In this study interactions between antimicrobial substances produced by Paenibacillus elgii DS381, Burkholderia gladioli DS518, Streptomyces lienomycini DS620, and Paenibacillus elgii DS1515 were examined against various microorganisms including pathogenic bacteria and toxigenic fungi. When the interactions between antimicrobial substances from bacteria were investigated, several combinations of antimicrobial sub- stances displayed a synergistic effect against each different target organisms (0 < fractional inhibitory concentration index
< 0.75). In a time-kill assay, most combinations of antimicrobial substances showed a synergistic effect reducing more than 10
5cell/ml of bacteria and yeast during 24 h. Many combinations of antimicrobial substances also increased the degradation of spore of toxigenic fungi up to 100% compared to those by individual antimicrobial substances, and completely inhibited the germination of the residual spores. These findings have potential implications in delaying the development of resistance by harmful microbes against antimicrobial substances as the antimicrobial effect can be achieved with lower concentrations of antimicrobial substances.
Keywords: antimicrobial activity, antimicrobial substances, fractional inhibitory concentration index, synergistic effect
유기물이 주 원료로 이용되는 식품 및 화장품 산업에서는 미생물 증식으로 인한 제품의 변질, 부패 및 인간 질병 발생을 막기 위해 다양한 화학 또는 천연 방부소재를 첨가물로 사용 하고 있다. 하지만 미생물 제어를 위해 사용되는 화학 방부제 의 독성과 부작용이 문제가 되고 있을 뿐만 아니라(White and Groot, 2006), 항균소재에 대해 빈번하게 미생물의 내성이 발 생하며(Regmi et al., 2017), 천연 항균물질도 고농도로 식품 에 처리 시 인간 세포에 독성을 보일 수 있다고 보고되었다 (Aranha et al., 2004). 따라서 최근에는 새로운 항균물질을 찾 거나(Regmi et al., 2017), 항균물질을 조합하여 처리하는 방법 이 제시되었다(Magalhães and Nitschke, 2013). 그 중 두 가지 이상의 항균물질 조합 처리는 항균물질의 처리 농도, 물질의 반응 시간 및 부작용을 줄일 수 있는 장점이 있으며 이런 방식 을 다중 장애물 접근법(multiple hurdle approach)이라고 한다.
또한 항균물질 각각을 단독으로 처리했을 때보다 두 가지 이 상의 항균물질을 조합하였을 때 표적 미생물에 더 높은 효과 를 발휘하는 시너지(synergy) 효과도 일어날 수 있다(Regmi et al., 2017).
이전의 연구에서는 항균물질 활성 감소 문제를 해결하기 위해 식품 유래 병원체에 대한 ε-poly-L-lysine과 nisin 조합 (Najjar et al., 2009), nisin A와 polymyxin B (Naghmouchi et al., 2010)의 조합 그리고 Bacillus subtilis 유래 antimicrobial peptides 조합 처리의 시너지 효과를 보고하였다(Regmi et al., 2017). 이러한 항균물질의 조합은 빠르고 지속적으로 대상 미
세균 유래 항균물질의 다양한 미생물에 대한 항균활성의 상호작용
이다솔 ・ 송홍규*
강원대학교 생명과학과
Interactions between antimicrobial substances from bacteria against various microorganisms
Da-Sol Lee and Hong-Gyu Song*
Department of Biological Sciences, Kangwon National University, Chuncheon 24341, Republic of Korea (Received July 7, 2020; Revised July 24, 2020; Accepted August 10, 2020)
*For correspondence. E-mail: [email protected];
Tel.: +82-33-250-8545; Fax: +82-33-259-5665
생물의 사멸을 유도하였다. 하지만 대부분의 연구가 항생제, 식물 추출물과 nisin에 초점이 맞춰져 있기 때문에 본 연구에 서는 기존에 보고되지 않은 세균 유래 항균물질들 간의 다양 한 미생물에 대한 상호작용을 조사하였다.
재료 및 방법
항균활성 세균 및 조사대상 미생물
춘천과 평창의 토양에서 분리한 Paenibacillus elgii DS381, Burkholderia gladioli DS518, Streptomyces lienomycini DS620 과 Paenibacillus elgii DS1515를 실험에 이용하였는데 이 네 종류의 세균은 여드름 균, 인간 피부상재균, 병원성 세균 및 독소생성 진균에 대한 항균활성이 보고된 바 있다(Lee and Song, 2018a, 2018b, 2019a, 2019b). 특히 DS381과 DS1515는 lipopeptide성 계면활성 물질, DS518은 protease, 그리고 DS620 균주는 anthracyclic antibiotics 계열 중 daunomycinone이 주 된 항세균 및 항진균물질로 작용하였다(Lee and Song, 2018a).
균주의 항균물질은 Lee와 Song (2018a)에 보고된 내용과 동일 한 방법으로 추출하였다. DS381, DS518 및 DS1515 균주의 항균물질은 배양 상등액에 ammonium sulfate를 첨가하여 침 전시킨 단백질을 동결 건조한 뒤 이용하였고, DS620 균주의 경우 배양액을 ethyl acetate 용매로 추출한 뒤 유기용매층을 감압 건조하여 얻은 분말을 항균물질로서 이용하였다. 1.5 ml tube에 각각의 균주 항균물질 400 mg을 넣고 멸균 증류수 또는 methanol을 첨가하여 용해한 뒤(v/v) 2배씩 연속 희석하여 농 도 별로 이용하였다.
이 항균물질들은 다양한 인간 피부상재균, 병원성 세균 및 독 소생성 진균에 대하여 낮은 최소저해농도(minimum inhibitory concentration, MIC)를 나타내었는데 그 범위는 0.0078~10,000 µg/ml로 우수한 항균활성을 나타내었다(Lee and Song, 2018a, 2018b, 2019a, 2019b).
각 균주가 생성하는 주된 항균물질을 조합한 혼합물의 다 음 미생물들에 대한 항균활성을 조사하였다. 화장품업계에서 주된 오염 대상 미생물로 조사하는 피부 상재균 6종[Candida albicans (American Type Culture Collection: ATCC)10231, B. subtilis ATCC19659, S. aureus ATCC6538, Aspergillus niger ATCC16404, Pseudomonas aeruginosa (Korean Collection for Type Cultures: KCTC)2513과 E. coli ATCC8739], 경상대학 교 병원체 자원은행(Gyeongsang National University Hospital Branch of the National Culture Collection for Pathogens:
GNUH-NCCP)으로부터 분양받은 병원성 세균[Enterococcus
faecalis GNUH-NCCP (3728, 3738과 3788), Micrococcus luteus GNUH-NCCP (2837, 2922와 3683), Staphylococcus epidermidis GNUH-NCCP (44, 579와 672), Listeria monocytogenes GNUH- NCCP (2148, 2637과 2868), Klebsiella pneumoniae GNUH- NCCP (29, 4149와 4159), Propionibacterium acnes GNUH- NCCP (2875와 2876)], 국립농업과학원 미생물은행(Korean Agricultural Culture Collection: KACC)으로부터 분양받은 aflatoxin B
1생성 Aspergillus flavus KACC (44986, 45068과 45146), fumonisin B
1생성 Fusarium fugikuroi KACC (46888 과 48352), Fusarium proliferatum KACC48354와 Fusarium verticillioides KACC48356 및 한국미생물 보존센터(Korean Culture Center of Microorganisms: KCCM)로부터 분양받은 ochratoxin 생성 Aspergillus alutaceus KCCM60421과 Aspergillus awamori KCCM32316을 연구에 이용하였다.
Checkerboard assay
MIC 조사 방법을 기반으로 한 항균물질 간 상호작용 효과 는 Sopirala 등(2010)과 Fadli 등(2012)의 checkerboard assay 방법으로 조사하였다. 대상 미생물은 tryptic soy broth (TSB, Difco Lab) 또는 potato dextrose broth (PDB, Difco Lab) 배지 에 배양하고 개체수를 10
7cell/ml로 보정하여 이용하였으며, 항균물질은 다양한 농도(⅛MIIC~4MIC)로 준비하였다. 96 well plate에 각 균주 생장에 적합한 TSB 또는 PDB 배지 160 µl, A 항균물질 10 µl, B 항균물질 10 µl, 균 배양액 20 µl 를 첨 가 후 배양하였다(30~37°C, 24~48시간). 배양 후 균 생장이 나 타나지 않은 처리구에 대해서는 아래와 같은 식에 근거하여 FICI (fractional inhibitory concentration index)를 구해 항균물 질 간 상호작용을 조사하였다. FICI가 0.5 이하일 때 전체 시너 지, 0.5~0.75 일 때 부분적 시너지, 0.75~2 이하일 때 독립적 활 성, 2 보다 클 때는 적대적 활성으로 평가하였다.
FICI = 항균 물질 혼합 처리구에서 A 항균물질 MIC / A 항균 물질 단독 처리구 MIC +
항균 물질 혼합 처리구에서 B 항균물질 MIC / B 항균물질 단독 처리구 MIC
Time kill assay
항균물질 간의 시너지 효과를 검정하기 위한 time kill assay
는 Jayaraman 등(2010)과 Al-Ani 등(2015)의 방법으로 수행하
였다. 대상 미생물은 TSB 배지에 배양(30°C, 24시간)하고 개
체수를 10
5~10
6cell/ml로 보정하여 이용하였다. 균 배양액 20
ml를 50 ml conical tube에 담고 항균물질을 항균물질 비처 리구, A 항균물질(½MIC), B 항균물질(½MIC), A 항균물질 (½MIC) + B 항균물질(½MIC)와 같이 다양한 조합으로 나누 어 처리하고 배양하였다. 이후 0, 4, 8과 24시간 간격으로 시료 를 수집한 뒤 10배 단위로 희석하여 TSA 배지에 도말하였다.
배양(30~37°C, 24~48시간) 후 생긴 집락을 계수하였고 A 항 균물질(½MIC)과 B 항균물질(½MIC)을 동시에 처리한 시료 에서 균 개체수가 2 log CFU/ml 이하로 나타날 때 시너지 효과 가 나타났다고 판단하였다. 다양한 시너지 조합 처리 중에서 E. coli ATCC8739를 대상으로 항균물질 처리 후 균의 형태와 수를 한국기초과학지원연구원 춘천센터에 의뢰하여 투과 전 자현미경으로 관찰하였다. A 항균물질(½MIC) + B 항균물질 (½MIC) 조합과 항균물질 미처리 대조구로 실험을 진행하였 으며 항균물질 처리 6시간 후의 시료를 glutaraldehyde로 고정 한 뒤 시료 박편을 제작하여 관찰하였다.
독소생성 진균 포자의 분해 및 발아 저해
독소생성 진균의 포자 발아 저해에 대한 세균 항균물질 조
합 처리의 효과를 조사하기 위해 Veras 등(2016)의 방법을 수 정하여 이용하였다. 1.5 ml tube에 0.05% tween 80을 이용하여 회수한 진균 포자 현탁액(10
7spores/ml) 100 µl, A 항균물질이
½MIC로 첨가된 15% yeast extract saccharose (YES) 배지 50 µl, B 항균물질이 ½MIC로 첨가된 15% YES를 첨가하였다. 대 조구는 항균물질을 첨가하지 않은 15% YES 배지를 100 µl 첨 가하였다. 모든 실험구는 30°C에서 24시간 동안 배양하였고, 광학 현미경을 이용하여 포자 분해와 잔여 분생포자(conidia) 의 발아를 관찰하였다.
결과 및 고찰
Checkerboard assay
항균물질 간 상호작용은 세균 균주 항균물질 간의 조합으 로 조사하였다. MIC 조사 방법을 기반으로 한 checkerboard assay는 특정 항균물질 단독을 1MIC로 대상 미생물에 처리했 을 때보다 두 가지 이상의 항균물질을 동시에 처리했을 때 항
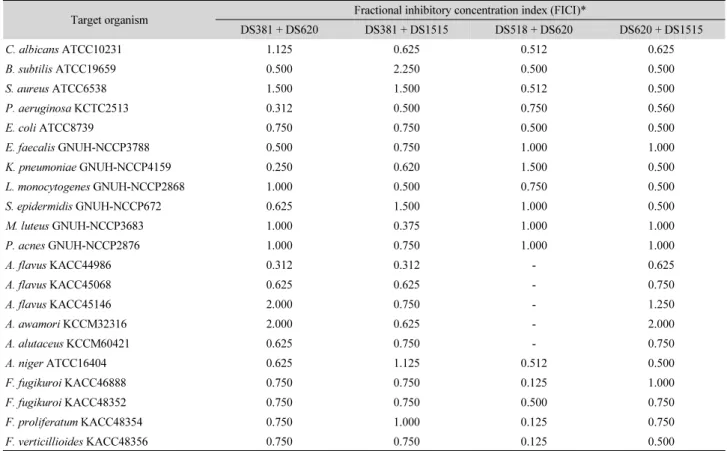
Table 1. Interactions of antimicrobial activity of antimicrobial substances combination from strains DS381, DS518, DS620, and DS1515 in a checkerboard assay against diverse microorganisms
Target organism Fractional inhibitory concentration index (FICI)*
DS381 + DS620 DS381 + DS1515 DS518 + DS620 DS620 + DS1515
C. albicans ATCC10231 1.125 0.625 0.512 0.625
B. subtilis ATCC19659 0.500 2.250 0.500 0.500
S. aureus ATCC6538 1.500 1.500 0.512 0.500
P. aeruginosa KCTC2513 0.312 0.500 0.750 0.560
E. coli ATCC8739 0.750 0.750 0.500 0.500
E. faecalis GNUH-NCCP3788 0.500 0.750 1.000 1.000
K. pneumoniae GNUH-NCCP4159 0.250 0.620 1.500 0.500
L. monocytogenes GNUH-NCCP2868 1.000 0.500 0.750 0.500
S. epidermidis GNUH-NCCP672 0.625 1.500 1.000 0.500
M. luteus GNUH-NCCP3683 1.000 0.375 1.000 1.000
P. acnes GNUH-NCCP2876 1.000 0.750 1.000 1.000
A. flavus KACC44986 0.312 0.312 - 0.625
A. flavus KACC45068 0.625 0.625 - 0.750
A. flavus KACC45146 2.000 0.750 - 1.250
A. awamori KCCM32316 2.000 0.625 - 2.000
A. alutaceus KCCM60421 0.625 0.750 - 0.750
A. niger ATCC16404 0.625 1.125 0.512 0.500
F. fugikuroi KACC46888 0.750 0.750 0.125 1.000
F. fugikuroi KACC48352 0.750 0.750 0.500 0.750
F. proliferatum KACC48354 0.750 1.000 0.125 0.750
F. verticillioides KACC48356 0.750 0.750 0.125 0.500
* Total synergy (FICI ≤ 0.5), partial synergy (0.5 < FICI ≤ 0.75), independent (0.75 < FICI ≤ 2), antagonism (FICI > 2).
균물질의 처리 농도가 감소하게 되면 시너지가 나타났다고 평 가하는 방법이다. 이 때 이 척도를 나타내는 FICI가 0.5 이하, 즉 항균물질 A (¼MIC) + 항균물질 B (¼MIC) 처리 시 각 항균 물질 1MIC와 동일한 효과를 나타나면 시너지가 나타났다고 평가할 수 있고, 그 보다는 높은 0.75일 때 부분적인 시너지가 나타났다고 평가할 수 있다(Sopirala et al., 2010; Fadli et al., 2012).
세균 균주 항균물질 간 조합을 다양한 세균과 진균에 처리 하였을 때 나타난 결과 중 가장 시너지 효과가 우수하였던 것 만을 Table 1에 제시하였다. 조사 결과 세균과 효모에 대해서 는 다양한 조합 중 DS381 + DS1515 및 DS620 + DS1515 균주 항균물질의 조합이 가장 우수하였다. 또한 DS620 + DS1515 항균물질 균주 조합에서 가장 광범위하게 전체 시너지가 나타 난 것은 두 균주 항균물질의 특성과 작용 방법이 다르기 때문이 라고 판단된다. 진균에 대해서는 DS381 + 1515 균주 항균물질 조합이 가장 폭 넓게 시너지를 나타내었으며, DS518 + DS620 조합은 Fusarium sp.에 가장 우수한 시너지 효과를 나타내었다.
항균물질 간 시너지 효과에 대한 이제까지의 보고는 대부분 식 물 추출물, nisin과 항생제에 초점이 맞춰져 있었다. 항생제 내 성 세균에 대한 식물의 essential oil과 항생제 간의 시너지(Lahmar et al., 2017), B. subtilis MTCC2756, S. aureus MTCC902, P.
aeruginosa MTCC2642와 E. coli MTCC2622에 대한 미생물 유래 항균물질인 stilbene 화합물(3,4',5-trihydroxystilbene과 3,5-dihydroxy-4-isopropylstilbene)과 항생제(ciprofloxacin과 cefotaxime) 간의 시너지 효과가 보고된 바 있다(Kumar et al., 2012). 또한 미생물 유래 항균물질인 ε-poly-L-lysine과 nisin의
시너지(Najjar et al., 2009)와 Bacillus subtilis 유래 antimicrobial peptides 간의 시너지 효과가 보고되었다(Regmi et al., 2017).
하지만 본 연구의 균주들이 생산하는 항균물질에 대해 다양한 미생물을 대상으로 한 시너지 효과에 대한 보고는 없었다.
Time kill assay
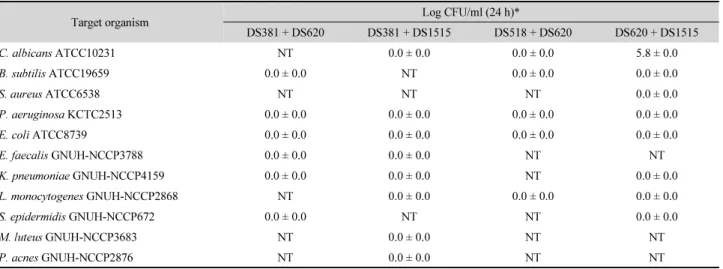
Checkerboard assay를 토대로 하여 세균 균주 항균물질을 조합하여 다양한 세균과 효모에 처리하였을 때 부분적 혹은 전체 시너지가 나타난 대부분의 처리구에서 균 개체수가 검출 되지 않는 시너지 양상을 나타내었다(Table 2). 세균 항균물 질의 조합 처리 시 나타난 사멸 효과에서는 DS381 + DS1515 균주 항균물질 조합과 DS620 + DS1515 균주 항균물질 조합 이 가장 우수하였다. 다양한 결과 중 피부 상재균에 대해 DS381 + DS1515 균주 항균물질 조합은 항균물질을 단독(½MIC) 으로 처리하였을 때와 항균물질 미처리 대조군의 세포수가 시간 경과에 따라 증가 또는 유지되는 양상과는 다르게 C.
albicans ATCC10231, P. aeruginosa KCTC2513과 E. coli ATCC8739를 각각 24, 8과 8시간 내에 사멸시키는 효과를 나 타내었다(Fig. 1). DS620 + DS1515 균주 항균물질의 조합은 최소 4시간 내에 P. aeruginosa KCTC2513를 사멸시키는 우 수한 시너지 항균효과를 나타내었고, E. coli ATCC8739는 8 시간 내에 사멸시키는 시너지 효과를 나타내었다(Fig. 2). 다양 한 병원성 세균에 대해서도 피부 상재균에 대한 결과와 유사하 게 항균물질 조합 처리 후 대상 미생물을 빠르게 사멸시키는 시 너지 효과를 나타내었다(결과 미제시). 이 두 항균물질 조합의 경우 모두 E. coli ATCC8739에 대해 부분적 혹은 전체 시너지
Table 2. Synergistic effect of combination of antimicrobial substances (½MIC) from strains DS381, DS518, DS620, and DS1515 in a time kill assay against diverse microorganisms
Target organism Log CFU/ml (24 h)*
DS381 + DS620 DS381 + DS1515 DS518 + DS620 DS620 + DS1515
C. albicans ATCC10231 NT 0.0 ± 0.0 0.0 ± 0.0 5.8 ± 0.0
B. subtilis ATCC19659 0.0 ± 0.0 NT 0.0 ± 0.0 0.0 ± 0.0
S. aureus ATCC6538 NT NT NT 0.0 ± 0.0
P. aeruginosa KCTC2513 0.0 ± 0.0 0.0 ± 0.0 0.0 ± 0.0 0.0 ± 0.0
E. coli ATCC8739 0.0 ± 0.0 0.0 ± 0.0 0.0 ± 0.0 0.0 ± 0.0
E. faecalis GNUH-NCCP3788 0.0 ± 0.0 0.0 ± 0.0 NT NT
K. pneumoniae GNUH-NCCP4159 0.0 ± 0.0 0.0 ± 0.0 NT 0.0 ± 0.0
L. monocytogenes GNUH-NCCP2868 NT 0.0 ± 0.0 0.0 ± 0.0 0.0 ± 0.0
S. epidermidis GNUH-NCCP672 0.0 ± 0.0 NT NT 0.0 ± 0.0
M. luteus GNUH-NCCP3683 NT 0.0 ± 0.0 NT NT
P. acnes GNUH-NCCP2876 NT 0.0 ± 0.0 NT NT
* Log CFU/ml < 2: synergy; NT, not tested.
를 보였기 때문에 항균물질 조합물 처리 후 세포수와 형태를 투 과 전자현미경으로 관찰하였다. 그 결과 배양 6시간 후 항균물 질을 처리하지 않은 대조군은 세포의 수가 많고 온전한 세포의 형태를 보였으나, DS381 + DS1515 그리고 DS620 + DS1515 항균물질 조합 처리구의 경우 세포의 형태가 온전하지 못하고 사멸하여 형태만 남은 경우가 대부분으로 시너지 효과에 대한 가시적인 결과를 관찰할 수 있었다(Fig. 3).
이러한 결과는 Hamoud 등(2014)의 연구에서 식물 유래 항 균물질인 thymol, 항생제 vancomycin 그리고 식품 방부제로 종종 이용되는 ethylenediaminetetraacetic acid (EDTA)의 다 양한 조합을 항생제 저항성 E. coli ATCC25922에 처리하였을 때 시간이 지남에 따라 대부분 조합 처리에서 개체수가 증가 하는 양상을 보인 것과 비교하여 우수하다고 할 수 있다. 또한 고농도의 thymol + EDTA + vancomycin 조합 처리 시 8시간 내 대상 미생물을 사멸시키지 못한 결과와 비교하여 본 연구 균주의 항균물질 조합 일부는 빠르게 E. coli를 포함한 여러 대 상 미생물을 제어하였다. 다른 E. coli 균주를 대상으로 한 Puño-Sarmiento 등(2020)의 연구에서 항생제 Ciprofloxacin + antimicrobial peptide 및 항생제 Meropenem + antimicrobial 조합을 shiga toxin 생성 E. coli O157:H7에 처리하였을 때 시 간이 지남에 따라 개체수가 감소하다가 배양 12시간부터 24 시간까지 다시 증가하는 양상을 보인 것과도 대조적인 결과를 나타내었다(Figs. 1 and 2). 또한 Aiyegoro 등(2009)의 연구에 서 밀집꽃속의 Helichrysum pedunculatum 잎 추출물과 항생 제 조합을 S. aureus ATCC6538에 처리하고 배양 24시간 후 계 (A)
(B)
(C)
Fig. 1. Population change of C. albicans ATCC10231 (A), P. aeruginosa KCTC2513 (B), and E. coli ATCC8739 (C) in the presence of anti- microbial substances of strains DS381 and DS1515. (X, antimicrobial substance of DS381 ½MIC + antimicrobial substance of DS1515 ½MIC;
◇, DS381 ½MIC; ○, DS1515 ½MIC; □, control).
(A)
(B)
(C)
(D)
Fig. 2. Population change of B. subtilis ATCC19659 (A), S. aureus ATCC6538 (B), P. aeruginosa KCTC2513 (C), and E. coli ATCC8739 (D) in the presence of antimicrobial substances of strains DS620 and DS1515. (X, antimicrobial substance of DS620 ½MIC + antimicrobial substance of DS1515 ½MIC; ○, DS620 ½MIC; ◇, DS1515 ½MIC; □, control).
수 시 개체수가 감소하지 않은 결과, 그리고 K. pneumoniae ATCC10031, B. subtilis와 S. epidermidis의 개체수를 대부분 1 에서 3 log CFU/ml 수준으로 감소시킨 것과 비교하여 우수한 사멸효과를 나타내고 있다. 추가적으로 Jayaraman 등(2010) 의 연구에서 다양한 항생제와 식물 생리활성물질을 각각
½MIC로 조합하여 P. aeruginosa에 처리하였을 때 배양 24시 간 후에도 개체수가 완전히 사멸하지 못한 결과와 비교하여 우수한 항균활성을 나타낸다고 할 수 있다.
독소생성 진균 포자의 분해 및 발아 저해
항균물질 조합의 독소생성 진균의 포자 분해와 발아 저해 율에 미치는 영향을 조사하기 위해서 checkerboard assay를 토 대로 세균 항균물질을 ½MIC로 다양하게 조합하여 진균 포자 에 처리하였다. 항균물질 조합 처리 시 상당수의 조합에서 checkerboard assay 결과와 유사하게 시너지 효과를 나타내어
Lee와 Song (2019a)이 보고한 동일한 항균 물질 단독 처리 시 나타난 결과와 비교하여 포자 분해나 발아 저해 정도가 증가 하였다(Table 3). Checkerboard assay에서 FICI가 0.75 이상으 로 시너지가 아닌 독립적 활성을 나타낸 조합은 포자 분해와 발아 저해를 조사하지 않았다. 특히 4개 균주 항균물질의 단독 처리 시에는 A. niger ATCC16404의 포자 분해가 13.1~30.6%
를 나타내었는데(Lee and Song, 2019a), 항균물질 조합 3개에 서 100% 내외의 포자 분해 증가를 보였으며, 모든 조합에서 대 부분 100%의 포자 발아 저해를 나타내었다(Table 3). 이 결과 를 현미경으로 직접 관찰하였을 때, 대조군의 포자는 대부분 발아하거나 존재하는 반면 항균물질 조합 처리구는 포자가 거 의 존재하지 않고 발아한 포자는 전혀 관찰되지 않았다(Fig. 4).
따라서 진균포자 제거는 물론 포자 발아 저해를 통해서 진균의 번식을 방지할 수 있을 것으로 기대된다. DS381, DS518과 DS1515 균주는 진균의 세포벽을 분해할 수 있는 chitinase 활성
(A) (B) (C)
Fig. 3. Transmission electron micrograph of E. coli ATCC8739 after treatment of antimicrobial substance (½MIC) combination [control (A), DS381 + DS1515 (B), and DS620 + DS1515 (C)].
Table 3. Inhibition effects of antimicrobial substances (½MIC) combination of strains DS381, DS518, DS620, and DS1515 on spore degradation (SD) and germination (SGI) of toxigenic fungi after 24 h incubation at 30°C in 15% YES medium
Target organism DS381 + DS620 DS381 + DS1515 DS518 + DS620 DS620 + DS1515
SD (%) SGI (%) SD (%) SGI (%) SD (%) SGI (%) SD (%) SGI (%)
A. flavus KACC44986 56.9 100.0 65.6 100.0 - - 40.0 100.0
A. flavus KACC45068 58.1 100.0 48.4 100.0 - - 41.2 100.0
A. flavus KACC45146 -* - 77.5 100.0 - - - -
A. awamori KCCM32316 - - 39.0 100.0 - - - -
A. alutaceus KCCM60421 42.5 100.0 42.5 99.6 - - 28.2 100.0
A. niger ATCC16404 60.1 99.9 - - 68.5 89.9 65.8 100.0
F. fugikuroi KACC46888 75.3 100.0 78.5 100.0 78.1 100.0 - -
F. fugikuroi KACC48352 87.2 100.0 86.2 100.0 85.2 100.0 85.6 97.2
F. proliferatum KACC48354 85.6 100.0 - - 93.1 100.0 91.6 100.0
F. verticillioides KACC48356 30.7 100.0 36.9 100.0 50.5 100.0 36.8 100.0
*-, not tested.
을 가지며(결과 미제시), 항균물질로 작용할 수 있는 protease를 생산하고, DS381과 DS1515 균주는 세포막을 표적으로 하는 계 면활성물질을 생산한다. 이외에도 DS620 균주는 daunomycinone 등의 항진균물질을 생산하기 때문에(Lee and Song, 2018a) 이 러한 물질을 토대로 독소생성 진균의 포자 제거와 발아 저해 를 나타냈을 것으로 추정된다. 이 결과와 유사하게 Lactobacillus plantarum PTCC 1058이 aflatoxin 생성 A. flavus의 포자를 분 해하고 발아를 저해한다고 보고되었으며(Khanafari et al., 2007), Bacillus cereus 균주 배양액, 그리고 Pseudomonas fluorescens 에 의해서 생산된 chitinase를 A. flavus에 처리 시 균사와 포자 의 형태적 손상이 나타날 수 있다고 보고된 바 있다(Akocak et al., 2015).
적 요
본 연구에서는 Paenibacillus elgii DS381, Burkholderia gladioli DS518, Streptomyces lienomycini DS620와 Paenibacillus elgii DS1515 균주에 의해서 생산된 항균물질의 병원성 세균 과 독소생성 진균을 포함한 다양한 미생물에 대한 상호작용 을 조사하였다. 세균 항균물질의 항균활성을 조사하였을 때,
여러 항균물질의 조합은 각각 다른 표적 생물에 대해서 시너 지 효과를 나타내었다[0 < fractional inhibitory concentration index (FICI) < 0.75]. Time kill assay에서 대부분의 항균물질 조합은 세균과 효모의 개체수를 24시간 이내에 10
5cell/ml 이 상 감소시키는 시너지 효과를 나타내었다. 또한 많은 항균물 질 조합이 단독 처리 시보다 여러 독소생성 진균 포자의 분해 를 최대 100%까지 증가시켰으며, 모든 잔류 포자의 발아를 저 해시켰다. 이러한 결과는 잠재적으로 낮은 항균물질 농도로도 항균효과를 나타낼 수 있으므로 유해 미생물의 항균물질에 대 한 내성 발달을 지연시키는데 의미가 있다고 할 수 있다.
References
Aiyegoro OA, Afolayan AJ, and Okoh AI. 2009. Synergistic interaction of Helichrysum pedunculatum leaf extracts with antibiotics against wound infection associated bacteria. Biol. Res. 42, 327–
338.
Akocak PB, Churey JJ, and Worobo RW. 2015. Antagonistic effect of chitinolytic Pseudomonas and Bacillus on growth of fungal hyphae and spores of aflatoxigenic Aspergillus flavus. Food Biosci. 10, 48–58.
Al-Ani I, Zimmermann S, Reichlinga J, and Wink M. 2015. Pharmacological
(A) (B) (C)
(D) (E) (F)
Fig. 4. Effect of antimicrobial substance (½MIC) combination of isolated strains on spore germination of A. flavus KACC45068 [control (A), DS381 + DS620 (B), and DS381 + DS1515 (C)] and F. fugikuroi KACC46888 [control (D), DS381 + DS620 (E) and DS381 + DS1515 (F)].
synergism of bee venom and melittin with antibiotics and plant secondary metabolites against multi drug resistant microbial pathogens. Phytomedicine 22, 245–255.
Aranha C, Gupta S, and Reddy KVR. 2004. Contraceptive efficacy of antimicrobial peptide Nisin: in vitro a nd in vivo studies.
Contraception 69, 333–338.
Fadli M, Saad A, Sayadi S, Chevalier J, Mezrioui NE, Pagès JM, and Hassani L. 2012. Antibacterial activity of Thymus maroccanus and Thymus broussonetii essential oils against nosocomial infection - bacteria and their synergistic potential with antibiotics.
Phytomedicine 19, 464–471.
Hamoud R, Zinmmermann S, Reichling J, and Wink M. 2014.
Synergistic interactions in two-drug and three-drug combinations (thymol, EDTA and vancomycin) against multi drug resistant bacteria including E. coli. Phymedicine 21, 443–447.
Jayaraman P, Sakharkar MK, Lim CS, Tang T, and Sakharkar KR.
2010. Activity and interactions of antibiotic and phytochemical combinations against Pseudomonas aeruginosa in vitro. Int. J.
Biol. Sci. 6, 556–568.
Khanafari A, Soudi H, and Miraboulfathi M. 2007. Biocontrol of Aspergillus flavus and aflatoxin B1 production in corn. J.
Environ. Health. Sci. Eng. 4, 163–168.
Kumar SN, Siji JV, Nambisan B, and Mohandas C. 2012. Activity and synergistic interactions of stilbenes and antibiotic combinations against bacteria in vitro. World J. Microbiol. Biotechnol. 28, 3143–3150.
Lahmar A, Bedoui A, Mokdad-Bzeouich I, Dhaouifi Z, Kalboussi Z, Cheraif I, Ghedira K, and Chekir-Ghedira L. 2017. Reversal of resistance in bacteria underlies synergistic effect of essential oils with conventional antibiotics. Microb. Pathog. 106, 50–59.
Lee DS and Song HG. 2018a. Antibacterial activity of isolated bacteria against Propionibacterium acnes causing acne vulgaris. Korean J. Microbiol. 54, 272–279.
Lee DS and Song HG. 2018b. Antimicrobial activity by Paenibacillus elgii DS381 and its antimicrobial substances against microbial residents on human skin and pathogenic bacteria. Korean J.
Microbiol. 54, 244–253.
Lee DS and Song HG. 2019a. Antifungal activity of bacteria isolated from soil against several toxigenic fungi. Korean J. Microbiol.
55, 377–384.
Lee DS and Song HG. 2019b. Antimicrobial activity of bacterial isolates against microbial residents on human skin and pathogenic bacteria. Korean J. Microbiol. 55, 385–391.
Magalhães L and Nitschke M. 2013. Antimicrobial activity of rhamnolipids against Listeria monocytogenes and their synergistic interaction with nisin. Food Control 29, 138–142.
Naghmouchi K, Drider D, Baah J, and Teather R. 2010. Nisin A and polymyxin B as synergistic inhibitors of Gram-positive and Gram-negative bacteria. Probiotics Antimicrob. Proteins 2, 98.
Najjar MB, Kashtanov D, and Chikindas ML. 2009. Natural antimicrobials ε-Poly-L-lysine and nisin A for control of oral microflora. Probiotics Antimicrob. Proteins 1, 143.
Puño-Sarmiento J, Anderson EM, Park AJ, Khursigara CM, and Foster DEB. 2020. Potentiation of antibiotics by a novel antimicrobial peptide against shiga toxin producing E. coli O157:H7. Sci. Rep.
10, 10029.
Regmi S, Choi YS, Choi YH, Kim YK, Cho SS, Yoo JC, and Suh JW.
2017. Antimicrobial peptide from Bacillus subtilis CSB138:
characterization, killing kinetics, and synergistic potency. Int.
Microbiol. 20, 43–53.
Sopirala MM, Mangino JE, Gebreyes WA, Biller B, Bannerman T, Balada-Llasat JM, and Pancholi P. 2010. Synergy testing by Etest, microdilution checkerboard, and time-kill methods for pan-drug-eesistant Acinetobacter baumannii. Antimicrob. Agents Chemother. 54, 4678–4683.
Veras FF, Correa APF, Welke FE, and Brandelli A. 2016. Inhibition of mycotoxin-producing fungi by Bacillus strains isolated from fish intestines. Int. J. Food Microbiol. 238, 23–32.
White IR and Groot AC. 2006. Cosmetics and skin care products, pp.
493–506. In Frosch PJ, Menne T, and Lepoittevin JP. (eds.), Contact Dermatitis, 4th edn. Springer, Berlin, Germany.