J. Fish Pathol., 22(3) : 383 ~ 389 (2009)
383 383
�Corresponding Author :
The olive flounder Paralichthys olivaceus is one of the most significant fish species in Korea due to human interests in aquaculture and fisheries. The olive flounder stocks currently used in aquaculture have become appreciably different to their wild counterparts and it seems likely that genetics will play an increasingly more important role in achiev- ing further improvements in the performance of the brood stocks. Traits that may be amenable to genet- ic improvement include growth, disease resistance and sex determination. Despite this interest, rela- tively little information is available about olive flounder genes.
Recently, we have conducted expressed sequence
tags (ESTs) analyses of liver, kidney and testis libraries in olive flounder (Lee et al., 2003, 2006, 2007). ESTs are widely used for identification of genes and for studying expression profiles (Adams et al., 1991; Liew et al., 1994). Several of these studies were specifically designed to identify genes involved in innate and acquired immunity. These studies include the EST researches on olive floun- der leucocytes infected with Hirame rhabdovirus (Aoki et al., 1999; Nam et al., 2000), olive flounder leucocytes stimulated with Con A/PMA (Nam et al., 2003) and kidney tissues from peptidoglycan- stimulated olive flounder (Kono et al., 2001). In addition, ESTs have been obtained from kidney and
An EST-based approach for identifying genes expressed in the gills of olive flounder Paralichthys olivaceus
Jeong-Ho Lee, Jae Koo Noh, Hyun Chul Kim, Choul-Ji Park, Byung Hwa Min, Young-Ok Kim
�, Jong-Hyun Kim
��, Kyung-Kil Kim
�, Woo-Jin Kim
�and Jeong-In Myeong
�Genetics and Breeding Research Center, NFRDI, Geoje, 656-842, Korea
�
Biotechnology Research Division, NFRDI, Busan, 619-902, Korea
��
Aquaculture Management Division, NFRDI, Busan, 619-902, Korea
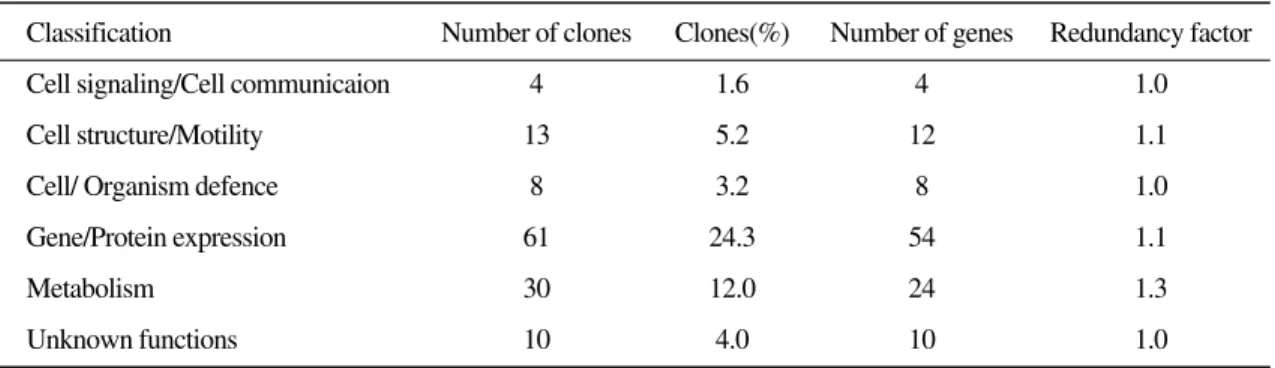
Analysis of expressed sequence tags (ESTs) is an efficient approach for gene discovery, expression pro- filing, and development of resources useful for functional genomics studies. As part of studies on the immune system of olive flounder, a total of 251 EST sequences from gill cDNA library were generated to identify and characterize important genes in the immune machanisms of olive flounder. Of the 251 clones, 126 clones (50.2%) were identified as orthologues of known genes from olive flounder and other organ- isms. Among the 126 EST clones, 16 clones (12.7%) were representing 9 unique genes identified as homol- ogous to the previously reported olive flounder ESTs, 100 clones (79.4%) representing 103unique genes were identified as orthologs of known genes from other organisms. We also identified several kinds of immune associated proteins, indicating EST as a powerful method for identifying immune related genes of fish as well as identifying novel genes. Further studies using cDNA microarrays are needed to identify the differentially expressed transcripts after disease infection.
Key words: Olive flounder, Gill, Expressed sequence tags (ESTs), Expression profile, Immune related
genes
�Corresponding Author : Jeong-In Myeong, Tel : 055-633-1272 Fax : 055-633-0891, E-mail : [email protected]
20...
gill tissues of rainbow trout infected with infectious hematopoietic necrosis virus (IHNV) and from head kidney cells of common carp after stimulation with lipopolysaccharide and concanavalin A (Kono et al., 2001; Savan and Sakai, 2002).
Gene cataloguing and profiling of the gill is an essential part of our effort in EST analysis of immune organs because gills directly contact with the external environment; therefore, the cells cru- cially respond to external biotic and abiotic factors.
For example, gills are the siteof formation of hemo- cyte nodules during foreign particle injection (Mar- tin et al., 2000) and accumulation of viable bacteria during infection (Burgents et al., 2005).
As a part of studies on the immune system of olive flounder, partial cDNA sequences or ESTs were generated to identify and characterize impor- tant genes in the immune machanisms. Here, we report a total of 251 EST sequences from olive flounder gill cDNA library on the purpose of identi- fying genes and their expression profiles in the gill, and improving our understanding of the immune system of olive flounder and the availability as a genomic tool.
Materials and Methods
Tissue preparation and RNA isolation
All experimental fish were raised at Genetics and Breeding Research Center, National Fisheries Research and Development Institute (NFRDI) and maintained in 10 tons flow-through tank at 15 ±1
℃ under a natural photoperiod. Gill samples of 10 randomly selected fish were collected and frozen in liquid nitrogen until isolation of RNA. Pooled gill tissues were ground with a mortar/pestle and then homogenized with a hand-held tissue tearor in RNA extraction buffer followed by the guanidium thio-
cyanate method (Chomczynski and Sacchi, 1987).
Poly (A)+ RNA was purified from total cellular RNA using the PolyA Tract mRNA isolation kit (Promega) according to the manufacturer's instruc- tions.
cDNA library construction
A directional cDNA library of the gills was con- structed using the Uni-ZAP XR cDNA synthesis/Gigapack cloning kit (Stratagene Cloning Systems). cDNA synthesis was carried out using an oligo-(dT)
18primer for the reverse transcription of approximately 5 ㎍ of mRNA and the libraries were constructed by directional cloning based on the man- ufacturer ’ s instruction manual. All primary libraries were amplified and aliquots of each amplified library were stored at both 4 and -70℃. Mass exci- sion was performed and the cDNA inserts from the amplified Uni-ZAP XR libraries were rescued as pBluescript phagemids in SOLR Escherichia coli.
Plasmid preparation and sequencing analysis The plasmid cDNA library was plated to a density appropriate for picking individual colonies. Random clones were grown in 1.5-ml LB medium overnight in 12 × 75 mm culture tubes. Plasmid DNA was prepared by alkaline lysis method (Sambrook et al., 1989) using the Qiagen Spin Column Mini-plasmid kits. Three microlitres of plasmid DNA (about 0.5- 1.0 ㎍) were used in sequencing reactions. Single- pass sequencing of the 5'-termini of selected cDNA clones in phagemid form was performed using the ABI 3100 automatic DNA sequencer (PE Applied Biosystems) and the ABI Prism Big Dye Terminator Cycle Sequencing Ready Reaction kit (PE Applied Biosystems).
Bioinformatic analysis
Bioinformatic analysis was conducted to deter-
mine gene identities using GeneMaster software (Ensoltek). ESTs were assembled in clusters of con- tigous sequences (contig) using ICAtools program (Parsons, 1995). Gene annotation procedures and homology searches of the sequenced ESTs have been locally done by BLASTX for amino acid simi- larity comparisons (Altschul et al., 1997). Matches with the Expect value (E) less than 1.0 × e
-4were considered to be significant. After the BLAST searches, a visual inspection was made to determine if the significant similarity was caused by simple sequences. ESTs with significant similarities in searches were considered orthologs of known genes only when the similarities were not caused by sim- ple sequences. All ESTs that were not identified as orthologs of known genes were designated as unknown EST clones.
Results and Discussion
Gene and orthologue identification
To identify genes expressed in the olive flounder gill and establish their expression profiles, we
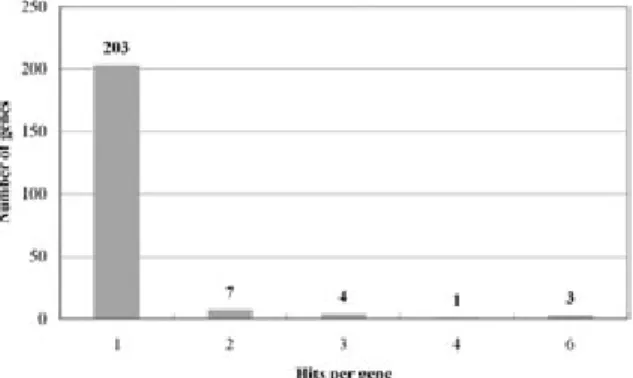
sequenced 251 ESTs from a non-normalized cDNA library of the olive flounder gill. The number of clones sequenced from the cDNA library, the aver- age size of inserts, and the redundancy of the obtained sequences, are given in Table 1. The aver- age insert size was estimated to be 1.6 ± 0.6 kb by PCR amplification of inserts from 20 randomly selected clones. Clustering of the sequences using the assembly program ICAtools software (Parsons, 1995) showed that the 251 gill ESTs were com- posed of 15 clusters and 203 singletons, suggesting that the overall redundancy of the library was 13.1%.
Of the 251clones, 126 (50.2%) were identified as orthologues of known genes from olive flounder and other organisms. The remaining 125 clones (49.8%) could not be identified by similarity com- parisons (E≥1×e
-4). Among the 126 EST clones, 16 clones (12.7%) were representing 9 unique genes identified as homologous to the previously reported olive flounder ESTs, 100 clones (79.4%) representing 103 unique genes were identified as orthologs of known genes from other organisms.
Table 1. General characteristics of olive flounder gill ESTs.
Total cDNA sequenced
a251
Average insert size
b1.6 ± 0.6 kb
Average EST length 621 bp
EST clusters
c15
Singletons
d203
Redundancy
e13.1 %
ESTs with E value<1 ×e
-4(matched) 126 (50.2%)
ESTs with E value ≥1×e
-4(unknown) 125 (49.8%)
a
Length of sequence used for comparison after editing (inserts<100 base pairs were excluded).
b
The average insert size was calculated for 20 randomly selected cDNA clones.
c
ESTs with 90% or greater identity over a 100 bp region were clustered together forming 15 EST clusters.
d
203 sequences did not sufficiently match any sequence in the data set to allow assembly.
e