Article
http://dx.doi.org/10.4217/OPR.2013.35.2.157
남해동부해역의 표층 수괴 변화에 따른 환경요인과 식물플랑크톤 군집의 계절적 변화
장풍국·현봉길·차형곤·정한식·장민철·신경순
* 한국해양과학기술원 남해연구소(656-830) 경상남도 거제시 장목면 장목1길 41
Seasonal Variation of Phytoplankton Assemblages Related to Surface Water Mass in the Eastern Part of the South Sea in Korea
Pung-Guk Jang, Bonggil Hyun, Hyung-Gon Cha, Han-Sik Chung, Min-Chul Jang, and Kyoungsoon Shin
*South Sea Research Institute, KIOST Geoje 656-830, Korea
Abstract : We investigated the seasonal succession of phytoplankton assemblages in the eastern part of the South Sea of Korea in relation to surface water masses. The study areas are under the direct influence of the Tsushima Warm Current (TCW) throughout the whole year, with its strength known to be seasonally variable. The region is also influenced by coastal waters (CW) driven from the South Sea of Korea and East China Sea, particularly in summer, as indicated by low salinity in the surface water. Nutrient property of the TCW can reveals whether the origin of the TCW is the nutrient-rich Kuroshio Current or the oligotropic Taiwan Warm Current. Surface chlorophyll-a (Chl-a) concentrations displayed a large seasonal variation for all stations, with high values found in spring and autumn and low values in summer and winter. At station M (offshore) and P (intermediate location between M and R), Chl-a concentrations in October were higher than those in March, when spring bloom normally occurs. This may be related to deeper mixed layer depths in October. Diatoms dominated under conditions of high nutrient supply in which Chaetoceros spp. and Skeletonema costatum-like spp. were abundant. S. costatum-like spp. dominated at stations R (onshore station) and P in December when there was greater nutrient supply, especially of phosphate. Flagellates and dinoflagellates dominated at all three stations after diatoms blooms. Dominant species were Scrippsiella trochoid in April and Ceratium furca in October at station R, and Gyrodinium spp. and Gymnodinium spp.
at station M during summer, when the effect of the oligotropic Taiwan Warm Current and the oligotropic coastal water from East China Sea were strong. Redundancy analysis showed clear seasonal successions in the phytoplankton community and environmental conditions, in which both principal components 1 and 2 accounted for 69.6% of total variance. Our results suggested that environmental conditions seemed to be determined by the origin of the TCW and the relative seasonal strength of the water masses of the TCW and CW, which may affect phytoplankton growth and compositions in the study area.
Key words : nutrient, Phytoplankton assemblage, Tsushima Warm Current, Korea Strait
*Corresponding author. E-mail : [email protected]
1. 서 론
온대 기후 지역에서 식물플랑크톤 군집의 계절적 변화 를 조절하는 주요 인자는 수온, 빛, 영양염, 동물플랑크톤 에 의한 섭식 및 수괴의 안정도 등이다. 특히 이중에서도 영양염 농도와 구성비가 중요한 역할을 한다고 보고되고 있다(De Baar 1994; Officer and Rather 1980; Tilman 1982). 영양염과 식물플랑크톤 관계에 관련된 최초 이론은 19세기 Carl Sprengel과 Justus von Liebig가 제안한 Liebig's 최소량의 법칙이며, 이와 관련된 연구들은 식물플 랑크톤의 성장이 하나의 영양염에 의해 제한되며, 담수에 서는 인산염, 해양에서는 질산염이 주요 제한 인자로 작용 한다고 보고하였다(De Baar 1994). 또한 Office and Ryther (1980)는 영양염 제한이 식물플랑크톤의 일차생산력뿐만 아니라 군집구조에도 영향을 미친다고 보고하였다. Tilman (1982)은 영양염의 절대적인 농도보다도 영양염의 구성비 에 따라서 식물플랑크톤 종조성에 영향을 미친다고 보고 하였다. 최근에 연구는 한 가지 이상의 영양염 제한이 식 물플랑크톤 일차생산력 및 종조성에 영향을 줄 수 있고, 또한 식물플랑크톤 종에 따른 영양염 비에 대한 특이성이 보고되었다(Arrigo 2004).
우리나라 남해동부해역에 위치한 대한해협은 대마도를 중심으로 서수도와 동수도로 나누어지며, 연구 지역인 서 수도는 중앙에 위치한 수심 200 m의 해곡을 제외하면 수 심 130 m 미만의 평탄한 지형으로 이루어진 좁은 대륙붕 지역이다. 대한해협을 연중 통과하는 대마난류는 동중국 해에서 중국연안수와 황해 냉수 등과 같은 수괴와 혼합되 어 제주도 인근 해역을 걸쳐 대한해협을 통과 한 후 동한 난류를 형성하며 동해로 유입되어 우리나라 남해와 동해 에 열과 염, 그리고 영양염 등 다양한 물질을 수송함과 더 불어 해수 순환을 조절하는 중요한 역할을 한다(Yanagi 2002; Onitsuka et al. 2007; Morimoto et al. 2009). 대한 해협 서수로에 유입되는 표층 수괴는 대마난류의 기원에 따라서 다른 특성을 나타낸다. 쿠로시오에서 분주된 수괴 가 우세한 시기에는 고 염분수의 영양염이 풍부한 특성을 보이고, 타이완 난류에서 기원하는 수괴가 우세한 시기에 는 저 염수의 빈영양염의 특성을 보인다(Jang et al.
2013). 대한해협을 포함한 남해동부해역은 대마난류 이외 에도 남해 연안수와 장강 희석수 등의 다양한 수괴가 존 재하고, 계절에 따라 그들의 확장 범위가 달라 이 해역의 수괴의 계절적 변동성이 크다(승 1992; 조 등 1994). 양 등 (1998)은 우리나라 남해안에 형성되는 수온 전선이 겨 울철에는 북서계절풍과 해표면의 복사열 등의 영향으로 인해 연안수가 남해동부해역의 외해로 확장되는 반면 여 름철에는 대마난류의 세기가 강해지면서 수온 전선이 연 안쪽으로 이동하는 등 대마난류의 세기와 남해연안수의
확장에 따라서 수온전선의 계절적 변동이 발생한다고 보 고하였다.
결과적으로 남해동부해역은 계절마다 다른 특성의 수괴 들이 나타남으로써 해양생태계에 물리·화학적인 요인들 의 계절적 변동에 요인이 될 수 있으며, 이로 인해 식물플 랑크톤 개체수 및 군집구조의 계절적 변화가 발생할 가능 성이 있다. Jang et al. (2013)은 대한해협에서 대마난류의 계절적 세기의 변화가 물리·화학적인 요인 및 식물플랑 크톤 군집구조의 계절적 변동성을 조절한다고 보고하였지 만, 이 결과는 대한해협 서수로 중앙에 위치한 외해의 한 정점에서만의 조사결과로서 외해수 유입에 따른 남해안 연안을 포함하는 남해동부해역의 식물플랑크톤 군집구조 에 미치는 영향을 이해하기에는 아직 미흡하다. 또한 그 외 대한해협 인근에서 수행된 조사들은 이 해역을 통과하 는 수괴의 계절적 변동 및 연변화에 대한 물리적인 조사 가 주를 이루고 있으며(Teague et al. 2003; Yun et al.
2004; Li et al. 2006), 식물플랑크톤에 대한 연구(Park and Park 1997; 정 등 1998)와 영양염을 포함한 화학적인 인자 들에 대한 연구(이 1999; 김과 이 2004)는 주로 특정한 시 기(여름)에 간헐적으로 이루어져 계절적 수괴 변동에 따른 부유생태계 변화를 정확히 이해하기에는 아직 어려움이 있다.
향후 기후변화에 따라 외해수의 세기가 달라지면 남해 안 표층 수괴의 특성이 달라질 것으로 예측되며, 이는 남 해안 연안뿐만 아니라 외해의 부유생태계에도 영향을 미 칠 것이다. 본 연구의 목적은 남해동부해역에 영향을 미치 는 외해수의 세력변화가 남해연안 및 외해의 물리·화학 적인 요인 및 식물플랑크톤의 계절적 분포와 연관성이 있 는지를 파악하고자 한다.
2. 재료 및 방법
연구 지역의 조사정점은 연안수, 외해수의 영향 등을 고려하여 연안지역에 인접한 정점 R(128o 54.108E, 34o 55.007N), 대한해협 서수로에 위치한 정점 M(129o 05.903E, 34o 47.419N), 그리고 부산 외해의 정점 P(129o 11.817E, 34o 01.103N)을 선정하여 2012년 1월부터 12월까지 해양 조사선 장목호를 이용하여 월별 환경인자 및 식물플랑크 톤 군집구조의 변화를 조사하였다(Fig. 1). 수온과 염분은 장목호에 장착된 SBE 911plus CTD(Sea-Bird Electronics) 를 이용하여 관측되었으며, 영양염과 엽록소-a 그리고 식 물플랑크톤 분석을 위한 해수시료는 로젯멀티 샘플러 (Rosette Multi-sampler)에 부착된 Niskin 채수기를 이용하 여 채수되었다. 영양염 분석시료 해수는 유리 섬유 여과지 (GF/F filter, 직경 47 mm, Whatman, pore size 0.7 µm)로 여과하고, 100 ml 정도만을 scintillation vials에 넣어 분석
전까지 냉동 보관하였다. 영양염(질산염+아질산염(질산 염), 인산염, 규산염)은 Flow Injection Analyzer(QuAAtro 8000, SEAL Analytical Co.)를 이용해 분석하였다. 해수 표준물질(CSK standard solution, Wako Pure Chemical industries, Osaka, Japan)을 이용하여 보정하였으며, 각 영 양염에 대한 정밀도는 5% 이내의 값을 유지하였다.
엽록소-a 분석을 위한 시료(여과지)는 일정양의 채수된 해수(0.5-1 L)를 유리 섬유 여과지(GF/F filter, 직경 47 mm, Whatman, pore size 0.7µm)에 여과하여, 그 여과지를 15 ml 원심분리용 tube에 넣어 분석 전까지 냉동 보관하였 고, 분석 시 90% 아세톤(Acetone) 10 ml을 넣은 후 교반 시킨 다음 냉암소에서 overnight 시키고 24시간 이내에 측 정하였다. 추출된 색소 중에 섞여 있는 입자를 제거하기 위해 1000 g에서 5분 동안 원심분리시킨 후 상등액만을 취하여 형광광도계(Turner Designs 10-AU Fluorometer) 로 측정하였다(Parsons et al. 1984).
식물플랑크톤 분석을 위한 해수시료 약 500 ml는 전체 부피의 약 1% 정도 루골용액(Lugol solution)을 첨가하여 고정되었고, 고정된 해수는 하루 이상 침전 시킨 후 상등 액을 걷어내고 약 50 ml로 농축시켜 일정한 양을 취하여 Sedwick-Rafter counting chamber에 옮긴 후 광학현미경 으로 식물플랑크톤을 계수 및 동정하였다(Sournia 1978).
식물플랑크톤의 종동정은 Cupp (1943), Yamaji (1979), Dodge (1982), Rines and Hargraves (1988) 등을 참조하
여 규조류와 와편모류 중심으로 이루어졌으며, 은편모류 와 유글레나류 및 20 µm 이하의 작은 크기의 편모류는 광 학현미경 상으로 종을 구분할 수 없어 그룹 수준으로만 개수되었다.
수괴의 특성 변화에 따른 환경요인(수온, 염분, 영양염, 및 엽록소-a)과 식물플랑크톤 그룹 간의 변화를 이해하기 위해서 이들 인자를 사용해 Primer 6.0을 이용한 집괴분석 (cluster analysis) 및 Canoco 4.5 software를 이용한 중복 분석(redundancy analysis, RDA)을 수행하였다.
3. 연구 결과
표층 수온과 염분의 월별 분포는 TS diagram(Temperature -salinity diagram)으로 도식화하였으며(Fig. 2), 표층 영양 염 및 엽록소-a의 월별 변화는 Fig. 3에 나타내었다. 표층 식물플랑크톤 그룹과 한번이라도 20% 이상 우점율을 보 인 식물플랑크톤 종들의 우점율은 Fig. 4과 5에 나타내었 다. 정점 R에서는 1, 2, 3월에 수온은 10oC 이상, 염분은 34‰ 이상을 나타내었다. 4월 이후부터는 염분이 34‰ 이 하를 나타내었으며, 9월에 31.5‰로 최소치를 보였다. 정 점 R의 영양염 월별변화는 비록 4월에 영양염의 갑작스런 증가가 관측되었지만 1월부터 감소하기 시작하였으며 9월 이후에 다시 증가하는 경향을 보여, 겨울철에 높은 농도를 나타내고 그 이후에 감소하다가 가을철이 되면서 다시 증 가하는 계절적 경향을 보였다. Dortch et al. (1992)은 영 양염 제한을 영양염 농도와 영양염 비의 조합으로서 언급 하였다. 이 논문은 식물플랑크톤 성장에 제한을 주는 영양 염 임계농도(threshold)를 각각 질산염 1 µM, 인산염 0.2µM, 규산염 2 µM 이하로 규정하였다. 또한 영양염 농도가 임계농도보다 높은 상태에서 NP 비가 30 이상, SP 비가 3 이상이면 잠재적인 인산염 제한(P potential limitation), NP 비가 10 이하, SN 비가 1 이상이면 잠재 적인 질산염 제한(N potential limitation), SN 비가 1 이하 SP 비가 3 이하면 잠재적인 규산염 제한으로 규정했다(Si potential limitation). 정점 R에서 영양염 제한을 보면 3월 에 모든 영양염이 공동으로 제한되었으며, 4, 5월과 9, 10, 11월은 인산염 제한, 그리고 6, 7, 8월은 질산염과 인산염 이 공동으로 제한되었다(Fig. 6). 엽록소-a의 월별 농도 분 포를 보면, 2월부터 증가하기 시작하여 3월에 가장 높은 값을 보였으며 1, 2, 7, 8월을 제외한 시기에 1 µg L−1 이 상의 높은 값을 보였다. 계절별로 보면, 봄철(3-5월: 1.83 µg L−1)과 가을철(9-11월: 1.72 µg L−1)이 여름철(6-8월:
0.94µg L−1) 및 겨울철(12-2월: 0.81 µg L−1) 보다 높았다.
정점 R에서 출현한 식물플랑크톤 그룹을 보면 규조류가 가장 우점(평균 점유율 72%)하여 나타났으며, 와편모류는 5월과 10월에 평균 52%의 높은 점유율을 보였다. 특히 규 Fig. 1. Map of the study area showing locations of
sampling station in the western channel of Korea Strait
조류 중 주로 Chaetoceros spp.가 가장 우점(평균 점유율 39.7%)하여 출현하였으며, 12월과 1월에 Skeletonema costatum-like spp.가 각각 28%와 20%의 높은 점유율을 보 였다. 와편모류가 우점한 5월과 10월에는 주로 Scrippsiella trochoidea와 Ceratium spp.가 각각 우점하여 출현하였다.
정점 P의 TS diagram 분포를 보면, 겨울철과 봄철에 수 온이 10oC 이상, 염분 34‰ 이상을 나타내었고 그 외 시 기에는 34‰ 이하의 염분 농도를 보였다. 가장 낮은 염분 은 9월에 31.3‰을 보였다. 영양염 농도의 월별 및 계절별 변화는 정점 R과 유사한 경향을 보였으며, 다른 정점에 비해 12월에 인산염 농도가 높게 나타났다. 인산염은 4, 9, 10, 11월에 제한인자로 나타났으며, 3, 5, 6, 7, 8월은 질산염과 인산염 모두가 제한인자로 나타났다. 엽록소-a 의 월별 변화는 2월부터 증가하고 4월부터 감소하였다가 8월부터 다시 증가하여 10월에 가장 높은 2.59 µg L−1의 농도를 보인 후 다시 감소하는 양상을 보였다. 엽록소-a의 계절적 변화 경향은 가을철(평균 1.77 µg L−1)에 봄철 및 겨울철(평균 0.98, 0.85 µg L−1)보다 높았으며, 여름철(평 균 0.26 µg L−1)은 최저값을 보였다. 식물플랑크톤 그룹 중 규조류는 연평균 점유율 69%로 항상 우점하여 출현하 였으며, 규조류의 점유율이 낮아진 4월에는 은편모류의 점유율이 35%로 증가하였고, 5월에는 20 µm 이하의 편모 류가 42%, 11월에는 이 두 그룹이 각각 30% 이상의 점유 율을 나타내었다. 규조류 우점 시기에는 주로 Chaetoceros
spp.(규조류 중 57%)가 출현하였지만, 12월에는 규조류 중에서 S. costatum-like spp.가 점유율 51.7%로 우점하여 나타났다. 와편모류는 연평균 10% 이하로 낮은 점유율을 보였으며, 전체 식물플랑크톤 중에서 20% 이상 우점한 와 편모류 종은 관찰되지 않았다.
정점 M의 TS diagram을 보면 겨울철에서 봄철까지 수 온이 10oC 이상의 값을 보였으며, 염분도 34‰ 이상을 나 타내어 정점 P와 유사한 경향을 보였다. 여름철부터 가을 철까지 34‰ 이하의 염분을 나타냈었으며, 7월에는 32.0‰
로 최소치를 보였다. 영양염은 3월 이후에 급격히 감소하 다가 9월부터 증가하기 시작, 봄철부터는 낮은 값을 보이 다가 가을철부터 다시 증가하는 경향을 보였다. 특히 인산 염과 질산염 모두 겨울철을 제외한 대부분 시기에 제한인 자로 나타났다. 엽록소-a의 월별 분포는 2월부터 증가하기 시작하여 3월 이후에는 낮은 값을 보이다가 9월과 10월에 증가하고 그 이후 다시 감소하는 경향을 보였다. 계절별 변동양상은 가을(평균 1.35 µg L−1)이 봄(평균 0.64 µg L−1) 또는 겨울(평균 0.78 µg L−1)보다 높았으며, 여름(평균 0.18µg L−1)에는 최소치를 보였다. 식물플랑크톤 그룹 중 에서 규조류는 1월부터 4월까지 그리고 9, 10, 12월에 평 균 83%의 높은 점유율을 보이지만 그 외 시기에는 와편 모류를 포함한 편모류의 점유율이 증가하는 경향을 보인 다. 규조류 우점시기에는 Chaetoceros spp.가 가장 우점 (규조류 중 57%)하여 출현하고 12월에 다른 정점과는 달 Fig. 2. Temperature and salinity diagram for surface waters at the study areas in 2012
리 Chaetoceros spp.의 점유율이 감소할 때 Lauderia spp.
와 Nitzschia spp.의 점유율이 증가하였다. 와편모류 점유 율은 5월부터 증가하기 시작해서 8월까지 높은 점유율을 보였으며, 주로 출현한 종은 Gyrodinium spp.(와편모류 중
28.3%)와 Gymnodium spp.(와편모류 중 42.5%)으로 나타 났다.
조사기간 동안에 우점한 식물플랑크톤 그룹의 개체수와 환경요인과의 상관성을 분석하기 위해서 이들 인자를 이 Fig. 3. Monthly variation of nitrate, phosphate, silicate and chlorophyll-a in the surface water of stations R, P, and M
during 2012
용해 중복분석과 집괴분석을 실시하였다(Fig. 7). 중복분 석 결과 수온과 규산염이 식물플랑크톤 그룹의 변동에 유 의(p < 0.05)한 영향을 보였다(Table 1). 제 1, 2축에서 환 경요인 수온, 염분, 영양염, 엽록소-a)이 출현한 식물플랑 크톤 그룹 변화의 69.6%을 설명하였다. 환경요인 중에서 수온, 규산염, 엽록소-a가 제 1축과 양(+)의 상관성을 보였 으며, 제 2축과는 수온 만이 양(+)의 상관성을 보였다. 수
층혼합이 강하게 일어나는 겨울철 조사 시기는 영양염 인 자와 유사한 방향에 위치하였고, 성층 형성이 강하게 일어 나는 여름철 조사 시기는 영양염 인자와 반대되는 방향에 위치하였다. 조사시기 3월에 엽록소-a 농도가 높았음에도 불구하고 겨울철 조사 시점보다 엽록소-a 요인의 오른쪽 에 위치하고 있으며, 3월에 비해 상대적으로 영양염 농도 가 높고 엽록소-a 농도가 높았던 가을철은 왼쪽에 위치하 Fig. 4. Kite diagram of dominant phytoplankton groups at the surface water of stations R, P, and M in 2012
고 하고 있다. 엽록소-a와 규조류도 영양염과 유사한 위치 에 있어 이 들간의 상관성이 있음을 보여준다. 와편모류는 수온이 높아지는 시기인 여름철에 많이 나타나 수온과 유 사한 위치에 있다. 20 µm 이하의 편모류 종(nanoflagellate)
은 봄철과 여름철에 개체수가 증가하는 경향을 보여 수온 과 염분 사이에 위치하였다. 결과적으로 중복분석 결과 환 경 인자와 식물플랑크톤 그룹간에 시계 반대 방향의 뚜렷 한 계절성을 나타내었다. 환경인자와 식물플랑크톤 그룹 Fig. 5. Kite diagram of dominant phytoplankton species at the surface water of stations R, P, and M in 2012
간의 집괴분석 결과를 보면, 모든 정점에서 1, 2, 11월, 정 점 R과 P는 3, 8, 12월, 그리고 정점 M과 P는 5, 6, 10월 에 95% 이상의 유사성을 보였다.
4. 토 의
대마난류 기원에 대해서는 아직까지 명확하게 규명되고 있지 않지만, 최근 연구에 따르면 타이완에서 유입되는 수 괴와 쿠로시오 해류(Kuroshio Current)에서 분주된 수괴의 계절적 변동이 대한해협으로 유입되는 대마난류의 특성을 결정하는 것으로 보고하고 있다(Isobe 1999; Guo et al.
2006). Isobe (1999)는 타이완에서 유입되는 수괴가 대마 난류의 일차적인 수괴이고 가을철에는 쿠로시오 해류에서 분주된 수괴가 대마난류의 66% 정도 차지한다고 보고하 였으며, Guo et al. (2006)는 여름철에는 타이완에서 유입 되는 수괴와 쿠로시오 해류에서 분주된 수괴가 절반씩 영 향을 미치는 반면, 겨울철에는 쿠로시오 해류에서 분주된 수괴가 80% 이상을 차지한다고 보고하고 있다. 즉, 쿠로 시오 해류 기원의 대마난류는 가을철부터 겨울철까지 영 향을 미치고, 타이완 난류 기원의 대마난류는 봄철부터 여 름철까지 영향을 미치는 것으로 판단된다. 쿠로시오 해류 는 동중국해로 유입되면서 지형적인 특성으로 인해 수층 이 혼합되어 영양염이 풍부한 수괴가 되지만, 타이완 난류 는 빈영양수괴인 것으로 알려져 있다(Gong et al. 1996;
Jang et al. 2013). 결과적으로 조사지역에 유입되는 외해 수의 역학적 변동이 영양염 농도의 계절적 변화에 상당한 영향을 주고 있다.
겨울철 수층 혼합은 조사지역의 표층수에 풍부한 영양 염을 공급하지만, 빛의 조건이 좋은 상층부에 존재하는 식 물플랑크톤을 유광층 아래로 이동시킴으로써 식물플랑크 톤 성장에 필수적인 광조건이 제한요인으로 작용하여 엽 록소-a 농도가 낮게 나타나는 것으로 여겨진다(Bouman et al. 2010). 12월에 정점 M에서 정점 R과 P보다 상대적 으로 낮은 영양염 농도를 보이는데, 이는 정점 R의 수심 이 앝아서 저층에 축적된 고농도의 영양염이 보다 더 수 월하게 공급되고, 정점 P는 동해로부터 유입되는 고영양 염의 저층냉수의 영향으로 정점 M보다 높은 영양염 농도 를 보이는 것으로 여겨진다. 대한해협 저층냉수는 여름과 겨울에 동해로부터 대한해협 서수로로 유입되며(민 등 2011), 정점 P는 동해로 연결되는 입구에 위치하고 있기 때문에 정점 M보다 이러한 저층냉수의 영향을 더 받을 수 있는 지역으로 판단된다. 저층냉수는 대마난류 아래에 존재하고 대마난류보다 영양염 농도가 높으며, 특히 인산 염 농도가 높은 것으로 보고되고 있다(김과 이 2004; Jang et al. 2013). 식물플랑크톤 그룹 중 규조류가 모든 정점에서 가장 우점하여 나타났다. 규조류 우점시기에는 Chaetoceros Fig. 6. Scatter diagrams of atomic nutrient ratios for the
surface water at stations R, P, and M in 2012.
Nutrient limitation defines threshold concentration (nitrate <1µM, phosphate <0.2 µM, and silicate
<2µM): red circle means N P Si co-limitation, yellow circle means N and P co-limitation, and green circle means P limitation. P potential limitation defines NP ratio >30 and SP ratio >3, and N potential limitation defines NP ratio <10 and SN ratio <1 (Dortch and Whitledge 1992)
Fig. 7. Relationship between environmental factors and abundance of phytoplankton groups obtained with the redundancy analysis (RDA) at stations R, P, and M in 2012. The grouping of month by each station (dotted circles) was based on similarity >95% obtained from cluster analysis on correlation matrix of log transformed data
Table 1. Summary of redundancy analysis (RDA) for abundance of phytoplankton groups and environmental factors at stations R, M and P in the western channel of Korea Strait during 2012
Axis-1 Axis-2 Axis-3 Axis-4
Eigenvalues 0.155 0.080 0.056 0.034
Groups-environment correlations 0.680 0.633 0.533 0.660 Cumulative percentage variance
of groups data 15.5 23.5 29.2 32.6
of groups-environment relation 46.0 69.6 86.2 96.3 Sum of all eigenvalues 1.000
Sum of all canonical eigenvalues 0.338
Environmental factors λ F-values P-values Axis-1 Axis-2
Temperature 0.11 4.19 0.002 -0.477 0.397
Silicate 0.08 3.41 0.008 -0.135 -0.508
Chlorophyll-a 0.06 2.24 0.054 -0.097 -0.362
Nitrate 0.05 2.18 0.070 0.142 -0.505
Phosphate 0.02 1.22 0.296 0.165 -0.398
Salinity 0.02 0.67 0.622 0.495 -0.090
spp.가 모든 정점에서 가장 우점하여 출현하였지만, 12월 에는 정점마다 다소 다른 경향을 보였다. 이 시기에 정점 R과 P는 S. costatum-like spp.의 점유율이 증가하였으며, 특히 정점 P에서는 점유율이 전체 식물플랑크톤 중 58%
로 Chaetoceros spp.보다 더 우점하여 출현하였다. 이러한 규조류의 시공간적 우점 출현종 차이는 영양염 구성비에 따른 생존경쟁의 차이에 의한 것으로 추정할 수 있다. 인 공생태계 실험을 통한 선행 연구에 따르면 S. costatum- like spp.은 영양염 농도가 높고 NP 비가 낮은 시기에 경 쟁력이 있는 반면, Chaetoceros spp.는 영양염 농도가 낮고 NP 비가 높은 시기에 경쟁에서 앞선다고 보고하였 다(Lagus et al. 2004). 특히 인산염 농도가 높은 경우에 식물플랑크톤의 빠른 성장과 관련된 ATP(Adenosine triphosphate)을 많이 가지고 있는 S. costatum-like spp.가 다른 규조류에 비해 경쟁에서 앞서며(Arrigo 2004), 남해 안 연안에 위치한 장목만에서도 이 두 종의 경쟁이 인산 염 농도에 의해 조절된다고 보고한 바 있다(장 등 2010).
이와 같은 연구결과는 12월 영양염 농도가 높고, 특히 인 산염 농도가 높게 나타난 정점 R과 P의 결과와 일치한다.
이 시기에 정점 M은 S. costatum-like spp.보다 Lauderia spp.(대부분 Lauderia delicatula)와 Nitzschia spp.가 우점하 였다. Sommer (1994)는 광조건에 대한 식물플랑크톤 실 험에서 pennate 규조류인 Lauderia borealis와 Nitzschia pungens는 centric 규조류인 Stephanopyxis palmeriana보 다 낮은 광조건에서 더 경쟁력이 있음을 보고하였다. 정점 M은 수심이 깊어 수층혼합이 일어나는 시기에 빛의 제한 을 받을 수 있으며 정점 R과 P에 비해 영양염 농도도 낮 다. 결과적으로 정점 M은 상대적으로 낮은 인산염 농도 및 광조건 때문에 Lauderia spp.와 Nitzschia spp.가 centric 규조류인 S. costatum-like spp. 보다 우점한 것으로 판단된다.
연구지역의 모든 정점에서 3월에 영양염 농도가 낮아지 고 엽록소-a 농도가 증가하지만, 정점 M은 다른 두 정점 에 비해 영양염 농도가 높은 경향을 보였다. 정점 R과 P 는 연안수의 영향으로 4월에 영양염 농도가 증가하고 엽 록소-a 농도도 높게 나타났으며, 정점 R은 5월까지 질산 염 농도가 1.55 µM로 질산염 임계농도(1 µM 이하) 보다 높은 값을 보여 엽록소-a 농도가 높게 유지된 것으로 보인 다. 정점 M은 4월부터 영양염 농도가 낮고 엽록소-a 농도 도 낮아져 봄철 외해수의 세력이 약해지면서 연안수의 영 향이 외해까지 확장되지만 정점 M까지는 영향을 미치지 못하는 것으로 파악된다. 4월 정점 R과 P의 수온이 정점 M보다 약 1.5oC 낮은 것도 이러한 특성을 반영하고 있는 것으로 보여진다. 봄철 식물플랑크톤 그룹은 모든 정점 에서 3월까지 규조류가 우점하지만, 그 이후 영양염 농 도가 제한되면서 규조류 점유율이 감소하고 와편모류를
포함한 편모류 점유율이 모든 정점에서 증가하였으며, 이러한 경향은 5월에 더욱 뚜렷해진다. 정점 R은 5월에 와편모류의 기여도가 54%로 증가하고, 정점 P와 M은 각각 nanoflagellate가 42%, 와편모류를 포함한 편모류가 80% 정도의 높은 점유율을 보인다. 정점 R은 와편모류 중 Scrippsiella trochoide가 우점하여지만, 정점 M은 Gyrodinium spp.와 Gymnodinium spp.가 우점하는 특성을 보였다. S. trochoide는 봄철과 여름철 연안에서 대발생을 일을 킬 수 있는 종으로 알려져 있으며(임 등 1993; Song et al. 2004), 연안 지역에서의 이 종의 대발생 및 사멸은 영양염 변화와는 직접적인 관계가 없고 바람과 같은 물리 적인 요인에 의해서 발생한다고 보고되고 있다(Yin et al.
2008). 결과적으로 정점 R에서 영양염 농도가 낮음에도 불구하고 이 종이 우점한 이유는 연안 가까운 정점 R의 식물플랑크톤 분포가 바람이나 해류와 같은 물리적인 요 인에 의해 연안의 영향을 받았기 때문인 것으로 추측된 다. 특히 봄철은 대마난류수의 세기가 약해지기 때문에 겨 울철보다 연안수가 더 쉽게 외해에 영향을 미칠 수 있는 여건이 된다. Gyrodinium spp.와 Gymnodinium spp.는 무 각의 혼합영양을 하는 와편모류로 정점 M에서 여름철에 도 주로 우점하는 종이며, 다른 종에 비해 빈영양상태에서 경쟁력을 가지는 것으로 보고되고 있다(Jang et al. 2013).
정점 M은 빈영양의 대마난류의 영향으로 4월부터 지속적 으로 질산염과 인산염이 공동제한요인(co-limitation)으로 나타나며, 이로 인해 이 종들이 여름철까지 우점하는 것으 로 보인다. 정점 P는 3월과 5월에 질산염과 인산염이 성 장제한요인으로 나타나지만, 4월에는 영양염이 증가하는 경향도 보여 정점 M 보다는 영양염 조건이 좋은 것으로 판단된다. 이러한 영양염 조건은 작은 크기의 편모를 가진 식물플랑크톤(unidentified phytoplankton, nanoflagellate) 이 우점하기에 유리한 조건이다. 이 종들은 편모를 가지고 있어 운동성이 규조류보다 뛰어나고 크기가 작아 단위면 적당 표면적이 넓어 규조류보다는 영양염을 더 효율적으 로 흡수할 수 있기에 낮은 영양염 조건하에서도 잘 적응 하는 종으로 알려져 있다(Raven 1998; Finkel et al. 2004).
결과적으로 봄철에는 각각의 정점마다 수괴의 영양염 특 성에 따라 서로 다른 식물플랑크톤 그룹 및 종이 우점한 것으로 판단된다. 중복분석을 보면 겨울에서 봄철로 전이 되는 시점인 3월에는 정점 R과 P가 95%의 유사성을 보이 는 반면, 봄철에서 여름철로 전이되는 시점인 5월에는 정 점 P와 M이 95%의 유사성을 보인다. 이는 봄철 외해수 세기의 차이에 따라서 조사 지역의 부유생태계 차이를 나 타낼 수 있음을 보여준다.
가장 낮은 염분을 나타낸 시기는 정점 R과 P는 9월인 반면 정점 M은 7월에 나타났다(Fig. 2). 여름철에 정점 M 은 연구 지역 중에서 가장 외해에 위치하고 있음에도 불
구하고 정점 R과 P보다 낮은 염분 특성을 보였으며, 7월 과 8월에는 수온 또한 상대적으로 높은 값을 보였다. 이는 정점 M이 여름철에 동중국해에서 중국연안수인 장강희석 수가 타이완난류 기원 대마난류와 혼합되어 유입되기 때 문이다. 정점 M으로부터 장강은 남서쪽으로 1100 km 떨 어져 있어서 장강희석수가 방류된 후, 물론 바람의 방향과 장강의 방류량에 따라 약간의 시간차이는 있겠지만, 대략 한 달 정도의 시간 후에 대한해협으로 유입되는 것으로 보고되고 있다(Chang and Isobe 2003; Kim et al. 2009;
Jang et al. 2013). 이 시기에 빈영양 특성의 타이완 난류와 연안을 따라 흘러오며 식물플랑크톤에 의해 영양염이 고 갈된 장강희석수가 혼합되어 유입되기 때문에 이 시기에 대한해협으로 유입되는 표층 수괴는 영양염이 고갈되어 있으며, 이로 인해 엽록소-a 농도도 낮다. 또한 여름철에 대마난류의 세기가 강해져 남해안에서 형성되는 대마난류 수와 연안수간 수온 전선은 연안쪽으로 이동한다(양 등 1998). 이로 인해 정점 P도 빈영양 특성의 대마난류의 영 향을 받을 것으로 추정되며, 따라서 이 시기에 정점 M과 P에 영양염 농도도 낮고 엽록소-a 농도도 낮은 것으로 판 단된다. 하지만 식물플랑크톤 그룹의 우점율 변화를 보면 정점 M은 와편모류가 우점하고, 정점 P는 규조류의 점유 율이 정점 R 보다는 낮지만 정점 M 보다는 월등히 높다.
그리고 집괴분석에서도 8월 정점 R과 P는 95%의 유사성 을 나타낸다. 결과적으로 정점 P가 여름철에 외해수의 영 향을 지속적으로 받지 않는다는 것을 의미한다. 정점 R에 서 영양염 농도가 낮은데도 불구하고 엽록소-a 농도가 다 른 두 정점보다 높은 이유는 아마도 여름철 수온 전선이 정점 R 근처에서 형성되어 식물플랑크톤이 집적되기 때 문으로 판단된다. 여름철 외해수와 연안수 사이에 형성되 는 수온 전선은 수평적인 수층안정도 증가와 더불어 수직
적으로도 성층이 형성되면서 상층부 수괴의 수층안정도가 증가하게 된다. 이는 수온 전선이 형성되는 연안 안쪽에 식물플랑크톤 집적이 일어날 수 있는 조건이다. 실제 정점 R의 상층부 수괴(5-20 m)는 표층보다도 더 높은 엽록소-a 농도(평균 1.40 µg L−1)가 관측되었다.
우리나라는 여름철에 자주 태풍이 발생하고 9월에 발생 하는 태풍의 강도가 큰 경우가 많다. 태풍은 강우와 강한 바람을 동반하기 때문에 수층혼합을 유발하여 저층의 고 영양염을 표층으로 공급하고, 태풍이 지나간 후 10일 이 내에 식물플랑크톤을 번성하게 한다(Chang et al. 1996;
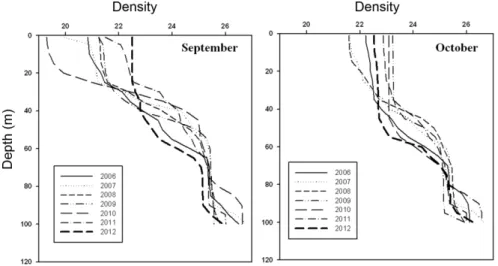
Jang et al. 2013). 본 조사 시기에도 9월 17일 15호 태풍 이 지나가고 난 후 9월 27일에 조사가 이루어져 모든 정 점에서 영양염 증가와 엽록소-a의 증가 및 규조류 우점이 관측되었다. 가을철 2차 식물플랑크톤 대발생은 일반적으 로 바람의 세기가 강해지면서 수직혼합이 활발해지며 성 층 아래에 있던 고영양염이 상층으로 공급되면서 발생한 다. 2006년에서 2011년까지의 정점 M의 조사 결과를 보 면 봄철에 식물플랑크톤의 대발생이 일어난 후 2차 대발 생은 주로 수직혼합층이 60 m까지 확장되는 11월에 나타 났고, 그 규모는 봄철에 나타나는 1차 대발생보다는 적은 규모로 발생하였다(한국해양과학기술원 2011). 하지만 2012년 조사에서는 10월에 정점 P와 M에서 오히려 봄철 보다 높은 값을 보였으며, 이는 다른 연도와 달리 9월과 10월에 수직혼합층이 수심 60 m 부근까지 확장된 것과 관 련이 있다(Fig. 8). 가을철 수직혼합층의 확장은 여름철 성 층아래에 존재하던 영양염이 상층부로 수월하게 공급됨으 로써 식물플랑크톤의 성장을 촉진시킬 수 있다. 2012년 수직혼합층이 확장된 것은 가을철 태풍의 빈도가 잦은 것 과 연관성이 있을 수 있다. 2012년에 8월말부터 10월 중 순까지 7개의 태풍이 조사 지역의 인근을 지나갔다. 특히
Fig. 8. Vertical profiles of density at station M in September and October from 2006 to 2012
8월 30일(14호 태풍)과 9월 17일(15호 태풍) 대한해협에 직접적인 영향을 준 태풍이 통과함으로 수직혼합층이 수 심60m 부근까지 확장되었고, 그 이후에도 태풍들이 일본 남쪽 해역을 통과함으로써 조사 지역에 간접적인 영향을 주어 수직혼합층이 수심 60 m 부근까지 확장/유지된 것으 로 보인다. 하지만 수직혼합층의 확장은 기후변화에 따른 외해수의 세기가 강해져서 나타날 수도 있기 때문에 향후 에도 지속적인 모니터링이 필요하다(Polovina et al. 1995).
정점 R은 10월에 정점 P와 M과 달리 와편모류의 우점율 이 증가하였고, 우점한 종은 Ceratium furca이다. 이 종은 혼합영향을 하는 종으로 연안에서 와편모류 대발생을 일 으키는 종으로 보고되고 있으며(임 등 1993; Baek et al.
2007), 광범위한 수온과 염분 및 광도에서 성장할 수 있는 종으로 알려져 있다(Baek et al. 2007). 수심이 낮은 정점 R은 수층혼합층이가 깊어짐으로써 수층 혼합이 다른 두 정점보다 더 활발하게 일어날수 있어 식물플랑크톤 성장 에 빛이 제한 요인으로 작용할 수 있다. 이러한 결과로 집 괴분석에서 10월에 정점 M과 P가 95%의 유사성을 나타 내었다.
5. 결 론
연구 지역은 연중 대마난류수가 통과하는 해역이지만, 대마난류 및 연안수의 세기가 계절마다 차이를 보이고, 이 로 인해 표층 수괴의 물리·화학적인 요인 및 식물플랑크 톤 군집구조에 시공간적으로 큰 변화를 가져온다. 식물플 랑크톤 그룹은 영양염 공급이 이루어지는 시점에 규조류 가 우점하고, 영양염 공급이 제한되는 시기에는 편모류의 점유율이 증가하였다. 규조류가 우점하는 시기에 대부분 Chaetoceros spp.가 우점하지만, 인산염 농도가 높은 경우 에 S. costatum-like spp.가 우점하였다. 인산염만 제한되 는 정점 R에서 5월에 S. trochoide 가 우점하고 빛이 제한 되는 10월에는 C. furca가 우점하였다. 지속적으로 질산염 과 인산염이 제한되는 정점 M은 Gyrodinium spp.와 Gymnodinium spp.가 우점하는 특징을 보였다. 집괴분석 결과를 보면, 겨울철 수층혼합이 강해지는 시점인 1월과 2월 모든 정점이 95% 이상의 유사성을 보이고, 대마난류 의 세기가 약해지는 3월에는 정점 R과 P, 그리고 다시 대 마난류의 세기가 강해지는 시점인 5월과 6월에는 정점 M 과 P의 유사성이 95% 이상을 나타낸다. 여름철인 8월에 는 정점 R과 P, 그 이후 다시 대마난류의 세기가 강해지면 서 10월에 정점 M과 P가, 그리고 11월에는 모든 정점에 서 95% 이상의 유사성을 보인다. 결과적으로 대마난류의 계절적 세기 변화와 영양염 농도 변화가 연구 지역의 식 물플랑크톤 군집구조의 변동을 유발할 수 있음을 암시하 고 있다. 이러한 경향은 향후 기후변화가 대마난류의 계절
적 세기와 영양염 특성의 변화를 가져 올 경우 남해동부 해역에 식물플랑크톤의 일차생산력 및 군집구조의 변화를 가져올 수 있음을 시사한다.
사 사
본 연구는 국가 R&D 과제인 “기후변화가 남해해양생 태계에 미치는 영향 및 기능평가 기술 개 사업(PM57090)”
과 한국해양연구원의 연구사업인 “북서태평양의 해양환경 변화가 한반도 주변해(대한해협)에 미치는 영향 연구 (PE99162)”의 일환으로 수행되었습니다.
참고문헌
김일남, 이동섭 (2004) OMP 방법으로 분석한 하계 동해의 수계 특성. Ocean and Polar Res 26(4):581-594
민흥식, 박재훈, 최아라, 박영규, 신경순, 장풍국 (2011) 2006~2010년 기간 동안 대한해협 서수로에서 관측된 해 수 물성의 변동. Ocean and Polar Res 33(3):325-336 승영호 (1992) 한반도 주변의 수괴와 해수순환. 한국해양학
회지 「바다」 27:324-331
양영진, 김상현, 노홍길 (1998) 한국 남·서해 및 동중국해 북 부해역에 출현하는 수온전선. 한국수산학회지 32(5):618- 623
이동섭 (1999) 여름과 겨울철 남해의 영양염 분포 특성. 한국 해양학회지 「바다」 4(4):371-382
임월애, 김학균, 박주석, 이원재 (1993) 적조와편모조 Scrippsiella trochoidea와 해양세균 Pseudomonas spp.의 동시배양시 지방산 조성의 변화. 한국수산과학회지 26(2):
198-203
장풍국, 장민철, 이우진, 신경순 (2010) 장목만에서 여름철 영 양염 특성 변화가 식물플랑크톤 군집구조에 미치는 영향.
Ocean and Polar Res 32(2):97-111
정익교, 강윤향, 김영진, 권오섭 (1998) 한국 연안 초미세플랑 크톤 생태: I. 대한해협 서수도 해역. Algae 13(1):101- 107
조양기, 김구, 노홍길 (1994) 여름철 남해 저온수의 특성과 기원. 한국해양학회지 「바다」 29:414-421
한국해양과학기술원 (2011) 북서태평양이 한반도 주변해에 미치는 영향 연구. 한국해양과학기술원, BSPE 98563- 10040-1, 458 p
Arrigo KR (2004) Marine microorganism and global nutrient cycles. Nature 437(7075):349-355
Back SH, Shimode S, Kikuchi T (2007) Reproductive ecology of the dominant dinoflagellate, Ceratium fusus, in coastal area of Sagami Bay, Japan. J Oceanogr 63:35-45 Bouamm HA, Nakane T, Oka K, Nakata K, Kurita K,
Sathyendranath S, Platt T (2010) Environmental controls
on phytoplankton production in coastal ecosystems: a case study from Tokyo Bay. Estuar Coast Shelf S 87(1):
63-72
Chang J, Chung CC, Gong GC (1996) Influences of cyclones on chlorophyll a concentration and Synechococus abundance in a subtropical western Pacific coastal ecosystem. Mar Ecol-Prog Ser 140:199-205
Chang PH, Isobe A (2003) A numerical study on the Changjiang diluted water in the Yellow and East China Seas. J Geophys Res 108(C9):3299. doi:10.1029/
2002JC001749
Cupp EE (1943) Marine plankton diatoms of the west coast of North America. University of California press, Berkeley and LA, 237 p
De Baar HJW (1994) von Liebig's low of the minimum and plankton ecology (1899-1991). Prog Oceanogr 33(4):347- 386
Dodge JD (1982) Marine dinoflagellate of the British Isles.
Her Majesty stationary office, London, 303 p
Dortch Q, Whitledge TE (1992) Does nitrogen or silicon limit phytoplankton production in the Missippi River plume and nearby regions? Cont Shelf Res 12(11):1293- 1309
Finkel ZV, Irwin AJ, Schofield O (2004) Resource limitation alters the 3/4 size scaling of metabolic rates in phyto- plankton. Mar Ecol-Prog Ser 273:269-279
Fu FX, Warner ME, Zhang Y, Feng Y, Hutchins DA (2007) Effect of increased temperature and CO2 on phytosynthesis, growth, and elemental ratios in marine Synechococcus and Prochlorococcus (Cyanobacteria). J Phycol 43:485- 496
Gong GC, Chen YLL, Liu KK (1996) Chemical hydrography and chlorophyll-a distribution in the East China Sea in summer: implications in nutrient dynamics. Cont Shelf Res 16:1561-1590
Guo X, Miyazawa Y, Yamagata T (2006) The Kuroshio onshore intrusion along the shelf break of the East China Sea: The origin of the Tsushima Warm Current. J Phys Oceanogr 36(12):2205-2231
Isobe A (1999) On the origin of the Tsushima Warm Current and its seasonality. Cont Shelf Res 19(1):117-133 Jang P-G, Shin HH, Baek SH, Jang MC, Lee TS, Shin K
(2013) Nutrient distribution and effects on phytoplan- kton assemblages in the western Korea/Tsushima Strait.
New Zeal J Mar Fresh 47(1):21-37
Kim D, Choi SH, Kim KH, Shim JH, Yoo SJ, Kim CH (2009) Spatial and temporal variations in nutrient and chlorophyll-a concentrations in the northern East China Sea surrounding Cheju Island. Cont Shelf Res 29(11-
12):1426-1436
Lagus A, Suomela J, Weithoff G, Heikkila K, Helminen H, Sipura J (2004) Species-specific differences in phytoplankton responses to N and P enrichments and the N:P ratio in the Archipelago Sea, northern Baltic Sea. J Plankt Res 26(7):779-798
Li G, Han X, Yue S, Wen G, Rongmin Y, Kusky TM (2006) Monthly variations of water masses in the East China Sea. Cont Shelf Res 26(16):1954-1970
Morimoto A, Takikawa T, Onitsuka G, Watanabe A, Moku M, Yanagi T (2009) Seasonal variation of horizontal material transport through the eastern channel of the Tsushima Straits. J Oceanogr 65(1):61-71
Officer CB, Ryther JH (1980) The possible importance of silicon in marine eutrophication. Mar Ecol-Prog Ser 3:83-91
Onitsuka G, Yanagi T, Yoon J-H (2007) A numerical study on nutrient sources in the surface layer of the Japan Sea using a coupled physical-ecosystem model. J Geophys Res 112(C5):C05042
Park Mo, Park JS (1997) HPLC method for the analysis of chlorophylls and carotenoids from marine phytoplankton.
J Korean Soc Oceanogr 32:46-55
Parsons TR, Maita Y, Lalli CM (1984) A manual of chemical and biological methods for seawater analysis.
Pergamon, New York, p 173
Polovina JJ, Mitchum GT, Evans GT (1995) Decadal and basin-scale variation in mixed layer depth and the impact on biological production in the Central and North Pacific. Deep-Sea Res 42(10):1701-1716
Raven JA (1998) The twelfth Transley Lecture. Small is beautiful: the picophytoplankton. Funct Ecol 12:503-513 Rines JE, Hargraves PE (1988) The Chaetoceros Ehrenberg
(Bacillariophyceae) Flora of Narragansett Bay, Rhode Island, USA. Bibl Phycol 79:1-196
Sommer U (1994) The impact of light intensity and daylength on silicate and nitrate competition among marine phytoplankton. Limmol Oceanogr 39(7): 1680- 1688
Song X, Huang L, Zhang J, Huang X, Zhang J, Yin J, Tan Y, Liu S (2004) Variation of phytoplankton biomass primary production in Daya Bay during spring and summer. Mar Pollut Bull 49:1036-1044
Sournia A (1978) Phytoplankton manual. In : Monographs on Oceanographic Methodology Vol. 6. UNESCO, Paris, 337 p
Teague WJ, Jacobs GA, Ko DS, Tang TY, Chang KI, Suk MS (2003) Connectivity of the Taiwan, Cheju, and Korea Straits. Cont Shelf Res 23:63-77
Tilman D (1982) Resource Competition and Community Structure. Princeton University Press, Princeton, New Jersey, 296 p
Yamaji I (1979) Illustrations of the Matine Plankton of Japan. Hoikusha Publishing Co., Ltd., Osaka, Japan, 537 p
Yanagi T (2002) Water, salt, phosphorus and nitrogen budgets of the Japan Sea. J Oceanogr 58(6):797-804
Yin K, Song XX, Liu S, Kan J, Qian PY (2008) Is inorganic nutrient enrichment a driving force for the formation of
red tide? A case study of the dinoflagellate Scrippsillea trochoidea in an embayment. Harmful Algae 8:54-59 Yun JY, Magaard L, Kim K, Shin CW, Kim C, Byun SK
(2004) Spatial and temporal variability of the North Korean Cold Water leading to the near-bottom cold water intrusion in Korea Strait. Prog Oceanogr 60:90-131
Received Apr. 2, 2013 Revised May 31, 2013 Accepted Jun. 17, 2013