ABSTRACT
BACKGROUND/OBJECTIVES: The present study aimed to further investigate the potential health beneficial effects of long-term seaweed supplementation on lipid metabolism and hepatic functions in DIO mice.
MATERIALS/METHODS: Four brown seaweeds (Undaria pinnatifida [UP], Laminaria japonica [LJ], Sargassum fulvellum [SF], or Hizikia fusiforme [HF]) were added to a high fat diet (HFD) at a 5% ratio and supplemented to C57BL/6N mice for 16 weeks. Triglycerides (TGs) and total cholesterol (TC) in the liver, feces, and plasma were measured. Fecal bile acid (BA) levels in feces were monitored. Hepatic insulin signaling- and lipogenesis-related proteins were evaluated by Western blot analysis.
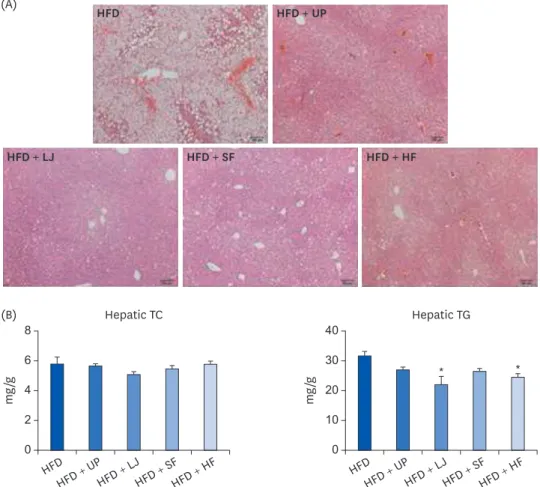
RESULTS: Fasting blood glucose levels were significantly reduced in the LJ, SF, and HF groups compared to the HFD group by the end of 16-week feeding period. Plasma TG levels and hepatic lipid accumulation were significantly reduced in all 4 seaweed supplemented groups, whereas plasma TC levels were only suppressed in the UP and HF groups compared to the HFD group. Fecal BA levels were significantly elevated by UP, LJ, and SF supplementation compared to HFD feeding only. Lastly, regarding hepatic insulin signaling-related proteins, phosphorylation of 5′-AMP-activated protein kinase was significantly up-regulated by all 4 types of seaweed, whereas phosphorylation of protein kinase B was up-regulated only in the SF and HF groups. Lipogenesis-related proteins in the liver were effectively down-regulated by HF supplementation in DIO mice.
CONCLUSIONS: Brown seaweed consumption showed hypotriglyceridemic effects in the prolonged DIO mouse model. Specifically, combinatory regulation of BA excretion and lipogenesis-related proteins in the liver by seaweed supplementation contributed to the reduction of plasma and hepatic TG levels, which inhibited hyperglycemia in DIO mice. Thus, the discrepant and species-specific functions of brown seaweeds provide novel insights for the selection of future targets for therapeutic agents.
Keywords: Seaweed; liver; lipogenesis; bile; mice
Original Research
Received: Mar 12, 2020 Revised: Jun 3, 2020 Accepted: Jul 21, 2020
§
Corresponding Author:
Yunkyoung Lee
Department of Food Science and Nutrition, Jeju National University, 9-431, Natural Science Building I, 102 Jejudaehak-ro, Jeju 63243, Korea.
Tel: +82-64-745-3555 Fax: +82-64-725-2539 E-mail. [email protected]
©2020 The Korean Nutrition Society and the Korean Society of Community Nutrition This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (https://
creativecommons.org/licenses/by-nc/4.0/) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
ORCID iDs A-Reum Han
https://orcid.org/0000-0002-5143-797X Jae-Hoon Kim
https://orcid.org/0000-0002-4410-9126 Eunyoung Kim
https://orcid.org/0000-0001-9018-1797 Jiamei Cui
https://orcid.org/0000-0002-9126-9667 In-Suk Chai
https://orcid.org/0000-0002-2673-9829 Guiguo Zhang
https://orcid.org/0000-0002-0515-2172 Yunkyoung Lee
https://orcid.org/0000-0001-6453-769X
A-Reum Han 1,2 , Jae-Hoon Kim 3 , Eunyoung Kim 1 , Jiamei Cui 1 , In-Suk Chai 1 , Guiguo Zhang 4 , and Yunkyoung Lee 1§
1
Department of Food Science and Nutrition, Jeju National University, Jeju 63243, Korea
2
Yerae Elementary School, Jeju 63537, Korea
3
College of Veterinary Medicine, Jeju National University, Jeju 63243, Korea
4