200
[Note]
아산만 연안하구 식물플랑크톤의 2006년 하계 종조성 변화
이향화·신용식*·양성렬1·박 철2
목포해양대학교 해양시스템공학부
1
광주대학교 공과대학 환경공학과
2
충남대학교 자연과학대학 해양학과
Summer Dynamics of Phytoplankton Taxonomic Composition in a Coastal Estuarine System of Asan Bay
HYANGHWA YI, YONGSIK SHIN*, SUNGRYULL YANG
1AND CHUL PARK
2Division of Ocean System Engineering, Mokpo National Maritime University, Mokpo 530-729, Korea
1
Department of Environmental Engineering, College of Engineering Kwangju University, Kwangju 503-703, Korea
2
Department of Oceanography, College of Natural Sciences, Chungnam National University, Deajeon 305-764, Korea
아산만 식물플랑크톤의 하계 종조성 변화를 조사하기 위하여 2006년 6월, 7월, 8월에 매달 두 번씩 실시하였으며 조사 정점은 공간적 분포를 나타내는 5개 정점을 선정하여 조사하였다. 6월과 7월에 염분은 내만에서 감소하는 경 향을 보였다. 영양염은 6월과 7월에 높았으나 8월에 낮은 경향을 보였다. 2006년 여름철 아산만에서 식물플랑크톤 군집은 규조류(62.8%), 와편모조류(17.3%), 은편모조류(14.8%), 유글레나류(1.0%), 남조류(0.9%), 녹조류(0.4%) 등 6종과 확인되지 않은 종(2.8%)이 포함되었다. 6월에는 와편모조류(주요하게 Prorocentrum sp.(29.6%))가 우점 하였 고 총 현존량의 43.5%를 차지하였지만 7월과 8월에는 규조류(주요하게 Leptocylindrus sp.(21.4%), Chaetoceros
sp.(27.6%))가 우점 하였으며 각각 총 현존량의 69.1%, 89.9%를 차지하였다. 이와 같이 하계에도 강우에 따라 아산 만 각 정점에서의 우점속의 변화가 발생하였으며 이는 강우에 따른 담수유입이 하계 아산만 식물플랑크톤 종조성 변화에 영향을 미치고 있음을 암시한다.
Phytoplankton community was investigated in Asan Bay, South Korea. Samples were collected at 5 stations along Asan Bay axis during wet season from June to August, 2006. In June and July, salinity decreased espe- cially at inside stations. Nutrients were high in June and July, however, decreased in August. We observed the community of phytoplankton including diatoms(62.8%), dinoflagellates(17.3%), cryptophytes(14.8%), eugle- nophytes(1.0%), cyanophytes (0.9%), chlorophytes(0.4%), and some of unidentified taxa(2.8%) during summer 2006 in Asan Bay. In June, dinoflagellates (mainly Prorocentrum sp.(29.6%)) were dominated, accounting for about 43.5% of total cell number, whereas in July and August diatoms (mainly Leptocylindrus sp.(21.4%), Chaetoceros sp.(27.6%)) were dominated occupying 69.1% and 89.9%, respectively. The results suggest that freshwater inputs affected phytoplankton community in the Asan Bay ecosystem.
Keywords: Asan Bay, Prorocentrum sp., Leptocylindrus sp., Chaetoceros sp., phytoplankton community
서 론
식물플랑크톤은 수중 생태계의 일차생산자로서 , 수질환경변화
에 민감하며 수환경의 변화 양상 파악을 위한 요소 중의 하나로
중요한 역할을 한다 ( 정 등 , 2001). 식물플랑크톤 군집은 수중 내
자체유기물 생산의 중요한 역할을 하고 , 수중 생태계 다른 구성요
인들의 변화에 따라 종천이 및 현존량의 변화를 일으키며 (James,
1979), 특히 수온 , 염분 , 영양염 등과 같은 요인들에 의해 군집구
조 및 우점종 천이를 나타내는 것이 일반적이다 (Reynolds et al ., 1983; Sommer et al. , 1986).
식물플랑크톤의 현존량 그리고 우점종은 상향식과 하향식조절 인자에 의해서 민감하게 변화하기 때문에 수질에 대한 지표로 의
미가 있다 (UNESCO, 1978). 식물플랑크톤의 종 구성이나 현존량
은 환경의 시공간 변화에 따라 다르게 나타났으며 ( 문과 최 , 1991;
김 등 , 1998; 이과 부 , 1998), 염분 차이가 큰 하구역에서는 염분
과 계절에 따라서도 종의 구성이 다르게 나타났다 (Marshall and
Raymond, 1990; 심 등 , 1991; 권 등 , 2004). 따라서 종의 구성이 바뀌었다는 것은 수온 , 수괴의 안정성 등의 환경인자가 변화하였 음을 의미하기도 한다 .
식물플랑크톤 종은 계절과 공간에 따라 변동하는데 일반적으로 여름에 와편모조류가 우점하고 겨울에 규조류가 우점한다 . 와편모
*Corresponding author: [email protected]
조류는 해양생태계에서 규조류 다음으로 해양 식물플랑크톤의 중 요한 구성요인이다 ( 심 등 , 1981). 따라서 와편모조류의 분류는 식
물플랑크톤을 연구하는데 중요하고도 기본적인 연구과제이다 . 우
리나라에서 해산와편모조류에 관한 기록은 박 (1956) 이 한국해협
에서와 하기 남해안에서의 플랑크톤조성을 연구한 결과 Ceratium
속 및 Peridinium 속의 몇몇 종이 처음으로 기록되었다 . 그 이후에
유 (1960) 에 의해서 용호만에서 이매패인 불통의 식이 연구결과
Dinophysis sp. 와 Prorocentrum sp. 의 기록을 남겼으며 박과 김
(1967) 에 의해 진해만에서 Gymnodinium sp. 와 Noctiluca sp. 가 기
록되었다 . 최 (1967) 는 한국연안수역에서 Pyrophacus sp. 를 관찰하
였고 김과 조 (1970) 는 금강하구에서 Glenodinium foliaceum 의 출 현을 기록했으며 심과 이 (1979) 에 의해서 천수만에서 Gonyaulax polygramma 와 Gonyaulax Polyedra 가 기록되었다 . 우리 나라에서 적조를 일으키는 식물플랑크톤은 주로 와편모조류와 규조류였는 데 특히 우리 나라 연안에서 수산생물을 직접 죽일 수 있는 와편 모조류에 속하는 Gymnodinium mikimotioi 란 종이 1981 년도에 가 장 먼저 출현하였고 Cochlodinium polykrikoides 란 종은 1982 년도
부터 매년 수산피해를 일으키고 있으며 그리고 1992 년도에는 Gyrodinium sp. 라는 세계 신종이 수산피해를 일으켰다 ( 김과 주 , 2000). Alexandrium tamarense 는 Hiroshima 만에서 처음 적조를 발
생시킨 이래 매년 Hiroshima 의 굴 양식장에 패독을 발생시켜 많
은 피해를 주고 있다 (Asakawa et al ., 1993; 1995).
지금까지 아산만 내 크기구조를 고려한 군집구조와 우점속에 관
한 연구 ( 이 등 , 2005) 가 이루어졌지만 여름철에 강우에 따른 식물
플랑크톤의 생태지수 변화와 우점속 및 환경변화와의 연관성에 대 한 연구는 미흡하다 . 따라서 본 연구는 풍수기인 하계의 식물플랑
크톤 군집 동역학을 분석함으로써 아산만 식물플랑크톤 생태 연 구의 기초자료로 활용하고자 하였다 .
재료 및 방법 조사지역
조사 해역인 아산만은 길이 40 km, 너비가 2.2 km 이며 , 평균조
차가 6.1 m 최대 9.6 m 에 달하는 우리나라에서 조석 간만의 차가
큰 개방형만으로서 , 겨울에는 대륙성 기후 , 여름에는 해양성 기후
의 영향이 뚜렷하다 . 그리고 삽교 , 아산 및 대호 방조제등이 건설
되었으며 , 앞으로도 연안유역을 따라서 대규모 국가 산업단지가
들어설 전망이어서 많은 연안환경의 변화가 예상되어지는 해역이 다 . 특히 방조제 건설이후 우기에는 저장되었던 담수가 일시에 방 출되어 만내 환경 및 생물상에 영향 미칠 것이라 사료된다 ( 문 등 , 1993). 조사일자는 2006 년 6 월 16 일 , 6 월 23 일 , 7 월 13 일 , 7 월 24
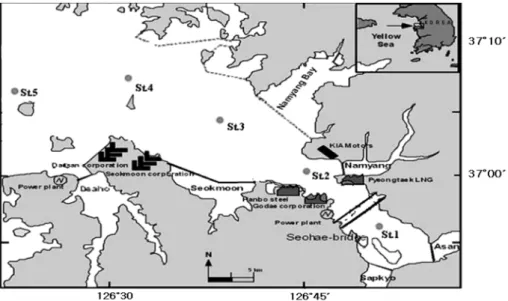
일 , 8 월 11 일 , 8 월 25 일로 월 2 회 조사하였으며 조사 정점은 공간 적 분포를 나타낼 수 있는 5 개 정점을 선정하여 조사하였다 (Fig. 1).
조사방법
채수는 5 l 니스킨 (Niskin) 채수기와 반돈 (Van Don water sampler) 채수기를 이용하여 해수 표면 1 m 아래 수심 ( 표층 ) 에서 채 수하였다 .
조사기간 동안 강우량 및 일조시간은 천안시 기상청 자료를 이 용하였고 강의 방출량은 삽교로부터 방출되는 양을 이용하였다 .
수온 , 염분 (Salinity) 및 용존산소는 YSI-Model 85 S-C-T 측정
장비를 이용하여 , 수면에서 1 m 아래 수심에서 측정하였으며 , 투
명도는 Secchi disk 를 이용하여 측정하였다 .
영양염은 Niskin 채수기를 이용하여 시료를 채수한 후 일정량
을 Whatman
®25 mm GF/F filter 를 통과시켜 여액을 20 ml plastic scintillation vial 에 넣어 dry ice 에 냉동 운반하여 -20
oC 냉동보관 후 Parsons et al. (1984) 에 준하여 Autoanalyzer(Bran Luebbe
®) 를 이용하여 측정하였다 .
식물플랑크톤을 2 개의 크기 즉 대형식물플랑크톤 (Netplankton
> 20 µ m), 소형식물플랑크톤 (Nanoplankton < 20 µ m) 으로 분류하
였는데 이를 위해 20 µ m Nytex
®mesh 를 이용하였다 . 채수된 해 수 시료 적당량 (100 ml) 을 20 µ m Nytex
®mesh 로 거른 후 펌프
(<120 mmHg) 를 사용해서 여과 되어진 여액을 유리 섬유 여과지
(GF/F filter, 직경 25 mm, Whatman
®, pore size 0.7 µ m) 로 여과 하였으며 , 전체클로로필 (chlorophyll a ) 은 채수된 해수시료 100 ml
를 유리 섬유 여과지로 필터 하였다 . 사용된 필터들을 클로로필
추출용액인 90% 아세톤 (Acetone) 8 ml 가 담긴 차광 시험관에 넣
고 12 시간이 지난 후 24 시간 이내로 Turner Designs
®10-AU
Fig. 1. Sampling stations in the Asan
Bay.
fluorometer 를 사용하여 chlorophyll a 를 측정하였다 . 한 두 방울
의 HCl(2N) 를 추가해 산성화에 따른 pheopigments 의 양을 측정
하였다 . 일반적으로 포식자들이 식물플랑크톤을 섭식한 후 혹은
자가 산화로 인해 클로로필 a 가 pheopigments 로 전환되기 때문에
클로로필 a 와 pheopigments 의 비를 통해 섭식의 활발도 ( 포식압 )
내지는 자가 산화 정도를 간접적으로 측정할 수 있다 (Welschmeyer and Lorenzen, 1985).
식물플랑크톤의 계수 및 동정을 위해 Lugol’s solution(Sourinia, 1978) 으로 고정시킨 후 Sedgewick-Rafter Chamber 에 넣어 계수
하였다 . 현장에서 1 l 채수병에 해수 시료 900 ml 와 Lugol’s solution
5 ml 를 넣어 잘 혼합하여 암실에서 약 2-3 일 정도 자연 침전시킨
다 . 순환 펌프를 이용해서 자연 침전 시킨 시료의 상등액을 제거 하고 , 남은 여액 200 ml 를 잘 혼합한 후 , 50 ml 용 시험관 (Conical
tube) 에 넣고 약 1-2 일 정도 농축시킨 다음 다시 45 ml 상등액을
제거하고 남은 5 ml 중 1 ml 를 S-R Chamber 에 넣고 계수하였다 .
동정은 영상 분석 장치 시스템 (colored image analyzer) 을 갖춘
Axioskop 2 MAT(ZEISS) 를 이용하였다 . 군집구조의 분석은 조사
정점별로 출현한 분류군의 수를 비교하여 출현 개체수 , 우점종 , 군
집지수 - 종다양성지수 (i.e. Shannon-Weiner index), 풍부도지수 (i.e.
Margalef index), 균등도지수 (i.e. Pielou index), 우점도지수 (i.e.
Simpson index) 를 계산하였으며 , 각 지수는 다음식에 의해 산출하
였다 (Simpson, 1949; Margalef, 1958; Whittaker, 1972; Ludwig and Reynolds, 1988).
Shannon-Weiner index:
Margalef index:
Pielou index:
Pi: i 번째 종의 점유율
S: 출현종수
Simpson index:
N: 총 개체수
n
i: i 번째 우점종의 개체수
결과 및 고찰
환경요인 2006 년 아산만의 강우량 분포를 보면 갈수기인 가을과 겨울에 적고 풍수기인 여름에 많았으며 7 월에 가장 많은 경향을 보였다
(Fig. 2A). 월 합 일조시간은 대체적으로 봄과 가을에 길고 여름과
겨울에 짧은 계절적 분포를 나타내었다 (Fig. 2A). 2006 년 아산 및
삽교 방조제로부터 만내로 유입되는 담수 유입량은 장마 등에 의 해서 강우량이 많았던 7 월에 4994.0( 천톤 ) 이었고 6 월에는 1389.8 ( 천톤 ) 이 유입되었다 (Fig. 2B).
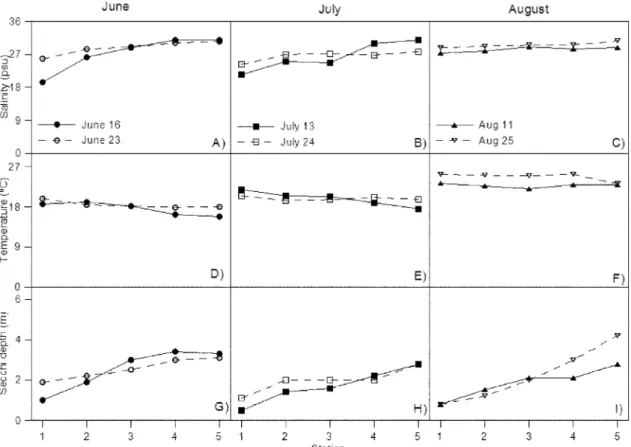
조사기간 동안 염분 변화를 보면 6 월 16 일에 19.4-31.0 psu, 6
월 23 일에는 25.9-30.5 psu 를 나타내었고 7 월 13 일에 21.4-30.9 psu, 7 월 24 일에는 24.2-27.7 psu 를 나타내었다 (Fig. 3A-B). 8 월에는 각 각 27.4-28.9 psu. 28.7-30.7 psu 를 나타내었고 (Fig. 3C) 대부분 2 차 조사에서 1 차 조사 시보다 높은 경향을 나타내었는데 특히 강
우량이 많은 7 월에 가장 낮은 분포를 보였다 (Fig. 3A-C). 염분의
공간적 분포를 보면 내측해역에서 낮고 외측해역으로 갈수록 증
가하는 경향을 보였는데 (Fig. 3A-C) 이는 담수 유입의 영향임을
알 수 있다 .
수온의 변화를 보면 6 월에는 각각 15.9-19.1
oC, 17.9-19.9
oC 를 나타내었고 7 월에는 각각 17.5-21.9
oC, 19.5-20.6
oC 를 나타냈으
며 8 월에는 각각 22.0-23.3
oC, 23.3-25.4
oC 를 나타내었는데 시간
적으로 증가하는 경향을 보였다 (Fig. 3D-F). 공간적으로는 거의 외
측해역이 내측해역보다 높은 분포를 보였다 (Fig. 3D-F).
투명도는 조사기간 동안 외측해역으로 갈수록 증가하는 경향을 보였고 , 6 월에는 각각 1.0-3.4 m, 1.9-3.1m 의 범위를 나타내었고
7 월에는 각각 0.5-2.8 m, 1.1-2.8 m 의 범위를 나타내었으며 8 월에 는 각각 0.8-2.8 m, 0.8-4.2 m 의 범위를 나타내었다 (Fig. 3G-I).
H ′ P
i* i 0=∑
sln ( ) P
i=
S S = ( – 1 ) ⁄ ln ( ) N J H = ′ ⁄ ln ( ) S
D n
i( n
i– 1 )
N N ( – 1 ) ---
i 0=
∑
s=
Fig. 2. Photo-period (monthly sum), precipitation(daily and monthly sum) and freshwater river discharge (monthly sum) from the Asan and
Sapkyo embankment.
7 월과 8 월의 내측해역에서 낮은 투명도는 담수의 유입에 의한 탁
도 물질 증가에 의한 것으로 사료된다 .
용존산소농도는 6 월에 각각 6.6-8.2 mg l
-1, 5.7-8.5 mg l
-1의 범위 를 나타냈고 7 월에는 각각 5.3-7.6 mg l
-1, 5.9-8.1 mg l
-1의 범위를 나타냈으며 8 월에는 각각 7.5-9.8 mg l
-1, 8.6-10.2 mg l
-1의 범위를 나타내었다 (Fig. 4A-C). 공간적으로 6 월과 7 월에 내측해역에서 낮 은 분포를 보였다 .
영양염의 농도변화를 보면 6 월에는 암모니아를 제외하고 모두
2 차 조사에서 높은 농도를 보였고 7 월과 8 월에는 반대인 경향을
보였으며 모두 내측해역에서 외측해역으로 갈수록 감소하는 경향
을 보였다 . 8 월의 영양염 농도가 6, 7 월에 비해 상대적으로 낮았
는데 이는 강우의 영향으로 판단된다 (Fig. 4). 6 월과 7 월의 암모니
아 농도는 외측해역으로 갈수록 감소하는 경향을 보였고 8 월에는 내측해역이 외측해역보다 다소 높은 농도를 보였지만 공간적으로
큰 차이를 보이지 않았다 (Fig. 4D-F). 아질산 + 질산과 인산염의 농
도는 8 월을 제외하고 외측해역으로 갈수록 감소하는 경향을 보였
으며 (Fig. 4G-L), 규산염의 농도 역시 8 월을 제외하고 조사기간 동
안 외측해역으로 갈수록 감소하는 경향을 보였다 (Fig. 4M-O). 공 간적으로 저염분 분포인 내측해역에서 고농도의 영양염 분포는 담
수 유입의 증가 ( 현 , 2006) 에 따라 영양염이 공급되었기 때문으로
판단된다 . 담수의 유입에 따른 영얌염의 공급은 여수 돌산 해역
( 이 , 2002) 과 섬진강 하구역 ( 권 등 , 2004) 에서도 보고되었다 . Chlorophyll a : Pheopigments ratio 는 환경 악화로 인한 식물플
랑크톤의 노화 혹은 식물플랑크톤의 포식자인 동물플랑크톤의 배
설물 등에서 생성되며 식물플랑크톤의 포식율 (grazing rate) 을 간
접적으로 알아보는 지표로 사용되어진다 . Ratio 가 낮을수록 포식
율이 높고 환경악화로 인한 식물플랑크톤의 사멸이 많다는 것을 의미하는데 일반적으로 순수한 Chlorophyll a 의 경우 비율은 2.2
이며 (Holm-Hansen et al ., 1965), 생리적으로 활발하게 성장하는 식물플랑크톤인 경우 보통 1.7 이상으로 알려져 있다 (Yentsch and
Menzel, 1963). 즉 담수의 유입에 의한 탁도의 증가 , 급속한 염분
의 변화 및 유속 (flushing rate) 의 증가는 식물플랑크톤의 서식 환
경을 악화 시켜서 식물플랑크톤의 성장을 생리적으로 억제할 것 으로 사료된다 . 실제로 조사 기간 동안 Chl:Pheo 가 8 월 11 일에
1.16-1.92 의 범위 (Fig. 5E) 로 다른 조사 시보다 낮게 나타났고 식
물플랑크톤 생체량도 높지 않으며 내측해역에서 염분도 다소 높 아진 것을 보아 식물플랑크톤에 대한 섭식보다 환경 악화에 의한 영향이 컸을 것으로 판단된다 . 6 월 16 일 정점 3 의 대형식물풀랑 크톤 분포를 제외하고 크기별 식물플랑크톤의 공간적인 변화가 작 게 나타났다 (Fig. 5A). 6 월 23 일 , Chl:Pheo 의 변화를 보면 내측해
역 정점 1, 2 는 소형식물플랑크톤이 높게 나타났고 외측해역으로
갈수록 대형식물플랑크톤이 높은 경향을 보였으며 소형식물플랑 크톤은 공간적인 변화가 작았다 . 대형식물플랑크톤은 정점 3 에서 가장 높았고 , 정점 1 에서 가장 낮았다 (Fig. 5B). 7 월 13 일에 Chl:Pheo
의 변화는 6 월 16 일에 Chl:Pheo 의 변화와 비슷한 경향을 보였다
(Fig. 5C). 7 월 23 일 대형식물플랑크톤의 Chl:Pheo 는 정점 3 에서
가장 높았고 , 정점 2 에서 가장 낮았으며 소형식물플랑크톤의
Chl:Pheo 는 외측해역으로 갈수록 감소하는 경향을 나타냈다 (Fig. 5D).
Fig. 3. Spatial variations of salinity, water temperature, secchi disk depth during June, July and August in the Asan Bay.
8 월 11 일에 대형식물플랑크톤 Chl:Pheo 는 7 월 23 일에서의 변화와 비슷한 경향을 나타내었고 소형식물플랑크톤의 Chl:Pheo 는 외측
해역으로 갈수록 다소 증가하는 경향을 보였다 (Fig. 5E). 8 월 23
일 대형식물플랑크톤의 Chl:Pheo 는 내측해역이 다소 높은 경향을
보였고 소형식물플랑크톤의 Chl:Pheo 는 공간적인 변화가 작게 나 타났다 (Fig. 5F). 조사기간 동안 Chl:Pheo 는 담수 유입 시 낮은 분 포를 보였지만 대부분 1.7 이상으로 식물플랑크톤의 성장에 양호 한 환경임을 알 수 있다 . 일부시기에 정점 3 에서 대형식물플랑크
톤의 Chl:Pheo 가 높았으나 동시에 생체량도 높게 나타났으며 (Fig. 6)
종조성에서도 대형크기 (micro-sized) 인 사상군체의 Skeletonema costatum , Leptocylindrus sp . , Chaetoceros sp . 가 우점했던 것으로
보아 낮은 포식압이 정점 3 에서 높은 net-size 의 chl:pheo 에 영향
을 미쳤을 것으로 예상된다 . 특히 8 월에는 Chl:Pheo 의 공간분포
는 생체량의 분포와 서로 상반되는 결과를 보였는데 이는 포식압 이나 기타 환경인자들이 생체량 분포에 미친 영향을 암시한다고 할 수 있다 .
크기별 식물플랑크톤 생체량 및 기여율 분포
6 월 16 일에 전체식물플랑크톤 (whole-chlorophyll a ) 과 소형식물 플랑크톤 (nano-chlorophyll a ) 의 생체량은 내측해역에서 정점 3 까 지 증가하다가 외측해역으로 갈수록 감소하는 경향을 보였다 (Fig.
6A, 6C). 대형식물플랑크톤 (net-chlorophyll a ) 의 생체량 분포를 보
Fig. 4. Spatial distributions of dissolved oxygen, ammonia, nitrite+nitrate, phosphate and silicate in the surface water during June, July,
August in the Asan Bay.
Fig. 5. Spatial variations of size-fractionated chlorophyll
a: pheopigments during June, July, August in the Asan Bay.
Fig. 6. Spatial variations of size-fractionated chlorophyll
a(whole, net- and nano-sized) in the surface water during June, July, August in
the Asan Bay.
면 정점 3 에서 가장 높았고 다른 정점에서는 비슷한 분포를 보였 다 (Fig. 6B). 6 월 16 일에 정점 3 을 제외하고 소형식물플랑크톤의
기여율이 높은 공간적인 분포변화를 보였다 (Fig. 7A).
6 월 23 일에 전체식물플랑크톤 생체량 분포는 6 월 16 일과 반대
인 경향을 나타내었다 (Fig. 6A). 대형식물플랑크톤 생체량은 공간
적으로 변화가 작았으며 (Fig. 6B) 소형식물플랑크톤 생체량은 6 월
16 일에 비해 증가하였고 공간적으로도 외측해역으로 갈수록 다소
증가하는 경향을 보였다 (Fig. 6C). 이 시기 대형식물플랑크톤의 기
여율은 역시 낮았고 소형식물플랑크톤의 기여율이 높게 나타났다
(Fig. 7B). 정점 3 에서 6 월 16 일에 대형식물플랑크톤의 기여율이
높다가 6 월 23 일에 소형식물플랑크톤의 높은 기여율은 정점 3 의
전체 클로로필 농도가 6 월 16 일 6.82 µ g l
-1에서 23 일에 3.88 µ g l
-1로 감소한 영향도 있으나 소형식물플랑크톤의 농도가 1.42 µ g l
-1에서
1.78 µ g l
-1로 증가하였기 때문이다 .
7 월 13 일에는 전체식물플랑크톤의 생체량은 2.02-2.87 µ g l
-1의 범위로 6 월 비해 낮게 나타났고 전체식물플랑크톤과 대형식물플 랑크톤은 공간적인 변화가 크게 나타나지 않았으며 소형식물플랑 크톤은 내측해역에서 정점 3 을 기준으로 증가하다가 감소하는 경
향을 보였다 (Fig. 6D-F). 대형식물플랑크톤의 기여율은 외측해역
에서 높은 분포를 보였고 소형식물플랑크톤의 기여율이 높게 나 타났으며 공간적으로 내측해역에서 다소 높은 분포를 보였다 (Fig. 7C).
7 월 24 일 식물플랑크톤 생체량은 6 월과 7 월 13 일에 비해 모두
높게 나타났다 (Fig. 6D-F). 전체식물플랑크톤과 대형식물플랑크톤
은 외측해역에서 높게 나타나는 경향을 보였고 소형식물플랑크톤
은 정점 1 에서 높은 분포를 보였다 (Fig. 6E-H). 기여율의 분포를
보면 전반적으로 소형식물플랑크톤이 높은 경향을 나타냈으며 외
측해역에서 대형식물플랑크톤의 기여율도 상대적으로 높게 나타 났다 (Fig. 7D).
8 월 11 일 전체식물플랑크톤과 대형식물플랑크톤 생체량은 외측
해역으로 갈수록 증가하는 경향을 보였고 소형식물플랑크톤은 공
간적인 변화를 보이지 않았다 (Fig. 6G-I). 기여율의 분포를 보면
대형식물플랑크톤의 기여율이 우점 하였고 외측해역으로 갈수록
증가하는 경향을 보였다 (Fig. 7E).
8 월 25 일 식물플랑크톤은 높은 생체량을 보였고 전체식물플랑 크톤과 대형식물플랑크톤 생체량은 8 월 11 일 조사 시와 반대로 외
측해역으로 갈수록 감소하는 경향을 보였고 소형식물플랑크톤의
생체량은 역시 공간적인 변화를 보이지 않았다 (Fig. 6G-I). 정점 1
의 높은 생체량은 7 월말에 많은 강우와 삽교로부터 유입된 담수 의 유입 때문일 것이라고 판단된다 . 대형식물플랑크톤의 기여율은 내측해역에서 80% 이상으로 높은 분포를 보였고 생체량 분포와 마찬가지로 외측해역으로 갈수록 감소하는 경향을 보였으며 반면 소형식물플랑크톤의 기여율은 외측해역으로 갈수록 증가하는 경 향을 보였다 (Fig. 7F).
시간적인 변동을 보면 전체식물플랑크톤의 생체량은 8 월에 높
은 농도를 나타냈으며 , 대형식물플랑크톤은 역시 기여율이 높았던
8 월에 높은 생체량을 보였고 소형식물플랑크톤은 6, 7 월에 비슷한 생체량을 나타내었다 (Fig. 6).
식물플랑크톤 우점속 및 그룹 분포
조사기간 동안 총 6 개의 분류군 - 규조류 (Diatoms), 은편모조류
(Cryptophytes), 남조류 (Chlorophytes), 녹조류 (Cyanophytes), 유글
레나류 (Euglenophytes), 와편모조류 (Dinoflagellates) 중에서 6 월에
Fig. 7. Spatial variations of percentage contribution(%) of size-fractionated class (net-, nano-sized) to the total chlorophyll
ain the surface
water during June, July, August in the Asan Bay.
와편모조류 (Dinoflagellates) 가 우점 하였고 7 월 , 8 월에는 규조류
(Diatoms) 가 높은 우점율을 보였는데 특히 8 월에는 85% 이상으로
높았다 (Table 1).
6 월에는 6 월 23 일에 암모니아 , 아질산염 + 질산염 , 인산염 , 규산 염의 농도가 6 월 16 일 조사 시보다 높은 경향을 보였지만 7 월에 는 7 월 13 일에 영양염 농도가 7 월 24 일 조사 시보다 높은 경향을 보였으며 모든 시기에 내측해역에서 외측해역으로 갈수록 감소하
는 경향을 보였다 (Fig. 4). 그리고 내측해역에서의 낮은 염분 분포
를 보면 퇴적층에서 박테리아와의 광물질화작용에 의한 영향 보 다는 조사 며칠 전부터 내린 강우에 의한 육상기원 영양염의 유
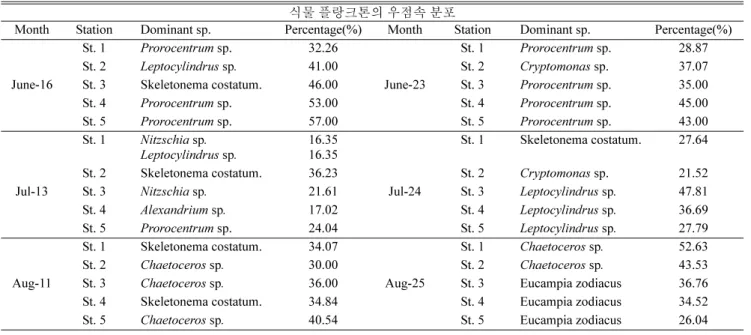
입된 결과라고 사료된다 ( 현 , 2006). 6 월 16 일에 전체식물플랑크톤
과 대형식물플랑크톤의 생체량이 높았던 정점 3 에서 굵고 기다란 주변 돌기환으로 세포들이 연결되어 사상군체를 이루는 Skeletonema costatum, 정점 2 에서 Leptocylindrus sp . 가 우점으로 나타났다 . 6
월 16 일 정점 2 에서의 상대적으로 높은 수온과 높은 암모니아 농
도가 Leptocylindrus sp. 에 영향을 준 것으로 판단된다 . 정점 1, 4,
5 는 1 개의 과와 2 개의 판으로 구성된 각을 갖고 있으며 우측판 뒤
쪽 끝에 유입구를 형성한 와편모조류의 Prorocentrum sp. 이 우점 을 보였다 (Table 2). Prorocentrum 속은 진해만 ( 박 , 1980; 이 등 , 1981; 유와 허 , 1982; 한과 유 , 1983; 유 , 1984) 그리고 총무항 ( 심
등 , 1984), 여수 근해 ( 심 등 , 1981) 에서 많이 출현 되었다 . 6 월 23
일에는 중형식물플랑크톤의 기여율이 높은 정점 2 에서 은편모조 류의 Cryptomonas sp. 의 우점을 제외하고 기타 지역에서는 여전 히 Prorocentrum sp. 가 우점으로 출현하였다 . 와편모조류는 증식 과 에너지 유지를 위해 충분한 영양염류와 일조량의 공급이 요구 되는데 많은 종류의 와편모조류는 이를 위해 주간에는 저층으로 이동하여 충분한 영양염을 공급받는 수직일주운동을 한다 . 종에
따라서는 수직일주운동의 양상이 조금씩 다른데 Cochlodinium polykrikoides 의 경우 주간에는 대부분 3 m 이내의 표층에만 존재 하면서 세포들이 강한 띠 (patch) 를 형성하나 Alexandrium 속은 표 , 중
층에 주로 존재하고 강한 띠를 형성하지 않는다 ( 김과 주 , 2000).
Cryptomonas sp. 는 영양염을 소모하면서 광합성을 할 때 박테리
아로부터 질소와 인을 이용하여 영양염을 대체한다 (Jotaro et al. ,
2000). 6 월 26 일에 25~33 psu 의 염분 농도 범위와 20
oC 이상의
수온 (Fig. 3), 그리고 담수에 인한 영양염 유입으로 와편모조류의
Prorocentrum sp. 가 우점을 보이는 것으로 판단된다 . 6 월 16 일에
정점 3 에서 규조류의 Skeletonema costatum 이 우점하다가 6 월 23
일에는 와편모조류의 Prorocentrum sp. 가 우점으로 나타났는데 이 는 6 월 16 일의 낮은 인산염으로 하여 성장이 제한되었을 것이라 고 판단된다 . 일본 하리마해역에서 영양염 농도와 식물플랑크톤
Table 1. Seasonal composition (%) of phytoplankton group during the sampling period from the study sites of the Asan Bay
아산만 식물 플랑크톤 그룹별 분포 (%)
Month Bacillariophyceae Cryptophyceae Dinophyceae Euglenophyceae Chlorophyceae Cyanophyceae Other
Jun-16 39.00 9.89 45.05 0.58 0.19 2.02 3.27
Jun-23 22.08 28.51 42.08 2.71 0.90 1.00 2.71
Jul-13 59.20 8.43 17.74 1.11 2.88 5.54 5.10
Jul-24 71.77 22.03 4.13 1.06 0.06 0.00 0.95
Aug-11 90.43 5.57 0.32 0.00 0.00 0.00 3.68
Aug-25 89.49 5.57 1.24 0.00 0.00 0.00 3.71
Table 2. Dominant genera of phytoplankton community during the sampling periods at the 5 stations of the Asan Bay
식물 플랑크톤의 우점속 분포
Month Station Dominant sp. Percentage(%) Month Station Dominant sp. Percentage(%)
St. 1
Prorocentrumsp. 32.26 St. 1
Prorocentrumsp. 28.87
St. 2
Leptocylindrussp
.41.00 St. 2
Cryptomonassp. 37.07
June-16 St. 3 Skeletonema costatum. 46.00 June-23 St. 3
Prorocentrumsp. 35.00
St. 4
Prorocentrumsp. 53.00 St. 4
Prorocentrumsp. 45.00
St. 5
Prorocentrumsp. 57.00 St. 5
Prorocentrumsp. 43.00
St. 1
Nitzschiasp
.Leptocylindrus