J. Fish Pathol., 22(3):343 ~ 352 (2009)
343
Molecular identification and expression analysis of a natural killer enhancing factor-A from black rockfish Sebastes schlegelii
Jeong-Ho Lee�, Joo-Won Kim and Chan-Il Park�
Department of Marine Biology & Aquaculture, Institute of Marine Industry, College of Marine Science, Gyeongsang National University, 455, Tongyeong, 650-160, Korea
�Genetics and Breeding Research Center, National Fisheries Research and Development Institute, Geoje, 656-842, Korea
Natural-killer-cell-enhancing factor (NKEF) belongs to the newly defined peroxiredoxin (Prx) family. It was originally isolated from human erythroid cells. The black rockfish NKEF cDNA was identified through the expressed sequence tag (EST) analysis of PBLs libraries. The full-length NKEF cDNA was 1433 bp long and contained an open reading frame (ORF) of 594 bp that encoded 198 amino-acid residues. The 5’
UTR had a length of 39 bp, and the 3’UTR 800 bp. The deduced amino-acid sequence of the black rock- fish had a density 93.4, 92.9, 87.8, 85.8, 84.8, 83.8, 80.3, 79.7, 77.2, and 75.2% that of the pufferfish, olive flounder, channel catfish, zebrafish, chicken, common carp, Myotis lucifugus, cattle, human PrxI, rat PrxI, human NKEF-A, and Xenopus tropicalis, respectively. The NKEF gene was expressed in all the tissues of the black rockfish. The RT-PCR indicated that the NKEF transcripts were predominantly in the spleen and gill, less dominantly in the PBLs, head kidney, trunk kidney, and liver, and least in the intestine and mus- cles. This is the first report on the existence of the NKEF-A gene in black rockfish.
Key words: Black rockfish, Expressed sequence tag (EST), NKEF, RT-PCR
Mammalian natural killer (NK) cells and cytotox- ic T (Tc) cells play important roles in the surveil- lance and destruction of foreign or infected tissues and cells. NK cells, which are distinguished from Tc cells by the expression of Fc receptor III (CD16), CD56, and by the lack of a rearranged TCR (Shen et al., 2002), are able to kill certain tumor cells in an apparently non-MHC-restricted manner, without need for prior sensitization (Herberman et al., 1975, 1981). As such, NK cells do not recognize presented antigens, and their cytotoxicity occurs earlier than that of Tc cells. NK activity is also influenced by various cellular and humoral components in the blood (Shau et al., 1994).
Natural killer cell enhancing factor (NKEF) belongs to the newly defined peroxiredoxin (Prx) family. Itwas originally isolated and cloned from
human erythroid cells and named for its ability to enhance the cytotoxicity of NK cells against tumor cells (Shau et al., 1993). In addition to cytotoxicity, NKEF acts as a member of the peroxiredoxin (Prx) family and has an antioxidant function (Sauri et al., 1995). It increases the cellular resistance to oxidative damage by hydrogen peroxide and protects cells from alkyl hydroperoxide and heavy metals, such as methyl mercury (Kim et al., 1997). Many members belonging to the Prx family have been identified in plants, animals, and microorganisms (Chae et al., 1994). There are five subfamilies in the mammalian Prx proteins (Jin et al., 1997; Sato and Bannai, 1999). In humans, NKEF is a 44-kDa protein con- sisting of two same-sized subunits linked by disul- fide bonds. These two subunits, NKEF-A and NKEF-B, contain 199 and 198 amino acids, respec-
�Corresponding Author : Chan-Il Park, Tel : 82-55-640-3103 Fax : 82-55-642-4509, E-mail : [email protected]
16...
tively (~22 kDa), and share 88% amino-acid sequence identity and 71% nucleotide sequence identity (Shau et al., 1994). NKEF-A and NKEF-B, later identified as PrxI and PrxII, respectively (Shau and Kim, 1994), can be detected in a wide range of tissues (Kim et al., 1997; Schroder et al., 1998). The NKEF protein may also be involved in apoptosis (Sauri et al., 1996 Geiben-Lynn et al., 2003), cell proliferation, differentiation (Yamamoto et al., 1989;
Prosperi et al., 1993), and antiviral activity in vitro (Geiben-Lynn et al., 2003). Although recombinant NKEF (rNKEF)-A and recombinant NKEF (rNKEF)-B have similar levels of antioxidant func- tion, only the reduced form of rNKEF-A can enhance NK cytotoxicity, which indicates that both the antioxidant and NK-enhancing functions of rNKEF-A and rNKEF-B probably involve the cys- teine residues of the proteins but are mediated by sep- arate domains of the molecules (Sauri et al., 1996).
In fish, NKEF genes have been isolated and char- acterized in some species. In particular, the NKEF-A gene was sequenced from rainbow trout (Zhang et al., 2001), common carp (Shin et al., 2001), channel catfish (Li and Waldbieser, 2006), pufferfish (Dong et al., 2007), and olive flounder (Wang et al., 2005).
The NKEF amino-acid sequence is highly con- served between fish and mammals (Chen et al., 2006). All of them have an identical structure con- sisting of six exons and five introns (Shin et al., 2001; Zhang et al., 2001). The presence of the NKEF gene in black rockfish, however, has not been reported to date.
Black rockfish (Sebastes schlegelii) is one of the most significant fish species in Korea due to the human interest in aquaculture and fisheries, but dis- eases, especially infectious ones, have occurred fre- quently in them and have limited the production effi- ciencies of this economic fish. To improve the black rockfish production efficiency, it is necessary to
understand the cellular and molecular processes involved in the disease resistance to this species. The cloning and molecular characterization of the NKEF gene from black rockfish is very helpful in the study of the growth, disease resistance, temperature toler- ance, and physiological process of this species. In this paper, the cloning, characterization, and expres- sion analysis of the NKEF cDNA from black rock- fish are reported.
Materials and Methods
Cloning and sequencing of black rockfish NKEF A cDNA
The black rockfish NKEF full-length cDNA was identified via the EST analysis of the LPS stimulat- ed PBLs library (Baeck et al., 2008). Bioinformatic analysis was conducted to determine the gene iden- tities, using the Genetyx ver. 8.0 software (SDC Software, Japan). Briefly, the vector sequences were removed, and the database search was limited to the ESTs that are > 400 bp in length.
Sequence analysis and alignment
The sequence of NKEF cDNA was compared with the sequences in the peptide sequence databases at the National Centerfor Biotechnology Information (NCBI), using the BLAST network service.
Nucleotide sequence comparisons were carried out using the program BLASTX (Gish and David, 1993).
The multiple alignment of the amino-acid sequence of the NKEF proteins was made with the program Clustal W (Thompson et al., 1994). The sequences that were used for the comparison, and their GenBank accession numbers, were as follows:
olive flounder NKEF (AAY25400), pufferfish NKEF (AAY21814), channel catfish NKEF (AAU29515), common carp NKEF (BAA32086), zebrafish NKEF (AAI52139), chicken NKEF
Joo-Won Kim, Kwan Yong Jun, Geun Hee An, Hyung-Jun Park and Chan Il Park 344
Fig. 1. The cDNA and deduced amino acid sequence of black rockfish NKEF-A. Oligonucleotide primers used in the study are indicated with arrows, conserved cysteine residues are indicated by boxed-black. VCP motifs are with boxed and the polyadenylation signal AAUAAA is underlined, and poly (A) is marked with boldface.
(XP_422437), human NKEF-A (L19184), human PrxI (NP_002565), cattle PrxI (NP_776856), Myotis lucifugus PrxI (Q6B4U9), Norway rat PrxI (1QQ2_A), and Xenopus tropicalis PrxI (NP_001011135).
The phylogenetic relationships were inferred using the Mega 4 program and distance analysis through the neighbor-joining method. The values supporting each node were derived from 2,000 resamplings.
RT-PCR analysis of the NKEF transcripts in dif- ferent tissues
Eight tissues (PBLs, head kidney, trunk kidney, spleen, liver, intestine, gill, and muscle) were isolated from about 440-g healthy black rockfish. Total RNA from each tissue was extracted using TRIzol reagent (Invitrogen, USA), according to the manufacturer’s instructions. First-strand cDNA synthesis was carried out using a first-strand cDNA synthesis kit (Takara, Japan), according to the manufacturer’s instructions.
The first-strand cDNAs were used as a PCR amplifi- cation template, with the specific primers. The oligonucleotide primers used to amplify NKEF cDNA fragment were NKEF-F (5’-TTCAGT- GACGCTGCCGACGACT-3’) and NKEF-R (5’- GATGGTGATCTGTCTCAGGA-3’). Thermal cycling was performed using ExPrime Taq premix (GENET BIO, Korea) as follows: predenaturation at 94℃ for 5 min, 25 cycles of denaturation at 94℃ for 1 min, annealing at 60℃ for 30 s, extension at 72℃
for 40 s, and final extension at 72℃ for 7 min. The β- actin mRNA was used as the house keeping gene.
The primers β-actin-F (5’-TTTCCCTC- CATTGTTGGTCG-3’) and β-actin-R (5’- GCGACTCTCAGCTCGTTGTA-3’) were used for amplifying β-actin. A fragment of about 200 bp was amplified, and 10 ㎕PCR amplification products were observed on 1.5% agarose gel electrophoresis
with a 100-bp DNA ladder (Takara, Japan).
Results
Cloning and sequencing of back rockfish NKEF A cDNA
The full-length NKEF cDNA was 1433 bp long and contained an open reading frame (ORF) of 594 bp that encoded 198 amino acid residues. The 3’ UTR contains a polyadenylation signal (AATAAA) and a polyadenylation (accession number:
AB490894) (Fig. 1).
Sequence alignment and phylogenetic analysis The deduced amino-acid sequence of black rock- fish NKEF had a 93.4% identity with pufferfish NKEF-A, 92.9% with olive flounder NKEF, 87.8%
with channel catfish NKEF, 85.8% with zebrafish NKEF, 84.8% with chicken NKEF, 83.8% with common-carp NKEF, 80.3% with Myotis lucifugus PrxI, 79.7% with cattle PrxI, human Prx1, and rat Prx I, 77.2% with human NKEF-A, and 75.2% with Xenopus tropicalis(data not shown). The amino- acid sequence analysis indicated the existence of two consensus Val-Cys-Pro (VCP) motifs (amino acids 51-53 and 172-174) and three consensus cys- teine residues (Cys-52, Cys-71, and Cys-173).
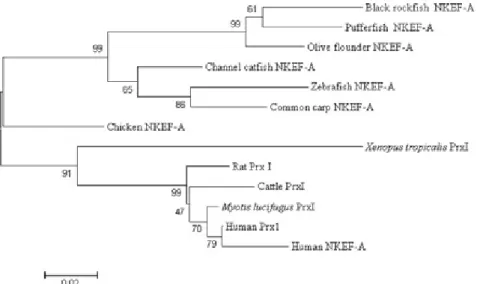
Among the three cysteine residues, Cys-52 and Cys-173 were involved in the VCP motifs, and Cys-71 was found in the N-terminus (Fig. 2). The phylogenetic analysis indicated that black rockfish NKEF forms an NKEF-A-like group and is closest to that of pufferfish NKEF among the examined species (Fig. 3).
Expression of the NKEF gene in different tissues The expression of the NKEF gene in the black rockfish tissues was detected via RT-PCR. The
Joo-Won Kim, Kwan Yong Jun, Geun Hee An, Hyung-Jun Park and Chan Il Park 346
Fig. 2. Multiple alignment of the deduced amino acid sequence of Black rockfish NKEF-A with the homologues in other species. Amino acids identical to the black rockfish sequence are indicated by an asterisk (*), the well-conserved cysteine residues are indicated by boxed-black and VCP motifs are with boxed.
NKEF gene was expressed in all the tissues of the black rockfish examined. The expression was found to be predominantly in the spleen and gill, less dominantly in the PBLs, head kidney, trunk kidney, and liver, and least in the intestine and muscle after 25 cycles of PCR (Fig. 4).
Discussion
Natural killer cells in mammals and natural-killer-
like cells in fish are important effectors of the innate immune response. They may be essential for prim- ing the adaptive immune response that plays an important role in the defense against pathogens (Kuznetsov, 1996). NKEF was first isolated from human red blood cells. It has the ability to enhance the cytotoxic activity of natural killer cells, and plays an important role in preventing DNA and pro- tein from being damaged by oxidative stress within living cells (Chen et al., 2006). In humans, NKEF is
Joo-Won Kim, Kwan Yong Jun, Geun Hee An, Hyung-Jun Park and Chan Il Park 348
Fig. 3. Neighbor-joining tree of Black rockfish NKEF-A constructed with Mega4. The bootstrap confidence values shown at the nodes of the tree are based on 2000 bootstrap 2000 replications. GenBank accession numbers for the sequences used are listed in Fig. 2.
Fig. 4. RT-PCR analysis of NKEF gene expression in various tissues of healthy black rockfish. β-actin was used as control.
Marker(M) indicates a 100-bp ladder marker.
encoded by two genes, NKEF-A and NKEF-B, which share an 88% amino-acid sequence identity and a 71% nucleotide sequence identity (Shau et al., 1994). Both NKEF-A and NKEF-B are induced by oxidative stress and have antioxidant activity, but only the reduced form of NKEF-A is able to enhance NK cytotoxicity (Sauri et al., 1995). In this study, full-length NKEF cDNA was isolated and characterized in black rockfish. Only one NKEF gene was obtained from the PBLs cDNA library of black rockfish, via EST analysis, which is similar with previous findings in the rainbow trout (Zhang et al., 2001), turbot (Chen et al., 2006), and channel catfish (Li et al. 2006).
In this study, the multiple alignments of amino acids showed that black rockfish NKEF shares a high sequence identity with the other known teleost NKEFs and vertebrate PrxIs. Especially, black rockfish NKEF contains two consensus Val-Cys- Pro (VCP) motifs, which the antioxidant activity of NKEF homologues critically depends on (Wood et al., 2003). Furthermore, black rockfish NKEF con- tains three cysteine residues (Cys-52, Cys-71, and Cys-173) that are conserved in all known NKEF-As and PrxIs, including those of pufferfish and olive flounder (Fig. 2). These results suggest that these three cysteine residues are essential in their func- tion. In addition, black rockfish contains only three cysteine residues, whereas the channel catfish (Ictalurus punctatus), zebrafish (Danio rerio), and common carp (Cyprinus carpio) NKEF-As contain an additionalcysteine residue (Cys-83). As such, the channel catfish, zebrafish, and common carp NKEF-As may potentially form two disulfide while only one disulfide bond can be formed in black rockfish NKEF. This is congruent with the report on pufferfish (Dong et al., 2007). The phylogenetic comparison of the NKEF sequences showed that black rockfish NKEF is closest to pufferfish NKEF-
A, and subsequently is more similar to the olive flounder genes than to the NKEFs of other teleosts and vertebrates (Fig. 3).
RT-PCR indicated that the NKEF gene was expressed universally in the tissues. The NKEF gene can be expressed in all the tissues of black rockfish, a phenomenon that is true as well for channel catfish (Li and Waldbieser, 2006), rainbow trout (Zhang et al., 2001), and carp (Shin et al., 2001). Huang et al. (2009) reported that the expres- sion level of the NKEF-A gene in the gill is the highest among the examined tissues where the carp had an uninfected condition with the Spring viraemia of the carp virus (SVCV). It coincided with the finding in the present study. The spleen is well known to be an important hematopoietic and lymphoid organ in fish, and is considered the source of immune cells, including lymphocytes, granulo- cytes, macrophages and some immune cytokines.
Thus, the higher expression level of the NKEF gene in the black rockfish spleen suggests that the teleost spleen is involved in the innate immune system.
Dong et al. (2007) reported that Tetraodon NKEF- A and NKEF-B, and of the change in the expression level after in-vivo stimulation with lipopolysaccha- ride (LPS), revealed that in the LPS-stimulated group, the expression levels of NKEF-A and NKEF-B were significantly increased in the spleen.
These results support the contention that LPS stim- ulation leads to the NKEF expression of black rock- fish. It must be determined in future researches, however, if they are sensitive not only to LPS but to other stimulants as well.
Through further investigation, these researchers intend to construct the recombinant NKEF of black rockfish so as to observe the localization and distri- bution of the NKEF-positive cells in black rockfish.
In addition, the effect of the black-rockfish rNKEFs and the possibility of developing immune activators
will be confirmed through in-vitro and in-vivo experiments.
Acknowledgement
This work was supported by a project grant from Yeongnam Sea Grant, Korea.
References
Baeck, G.W., Kim, J.W., Kim, K.H., Jun, K.Y., An, G.H. and Park, C.I.: Expressed Sequence Tags Analysis of Black Rockfish (Sebastes schlegelii) Peripheral Leukocytes Stimulated with Con A/PMA or LPS. J Fish Pathol., 21:129-137, 2008.
Baeck, G.W., Kim, J.W. and Park, C.I.: Identifica- tion and expression analysis of an interferon stimulated gene 15 (ISG15) from black rockfish, Sebastes schlegelii. Fish Shellfish Immunol., 25:679-81, 2008.
Chae, H.Z., Robison, K., Poole, L.B., Church, G., Storz, G. and Rhee, S.G.: Cloning and sequencing of thiol-specific antioxidant from mammalian brain: alkyl hydroperoxide reductase and thiol specific antioxidant define a large family of antioxidant enzymes. Proc. Natl. Acad. Sci. U.S.A.
91:7017-7021, 1994.
Chen, Y., Zhang, Y.X., F, T.J., Meng, L., Ren, G.C.
and Chen, S.L.: Molecular identification and expression analysis of the natural killer cell enhancing factor (NKEF) gene from turbot (Scophthalmus maximus) Aquaculture, 261:1186-1193, 2006.
Dong, W.R., Xiang, L.X. and Shao, J.Z.: Cloning and characterisation of two natural killer enhancing factor genes (NKEF-A and NKEF-B) in pufferfish, Tetraodon
nigroviridis. Fish Shellfish Immunol., 22:1- 15, 2007.
Geiben-Lynn, R., Kursar, M., Brown, N.V., Addo, M.M., Shau, H. and Lieberman, J.: HIV-1 antiviral activity of recombinant natural killer cell enhancing factors, NKEF-A and NKEF-B, members of the peroxiredoxin family. J Biol Chem., 278:1569-1574, 2003.
Gish, W. and David, J.S.: Identification of protein coding regions by database similarity search.
Nature Genetics, 3:266-272, 1993.
Herberman, R.B., Nunn, M.E., Holden, H.T. and Lavrin, D.H.: Natural cytotoxic reactivity of mouse lymphoid cells against syngeneic and allogeneic tumors. II. Characterization of effector cells. Int J Cancer, 16:230-239, 1975.
Herberman, R.B. and Ortaldo, JR.: Natural killer cells: their roles in defenses against disease.
Science, 214:24-30, 1981.
Jin, D.Y., Chae, H.Z., Rhee, S.G. and Jeang, K.T.:
Regulatory role for a novel human thiore- doxin peroxidase in NF-RB activation. J.
Biol. Chem., 272:30952-30961, 1997.
Kim, A.T., Sarafian, T.A. and Shau, H.: Characteri- zation of antioxidant properties of natural killer-enhancing factor-B and induction of its expression by hydrogen peroxide. Toxi- cology and Applied Pharmacology, 147:135-142, 1997.
Kim, H., Lee, T.H., Park, E.S., Suh, J.M. and Park, S.J., Chung, H.K.: Role of peroxiredoxins in regulating intracellular hydrogen peroxide and hydrogen peroxide-induced apoptosis in thyroid cells. J Biol Chem., 275:18266- 18270, 2000.
Kuznetsov, V.A.: ‘‘Harpoon’’model for cell-cell adhesion and recognition of target cells by the natural killer cells. Journal of Theoretical
Joo-Won Kim, Kwan Yong Jun, Geun Hee An, Hyung-Jun Park and Chan Il Park 350
Biology, 180:321-342, 1996.
Li, R.W. and Waldbieser, G.C.: Genomic organisa- tion and expression of the natural killer cell enhancing factor (NKEF) gene in channel catfish Ictalurus punctatus(Rafinesque).
Fish Shellfish Immunol., 20:72-82, 2006.
Prosperi, M.T., Ferbus, D., Karczinski, I. and Goubin, G.: A human cDNA corresponding to a gene overexpressed during cell prolifer- ation encodes a product sharing homology with amoebic and bacterial proteins. J Biol Chem., 268:11050-11056, 1993.
Sato, H. and Bannai, S.: Peroxiredoxin: a new host antioxidation system. Seikagaku 71:333- 337, 1999.
Sauri, H., Butterfield, L., Kim, A. and Shau, H.:
Antioxidant function of recombinant human natural killer enhancing factor. Biochemical and Biophysical Research Communications, 208:964-969, 1995.
Saur,i H., Ashjian, P.H., Kim, A.T. and Shau, H.:
Recombinant natural killer enhancing factor augments natural killer cytotoxicity. J Leukoc Biol., 59:925-931, 1996.
Schroder, E., Willis, A.C. and Ponting, C.P.:
Porcine natural killer enhancing factor-B:
oligomerisation and identification as a cal- pain substrate in vitro. Biochim Biophys Acta., 1383:279-291, 1998.
Shau, H., Gupta, R.K. and Golub, S.H.: Identifica- tion of natural killer enhancing factor (NKEF) from human erythroid cells. Cell.
Immunol., 147:1-11, 1993.
Shau, H., Butterfield, L.H., Chiu, R. and Kim, A.:
Cloning and sequence analysis of candidate human natural killer enhancing factor genes.
Immunogenetics, 40:129-134, 1994.
Shau, H. and Kim, A.: Identification of natural killer enhancing factor as a major antioxi-
dant in human red blood cells. Biochem Bio- phys Res Commun., 199:83-88, 1994.
Shen, L., Stuge, T.B., Zhou, H., Khayat, M., Barker, K,S. and Quiniou, S.M.: Channel catfish cytotoxic cells: a mini-review. Dev Comp Immunol., 26:141-149, 2002.
Shin, D.H., Fujiki, K., Nakao, M. and Yano, T.:
Organization of the NKEF gene and its expression in the common carp (Cyprinus carpio). Dev. Comp. Immunol., 25:597-606, 2001.
Thompson, J.D., Higgis, D.G. and Gibson, T.J.:
CLUSTAL W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position specif- ic gap penalties and weight matrix choice.
Nucleic Acids Res., 22:4673-4680, 1994.
Wang, Z., Chen, S.L. and Ji, X.S.: Cloning and expression analysis of natural killer enhanc- ing factor (NKEF) from Japanese flounder (Paralichthys olivaceus). Chin. High Tech- nol. Lett., 15:86-90, 2005.
Wood, Z.A., Schrode,r E., Robin-Harris, J. and Poole, L.B.: Structure, mechanism and regu- lation of peroxiredoxins. Trends Biochem Sci., 28:32-40, 2003.
Yamamoto, T., Matsui, Y., Natori, S. and Obinata, M.: Cloning of a housekeeping-type gene (MER5) preferentially expressed in murine erythroleukemia cells. Gene, 80:337-343, 1989.
Zhang, P., Liu, B., Kang, S.W., Seo, M.S., Rhee, S.G. and Obeid, L.M.: Thioredoxin peroxi- dase is a novel inhibitor of apoptosis with a mechanism distinct from that of Bcl-2. J Biol Chem., 272:30615-30618, 1997.
Zhang, H., Evenhuis, J.P., Thorgaard, G.H. and Ris- tow, S.S.: Cloning, characterization and genomic structure of the natural killer cell
enhancement factor (NKEF)-like gene from homozygous clones of rainbow trout. Dev.
Comp. Immunol., 25:25-35, 2001.
Manuscript Received : September 15, 2009 Revised : December 18, 2009 Accepted : December 24, 2009 Joo-Won Kim, Kwan Yong Jun, Geun Hee An, Hyung-Jun Park and Chan Il Park
352