.8 1 1.2
Folds of control .
Control Low High LPS LPS+ LPS+
Low High
Fig. 4. Protein expression of pro-inflammatory cytokines TNF-α, IL-1β, and IL-6, and inflammation related genes iNOS, and COX-2 in Raw 264.7 cell line by the treatment of LPS and chrysophanol. Low: chrysophanol (30 μM), High: chrysophanol (60 μM), LPS:
lipopolysaccharide (2 μg/mL), LPS+Low: lipopolysaccharide+chrysophanol (30 μM), LPS+High: lipopolysaccharide+chrysophanol (60 μM). Means with different letters (a-e) above the bars are significantly different (P<0.05) by Duncan’s multiple range test.
sophanol은 화합물로 유도된 전염증성 사이토카인과 염증 관련 효소의 발현을 억제하며, 이는 전사인자인 NF-κB의 발현을 억제하여 조절되는 것으로 생각된다.
Wen 등(2018)은 chrysophanol이 PPAR-γ를 활성화하 며 NF-κB의 전사를 조절하여 염증(TNF-α, IL-1β, iNOS 유전자 측정)을 억제하는 기작을 소개하였다. 그러나 PPAR- γ는 비만과 당뇨병의 주요 유발 전사인자이므로 이에 대한 더 많은 연구가 필요하다고 하겠다.
전염증성 사이토카인은 다양한 질병에서 마커로 사용되 며, 특히 염증반응에서 중요한 역할을 하고 있다(Watkins 등, 1995). 따라서 본 연구에서 chrysophanol이 LPS로 유 도된 전염증성 사이토카인을 억제한 것은 실제로 질병 발생 시 처리했을 때 염증반응을 억제할 수 있을 것으로 기대된 다. 본 연구실에서는 chrysophanol이 소리쟁이(Rumex
crispus L.)의 주요 활성 성분으로 확인되었으며, 이 chrys- ophanol의 사이토카인 생성 억제 효과를 통하여 소리쟁이 가 염증을 억제하는 중요한 기능성 원료로 사용될 수 있음을 확인하였다. 또한 이 연구에서 LPS를 처리하지 않은 Raw 264.7 대식세포에 chrysophanol의 Low와 High 농도를 처 리하면 염증관련 인자, 염증관련 mRNA 및 단백질의 발현을 낮추는 효과도 있어 chrysophanol은 일반 세포에서도 염증 을 억제할 수 있는 가능성을 보여주었다고 하겠다. 이러한 연구를 비롯하여 chrysophanol과 같은 식물유래 화합물에 대한 연구가 지속해서 필요하다고 하겠다.
요 약
Chrysophanol은 30~60 μM 범위에서 Raw 264.7 세포에
서 세포증식을 억제하지 않았다. 또한 LPS와 함께 chrys- ophanol을 처리했을 때 60 μM 처리 시 가장 높은 세포 성장 률을 보였다. Raw 264.7 세포에 LPS만 단독으로 처리했을 때 처리하지 않은 군보다 NO 생성이 증가하였으나 chrys- ophanol을 Low(30 μM), High(60 μM) 농도로 처리했을 때 NO의 생성이 유의적으로 감소하였다(P<0.05). 전염증성 사이토카인인 TNF-α, IL-1β, IL-6, IL-10은 LPS군에 비 해 chrysophanol을 Low, High 농도로 처리했을 때 유의적 으로 감소했으며, 마찬가지로 TNF-α, IL-1β, IFN-γ의 mRNA 수준도 감소하였다. 또한 염증관련 효소인 COX-2 의 mRNA 수준도 억제하였다. 단백질 발현 수준에서도 전염 증성 사이토카인인 TNF-α, IL-1β와 IL-6, 염증관련 효소 인 iNOS와 COX-2의 발현이 LPS와 chrysophanol을 Low, High 농도로 함께 처리했을 때 LPS만 처리한 군에 비해 유 의적으로 감소하였다(P<0.05). 이러한 결과를 종합해볼 때, 식물유래 화합물인 chrysophanol은 LPS로 유도된 염증반 응을 억제하며, 특히 전염증성 사이토카인과 염증관련 효소 (iNOS, COX-2)의 발현을 억제하여 염증반응을 조절하는 것으로 나타났다.
감사의 글
본 결과물은 농림축산식품부의 재원으로 농림수산식품기술 기획평가원의 고부가가치식품기술개발사업의 지원을 받아 연구되었습니다(116169-3).
REFERENCES
Bradford MM. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem. 1976. 72:248-254.
Gioannini TL, Weiss JP. Regulation of interactions of Gram- negative bacterial endotoxins with mammalian cells. Immunol Res. 2007. 39:249-260.
Guzik TJ, Korbut R, Adamek-Guzik T. Nitric oxide and super- oxide in inflammation and immune regulation. J Physiol Phar- macol. 2003. 54:469-487.
Han NR, Kim HY, Kang S, Kim MH, Yoon KW, Moon PD, et al. Chrysophanol, an anthraquinone from AST2017-01, pos- sesses the anti-proliferative effect through increasing p53 pro- tein levels in human mast cells. Inflammation Res. 2019. 68:
569-579.
Han NR, Moon PD, Yoo MS, Ryu KJ, Kim HM, Jeong HJ.
Regulatory effects of chrysophanol, a bioactive compound of AST2017-01 in a mouse model of 2,4-dinitrofluorobenzene- induced atopic dermatitis. Int Immunopharmacol. 2018. 62:
220-226.
Hotamisligil GS. Inflammation, metaflammation and immuno- metabolic disorders. Nature. 2017. 542:177-185.
Jeong HJ, Kim HY, Kim HM. Molecular mechanisms of anti-in- flammatory effect of chrysophanol, an active component of AST2017-01 on atopic dermatitis in vitro models. Int Immu-
nopharmacol. 2018. 54:238-244.
Kim SJ, Kim MC, Lee BJ, Park DH, Hong SH, Um JY. Anti-in- flammatory activity of chrysophanol through the suppression of NF-κB/caspase-1 activation in vitro and in vivo. Molecules.
2010. 15:6436-6451.
Korhonen R, Lahti A, Kankaanranta H, Moilanen E. Nitric oxide production and signaling in inflammation. Curr Drug Targets Inflamm Allergy. 2005. 4:471-479.
Lee KM, Kang BS, Lee HL, Son SJ, Hwang SH, Kim DS, et al. Spinal NF-κB activation induces COX-2 upregulation and contributes to inflammatory pain hypersensitivity. Eur J Neu- rosci. 2004. 19:3375-3381.
Lin F, Zhang C, Chen X, Song E, Sun S, Chen M, et al. Chryso- phanol affords neuroprotection against microglial activation and free radical-mediated oxidative damage in BV2 murine microglia. Int J Clin Exp Med. 2015. 8:3447-3455.
Lu YC, Yeh WC, Ohashi PS. LPS/TLR4 signal transduction pathway. Cytokine. 2008. 42:145-151.
Park ES, Song GH, Kim SH, Lee SM, Kim YG, Lim YL, et al. Rumex crispus and Cordyceps militaris mixture amelio- rates production of pro-inflammatory cytokines induced by lipopolysaccharide in C57BL/6 mice splenocytes. Prev Nutr Food Sci. 2018a. 23:374-381.
Park ES, Song GH, Lee SM, Kim TY, Park KY. Increased anti- inflammatory effects of processed curly dock (Rumex crispus L.) in ex vivo LPS-induced mice splenocytes. J Korean Soc Food Sci Nutr. 2018b. 47:599-604.
Poltorak A, He X, Smirnova I, Liu MY, Van Huffel C, Du X, et al. Defective LPS signaling in C3H/HeJ and C57BL/10ScCr mice: mutations in Tlr4 gene. Science. 1998. 282:2085-2088.
Qian ZJ, Zhang C, Li YX, Je JY, Kim SK, Jung WK. Protective effects of emodin and chrysophanol isolated from marine fun- gus Aspergillus sp. on ethanol-induced toxicity in HepG2/
CYP2E1 cells. Evid Based Complement Alternat Med. 2011.
Article ID: 452621.
Rim HK, Moon PD, Choi IH, Lee EH, Kim HM, Jeong HJ.
SoSoSo or its active ingredient chrysophanol regulates pro- duction of inflammatory cytokines & adipokine in both mac- rophages & adipocytes. Indian J Med Res. 2013. 137:142-150.
Song GH, Park ES, Lee SM, Kim TY, Park KY. An Atopic Preventive Drink (APD) reduces Th2 cytokines in LPS-treat- ed RAW 264.7 cells. TANG. 2017. 7:e15.
Vinuesa CG, Tangye SG, Moser B, Mackay CR. Follicular B helper T cells in antibody responses and autoimmunity. Nat Rev Immunol. 2005. 5:853-865.
Watkins LR, Maier SF, Goehler LE. Immune activation: the role of pro-inflammatory cytokines in inflammation, illness re- sponses and pathological pain states. Pain. 1995. 63:289-302.
Wen Q, Mei L, Ye S, Liu X, Xu Q, Miao J, et al. Chrysophanol demonstrates anti-inflammatory properties in LPS-primed RAW 264.7 macrophages through activating PPAR-γ. Int Immuno- pharmacol. 2018. 56:90-97.
Xie QW, Kashiwabara Y, Nathan C. Role of transcription factor NF-κB/Rel in induction of nitric oxide synthase. J Biol Chem.
1994. 269:4705-4708.
Yang X, Yang L, Wang S, Yu D, Ni H. Synergistic interaction of physcion and chrysophanol on plant powdery mildew. Pest Manag Sci. 2007. 63:511-515.
Zou W, Restifo NP. TH17 cells in tumour immunity and im- munotherapy. Nat Rev Immunol. 2010. 10:248-256.