Bacillus licheniformis GA9가 생산하는 키틴 분해효소의 정제 및 특성
황동호1, 홍성욱2, 황형서3, 정건섭1*
1연세대학교생명과학기술학부
2세계김치연구소
3세명대학교한방바이오융합과학부
Received: July 18, 2016 / Revised: October 19, 2016 / Accepted: November 19, 2016
서 론
생태계에서합성되는바이오매스에는키틴, 셀룰로오스, 단백질, 지질, 탄수화물, 리그닌등과같이매우다양한성분 들이 존재하는데 그 중에서 키틴은 N-acetylglucosamine (GlcNAc)이β-1,4 결합으로중합되어있는고분자물질로서 주로절족동물과곤충, 해양무척추동물등에의해서생합성 되고일부는진균의세포벽성분으로존재한다. 키틴은매년 약 10억톤정도생성되며, 분자구조는셀룰로오스와유사 하지만쉽게분해되지않는다[14].
키틴분해효소는 N-acetylglucosamine 중합체의 β-1,4 결 합을가수분해하는효소로 Bernard [2]에의해처음난초에 서키틴분해효소의존재가밝혀졌으며, 이후로키틴분해효 소가다양한생물종으로부터발견되어많은연구가진행되 었다. 최근에는사람에게도키틴분해효소가존재하는것으
로보고되어있다[5]. 최근에사용되고있는산업용세균유
래키틴분해효소들은주로 Bacillus 속미생물이생성하고 있으며 그 외에 Aeromonas, Arthrobacter, Clostridium, Enterobacter, Pseudomonas, Serratia, Streptomyces 등과 같은미생물들도키틴분해효소를생성하는것으로보고되 었다[7, 9].
키틴분해효소를활용한산업은매우다양하며, 고전적인 황산과염산을이용한산가수분해방법을사용해온의약품 및식품분야에키틴분해효소를적용하고있다[20]. 퇴행성 Purification and Characterization of a Chitinolytic Enzyme Produced by Bacillus licheniformis GA9
Dong Ho Hwang1, Sung Wook Hong2, Hyung seo Hwang3, and Kun Sub Chung1*
1Division of Biological Science and Technology, Yonsei University, Wonju 26493, Republic of Korea
2World Institute of Kimchi, Gwangju 61755, Republic of Korea
3School of Integrated Oriental Medical Bioscience, Semyung University, Jecheon 27136, Republic of Korea
A bacterium producing a large amount of chitinolytic enzyme was isolated from the intestinal tract of earthworm. The isolate was identified as Bacillus licheniformis by 16S ribosomal RNA analysis and desig- nated as B. licheniformis GA9. The enzyme was purified by 40−60% ammonium sulfate precipitation, diethyl-aminoethyl groups exchange chromatography, and gel filtration chromatography. The molecular weight was estimated to be 52.1 kDa and the N-terminal amino acid sequence was D-S-G-K-N-G-K-I-I-R-Y-Y- P-I-R. The optimum activity of the purified chitinolytic enzyme was shown at pH 5.0 and 40℃, and the enzyme was stable in the ranges of 20−50℃ and pH 5.0−6.0. Enzyme activity was increased by Co2+, while it was inhibited by Cu2+ and Fe2+. But it was recovered by chelating metals with ethylenediaminetetraacetic acid. The Km and Vmax values of the purified enzyme were 4.02 mg/ml and 0.52 mg/min, respectively. The chi- tinolytic enzyme characterized in this study has potential applications in areas such as biotechnology, bio- medicine, agriculture, and nutrition.
Keywords: Purification, chitinolytic enzyme, Bacillus licheniformis, GA9
*Corresponding author
Tel: +82-33-760-2252, Fax: +82-33-760-2183 E-mail: [email protected]
© 2016, The Korean Society for Microbiology and Biotechnology
관절염의통증완화에효과가있는글루코사민(glucosamine) 유사키틴단량체(GlcNAc) 생산에도이용하고있으며, 또한
진균의외벽을이루고있는키틴의 β-1,4 결합을분해함으로
서식물병원성진균의생장을억제하는친환경방제제로도
사용하고있다[18, 25]. 현재까지화학농약은식물병해를예
방함으로써식용및약용작물의수확량을증대시켰지만, 잔 류독성과농약으로인한환경오염및농업생태계파괴, 식 물병원성진균의농약에대한내성증가와같은부작용이 발생되었다[3]. 따라서최근친환경생물학적방제로서미생 물유래바이오제제에대한관심이높아지고있는추세이다 [11, 17].
본연구는키틴분해효소를산업적으로활용하기위해키 틴분해활성이우수한미생물을지렁이장내에서분리및선 발하였고, 선발미생물이생산하는키틴분해효소를정제하 고이화학적특성을확인하였다.
재료 및 방법
미생물 분리 및 배양
키틴분해활성이있는미생물의분리는지렁이농장(강원 도홍천)에서지렁이(Eisenia fetida)를구입하여각종우분, 도축부산물, 키틴, 제지슬러지, 톱밥등을먹이원으로공급 하면서 16주동안지렁이를사육하였다. Clean bench내에서 지렁이표면을 70% ethanol로소독하고화염멸균한 blade 로절개한후, 지렁이장내용물을취해 0.85% (w/v) NaCl 용액에현탁하였다. 현탁액 1 ml를 0.85% (w/v) NaCl 용액 9 ml에십진 희석하여 2% (w/v) colloidal chitin이포함된 screening 고체배지(0.2% colloidal chitin, 0.07% K2HPO4, 0.03% KH2PO4, 0.05% MgSO4· 5H2O, 0.001% FeSO4· 7H2O, 0.001% ZnSO4,0.01 g MnCl2, 0.2% yeast extract, 1.5% agar powder)에도말하였고 30℃에서 48시간동안배 양하였다. 배양후분해환(clear zone)을형성하는미생물군 집을순수분리하였다.
Colloidal chitin 제조
키틴(Sigma Co., USA) 40 g을 HCl 용액 400 ml에넣어 상온에서 30분간교반하였다. 염산에의해갈색으로점액질 용액이된키틴에증류수 2 L를첨가하여약 2시간동안교 반하였고 Whatman No.2 여과지(Whatman Intl. Ltd.,
England)를이용하여감압여과를수행하였다. 키틴수용액
의 pH가 6.0이될때까지감압여과를반복하였다. 최종생 성된 colloidal chitin은 121℃, 15분동안멸균하여 4℃에서 보관하였으며, sodium acetate buffer (pH 5.0)에 colloidal chitin을혼합하여기질로사용하였다.
효소활성 측정
분리미생물은최소배지인 DAVIS 액체배지(0.2% colloidal chitin, 0.07% K2HPO4, 0.03% KH2PO4, 0.05% MgSO4· 5H2O, 0.001% FeSO4· 7H2O, 0.001% ZnSO4, 0.01 g MnCl2, 0.2% yeast extract)에서 30℃, 72시간동안배양하였 으며배양액을원심분리(10,000 × g, 4℃, 10 min)한후, 상 등액을조효소액으로사용하였다.
키틴분해효소의활성측정은 Thamthiankul 등 [22]의방 법을 변형하여 1% (w/v) colloidal chitin (0.1 M sodium acetate buffer, pH 5.0)을기질로사용하여 DNS 환원당정 량법으로 수행하였다. 1% (w/v) colloidal chitin solution (0.1 M sodium acetate buffer, pH 5.0) 500 μl에조효소액 500 μl을첨가하여 37℃에서 2시간동안반응시킨후, 100℃ 에서 10분동안 열처리하여효소반응을중지시켰다. DNS solution (3,5-dinitrosalicylic acid) 1 ml을첨가하여 100℃ 에서 20분동안반응시키고원심분리(10,000 × g, 4℃, 10 min) 하여 상등액을 취한 후, spectrophotometer (VersaMaxTM ELISA microplate reader; Molecular Devices Co., USA)
를이용하여 550 nm에서흡광도를측정하였다. 효소활성
1 U (unit)는 1분동안에 1 μmole의 N-acetylglucosamine
(GlcNAc)을유리시키는데필요한효소의양으로나타내었다.
미생물의 동정
분리 미생물의 형태적 동정은 Bergey's Manual of
Systematic Bacteriology에따라분류하였다[19]. 분리미생 물의 생화학적 동정은 API 50 CHB kit (bioMerieux Co., France)로 49개탄소원에대한이용성을조사하였고결과는 API web database (http://apiweb.biomerieux.com)을이용 하여동정하였다.
분리미생물의유전학적동정은 16S ribosomal DNA 염기 서열분석을 통해수행하였다. 분리 미생물로부터분리한 chromosomal DNA를 universal primer인 27 forward primer (5’-AGAGTTTGATCATGGCTCAG-3’)와 1492 reverse primer (5’-GGATACCTTGTTACGACTT-3’)를사용하였다. PCR (MinicyclerTM; MJ research Inc., USA) 증폭은 순차 적으로 94℃에서 1분간 denaturation, 51℃에서 1분간 annealing, 72℃에서 1분 30초동안 extension을 30 cycles 실시하였고 72℃에서 5분간최종 extension을실시하였다. 증폭된 약 1.4 kb의 fragment를 T vector (Invitrogen, USA)에 결합시킨 후 형질전환하여 T vector sequencing primer를이용하여 Applied Biosystems 3730 DNA analyzer (Applied Biosystems, USA)로염기서열을분석하고 GenBank (NCBI, USA)의 database에등록된 16S rDNA 염기서열의 정보와 BLAST program (http://www.ncbi.nlm.nih.gov/
BLAST)을이용하여분리미생물을동정하였다.
효소의 분리 및 정제
분리미생물을 DAVIS 액체배지에 2% (v/v) 접종하여
30℃에서 72시간동안 배양한 후, 배양액을 원심분리
(10,000 × g, 4℃, 30 min)하여상등액에 ammonium sulfate (Junsei, Japan)를 0−20%, 20−40%, 40−60%, 60−80%, 80% 이상의농도로포화시키며분획별침전물을각각원심 분리(20,000 × g, 4℃, 25 min)하였다. 원심분리하여얻은침 전물을 10 mM Tris· HCl buffer (pH 7.2)에용해시킨후동 일한완충용액에 24시간동안투석(Spectra/Por 1, MWCO 6-8,000; Spectrum Laboratories Inc., USA)을수행하여잔 존하는염을제거하였다. 모든정제과정은효소단백질의변 성을 최소화하기 위하여 4℃에서 진행하였다. DEAE- sepharose fast flow으로 충진된 column (2.5 × 50 cm, Pharmacia Biotech., Sweden)을 10 mM Tris· HCl buffer (pH 7.2)로평형화시킨후, 투석한농축조효소액을주입하 여 1 ml/min의유속에서동일완충액으로흡착시켰다. 동일 완충액에용해시킨 linear gradient한 0−1 M NaCl로용출
하여 4 ml씩분획하였다. 각분획마다키틴분해활성을측
정하고단백질함량은 280 nm에서 흡광도를측정하였다.
Sephacryl S-300HR 으로 충진된 column (1.5 × 100 cm, Pharmacia Biotech.)을 10 mM Tris·HCl buffer (pH 7.2) 로평형화시키고효소액을 0.5 ml/min의유속으로용출하여
분획당 2 ml씩분획하였다. 각분획마다키틴분해활성을
측정하고단백질함량은 280 nm에서흡광도를측정하였다. 단백질 정량
키틴분해효소단백질의정량은 Lowry’s method에따라 수행하였다[15]. 정제한 각각의 효소액 50 μl에 biuret reagent (0.75 mmol/l cupric sulfate, 94 mmol/l sodium hydroxide) 550 μl를 넣어 25℃에 10분간 반응시킨 후, Folin-Ciocalteu's reagent (Sigma) 25 μl를넣고 30℃에서 30분동안반응하였다. Spectrophotometer를이용하여 725 nm 에서흡광도를측정하였으며, 표준단백질로 bovine serum albumin (BSA; Sigma)을사용하였다.
분자량 측정
키틴분해효소의분자량측정은 sodium dodecyl sulfate- polyacrylamide gel electrophoresis(SDS-PAGE)를사용하 였다. Sample buffer [12.5 mM Tris·HCl (pH 6.8), 2% β- mecaptoethanol, 20% glycerol, 0.02% bromophenol blue, 2% SDS]와효소액을 4:1로혼합하여 90℃에서 3분간열처 리한후, 80 V에서 2시간동안전기영동을수행하였다. 전기 영동이완료된 gel은 coomassie brilliant blue로 4시간동안
염색하고증류수로탈색하였다. 단백질분자량측정을위한 표준 단백질로 broad-range pre-stained marker (7−240 kDa; Elpis Biotech Inc., Korea)를사용하였다.
Zymography 분석
SDS-PAGE 전기영동한 10% (w/v) polyacrylamide gel을 1% Triton X-100 용액(10 mM Tris·HCl buffer, pH 7.5)과 멸균증류수를이용하여각각 15분동안세척하여 SDS를제 거한 후, 8% (w/v) colloidal 키틴을 함유한 10% (w/v) polyacrylamide gel 위에중첩시켰다. 중첩시킨두장의 gel 은 10 mM Tris· HCl buffer (pH 7.5)가 존재하는 square dish에보관하며 37℃에서 72시간동안반응시켰다.
N-terminal 아미노산 서열분석
정제한효소단백질을 SDS-PAGE 전기영동한후, transfer buffer [10 × transfer buffer (Tris 30.3 g, glycine 144.2 g, D.W 1 liter, pH 8.3) 100 ml, methanol 200 ml, D.W 700 ml]
에 평형화시킨 polyvinylidene difluoride (PVDF) membrane (Bio-Rad Laboratories, USA)에 17 V, 40분동안 transfer 하였다. Transfer한 후, PVDF membrane은 coomassie brilliant blue로염색하고 methanol로탈색하였다. PVDF
membrane을 건조하여 염색한 효소단백질을 확인한 후,
Edman 방법에 따라 Protein/Peptide sequencer (Applied Biosystems Inc.)를이용하여아미노산서열을확인하였다 [4].
최적반응 pH 및 pH 안정성
효소활성의최적 pH를측정하기위하여 0.1 M sodium acetate buffer (pH 4.0−6.0), 0.1 M sodium phosphate buffer (pH 6.0−8.0), 0.1 M Tris·HCl buffer (pH 8.0−10.0) 를키틴분해활성측정완충액으로사용하였다. 최적반응 pH 는조효소액과정제한효소액각각 50 μl에 10 mM sodium acetate buffer (pH 5.0) 450 μl를 첨가한 후, colloidal chitin을각 pH의 buffer (pH 4.0−10.0)에 1% 농도로녹여 기질로서사용하여측정하였다. 효소의 pH 안정성은조효소 액과정제한효소액각각 50 μl와 pH 4.0−10.0로조정한완 충액 450 μl를혼합하여 37℃에서 4시간동안방치한후잔 존효소활성을측정하였다.
최적반응 온도 및 온도 안정성
효소활성의최적반응온도는 20℃에서 80℃까지각온도 에서 조효소액과 정제한 효소액을 2시간동안 1% (w/v) colloidal chitin (0.1 M sodium acetate buffer, pH 5.0) 기 질과반응시켜측정하였고, 온도안정성은조효소액과정제 한효소액을최적온도조건과동일하게 4시간동안반응시킨
후, 잔존효소활성을측정하였다. 금속이온의 영향
키틴분해효소의활성에영향을미치는금속이온을조사 하기 위하여 CaCl2, CoCl2, CuCl2, FeCl2, MgCl2, MnCl2, ZnCl2를사용하여측정하였다. 정제한효소 50 μl와금속이 온 450 μl를혼합하여금속이온의최종농도가각각 2 mM 과 10 mM이되도록하여 37℃에서 30분간반응시켜잔존 효소활성을측정하였다.
Km및 Vmax 측정
키틴분해효소의 Km값과 Vmax값은기질농도([S])에따른 반응속도([V])를측정하고측정값의역수를취해 Lineweaver- Burk plot을작성하여 결정하였다. 기질은 0.1 M sodium acetate buffer (pH 5.0)에 0.1−0.4% (w/v) colloidal chitin 농도로 37℃에서 60분간반응시켜기질농도별효소활성을 측정하였다.
결과 및 고찰
키틴 분해효소 생산미생물 분리 및 선발
지렁이의장내용물을 0.85% (w/v) NaCl 용액에십진희 석하여 2% (w/v) colloidal chitin을함유한 고체배지에도 말·배양하여콜로니주위로투명하게분해환 (clear zone)을 생성하는미생물총 1,520주를 1차선발하였다. 1차선발미 생물은 2% (w/v) colloidal chitin을함유한액체배지에배양 한후, 조효소액을이용하여 키틴분해활성이우수한균주 를 2차선발하였으며, 키틴분해효소를정제하기위한영양
최소배지인 DAVIS broth에접종하여키틴분해활성이가장 우수한 GA9 균주를최종선발하였다(data not shown).
선발미생물의 동정
선발미생물을 Gram 염색과아포염색을하여현미경으로
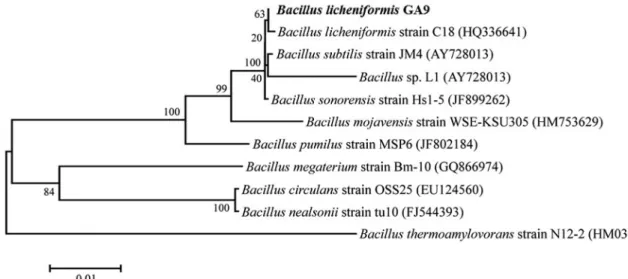
관찰한결과, 그람양성간균으로관찰되었고내생포자를생 성하는것으로확인되었다(data not shown). 생화학적동정 을 위하여 API 50 CHB kit test를 실시한 결과, Bacillus licheniformis와 99.9% 상동성을 나타내었다(data not shown). 유전학적동정을위하여 16S ribosomal RNA 유전 자염기서열을 CLUSTAL X로 Multiple sequence alignment 한후, MEGA 4.0을사용하여 Phylogenetic tree 분석한결 과, Bacillus licheniformis와가장가까운유연관계를보였 다. 따라서, 형태적특징, 생화학적특성과 16S rRNA 염기서 열분석결과를종합하여, 선발미생물을 Bacillus licheniformis GA9으로명명하였다(Fig. 1).
키틴 분해효소의 분리 및 정제
키틴 분해효소를 정제하기 위하여 2% (w/v) colloidal chitin이함유된 DAVIS 액체배지에서 30℃, 48시간동안배 양한 B. licheniformis GA9의배양상등액에 황산암모늄을 0−20%, 20−40%, 40−60%, 60−80% 농도로 포화시켰으며, 생성된침전물을각각키틴분해활성을비교한결과, 40−60%
황산암모늄침전물에서키틴분해효소의활성이존재함을 확인하였다(data not shown).
황산암모늄활성분획을 10 mM Tris· HCl buffer (pH 7.5) 에용해시킨후, 동일한완충용액으로투석하여황산암모늄 을제거한 시료를음이온교환수지 DEAE-sepharose fast
Fig. 1. Phylogenetic tree of isolate GA9 through 16S ribosomal DNA sequence homology.
flow에흡착시키고 0−1 M NaCl 용액을 linear gradient하 게용출시켜서분획마다키틴분해활성을측정한결과, 총 240개의분획중 149−186번분획에서키틴분해활성을확 인하였으며, 활성분획은투석을실시한후, 동결건조하여농 축하였다. 농축한키틴분해활성분획을겔크로마토그래피 로정제한결과, 총 160개의분획중키틴분해활성을나타 낸 55−71번분획을모아동결건조하여농축하였다(data not
shown). 키틴분해효소를최종정제한결과, 배양상등액에
비해 45.2배농축되었으며 회수율은 20.0%로확인하였다
(Table 1).
키틴 분해효소의 분자량 측정과 Zymography
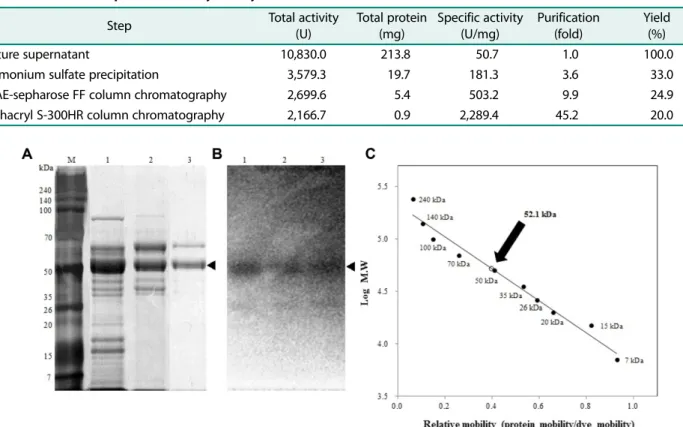
정제한키틴분해효소의분자량을측정하고단일성을확 인하기위하여 SDS-polyacrylamide gel과 zymography 전 기영동을수행하였다. SDS-PAGE와 zymography를비교하 여키틴분해활성이나타난위치의 band를확인한결과는 Fig. 2A와 Fig. 2B에각각나타내었으며, 황산암모늄분획, 이온교환크로마토그래피와겔크로마토그래피모두동일한
위치에서분해환을확인하였다. 정제한키틴분해효소단백 질의분자량은 52.1 kDa으로추정되었다(Fig. 2C).
키틴 분해효소의 N-terminal 아미노산 서열분석
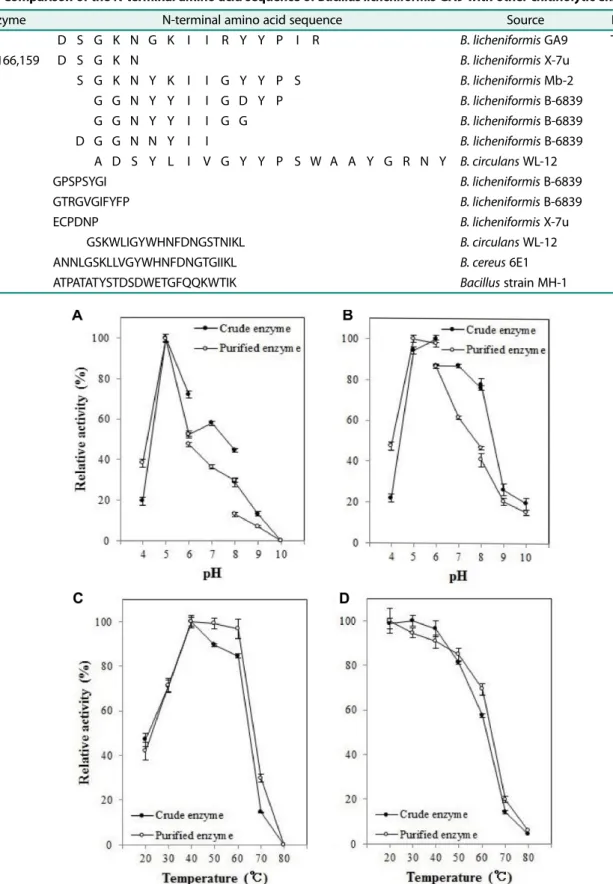
정제한키틴분해효소단백질의 N-terminal 아미노산서 열은 Table 2에 나타내었는데 Asp-Ser-Gly-Lys-Asn-Gly- Lys-Ile-Ile-Arg-Tyr-Tyr-Pro-Ile-Arg로분석되었다. Bacillus 속(이) 생산하는키틴분해효소의 N-terminal 아미노산서 열과비교한결과, B. licheniformis X-7u가생산하는키틴 분해효소인 Chi176, 166, 159와일부 아미노산서열(Asp- Ser-Gly-Lys-Asn)이일치하였고[21], B. licheniformis Mb-2 가생산하는 Chi67의 N-terminal 아미노산서열(Ser-Gly- Lys-Asn-Tyr-Lys-Ile-Ile-Gly-Tyr-Tyr-Pro-Ser)과 B. circulans
WL-12가생산하는키틴분해효소 ChiA1 역시부분적으로
일치(Ala-Asp-Ser-Tyr-Leu-Ile-Val-Gly-Tyr-Tyr-Pro-Ser-Trp- Ala-Ala-Gly-Arg-Asn-Tyr)하는것을확인하였다[23, 27]. 그 러나그외에 Bacillus 속(이) 생성하는키틴분해효소의아 미노산서열은일치하지않는것으로확인되었다(Table 2).
Table 1. Purification step of the chitinolytic enzyme from Bacillus licheniformis GA9.
Step Total activity
(U)
Total protein (mg)
Specific activity (U/mg)
Purification (fold)
Yield (%)
Culture supernatant 10,830.0 213.8 50.7 1.0 100.0
Ammonium sulfate precipitation 3,579.3 19.7 181.3 3.6 33.0
DEAE-sepharose FF column chromatography 2,699.6 5.4 503.2 9.9 24.9
Sephacryl S-300HR column chromatography 2,166.7 0.9 2,289.4 45.2 20.0
Fig. 2. Protein pattern and determination of molecular weight of the purified enzyme by SDS-PAGE. (A) SDS-PAGE, (B) zymog- raphy, (C) molecular weight. Lane M, protein molecular weight markers; lane 1, proteins precipitated by 40−60% ammonium sulfate;
lane 2, proteins eluted by DEAE-sepharose FF ion exchange chromatography; lane 3, proteins eluted by sephacryl S-300HR gel filtration.
Molecular weight marker proteins: 7 kDa, aprotinin; 15 kDa, lysozyme; 20−50 kDa; recombinant protein; 70 kDa, bovine serum albumin;
and 100-240 kDa; recombinant protein.
Table 2. Comparison of the N-terminal amino acid sequence of Bacillus licheniformis GA9 with other chitinolytic enzymes.
Enzyme N-terminal amino acid sequence Source Reference
D S G K N G K I I R Y Y P I R B. licheniformis GA9 This study
Chi176,166,159 D S G K N B. licheniformis X-7u [21]
Chi67 S G K N Y K I I G Y Y P S B. licheniformis Mb-2 [23]
Chi153 G G N Y Y I I G D Y P B. licheniformis B-6839 [24]
Chi143 G G N Y Y I I G G B. licheniformis B-6839 [24]
Chi166 D G G N N Y I I B. licheniformis B-6839 [24]
ChiA1 A D S Y L I V G Y Y P S W A A Y G R N Y B. circulans WL-12 [27]
Chi162 GPSPSYGI B. licheniformis B-6839 [24]
Chi149 GTRGVGIFYFP B. licheniformis B-6839 [24]
Chi189 ECPDNP B. licheniformis X-7u [21]
ChiD GSKWLIGYWHNFDNGSTNIKL B. circulans WL-12 [27]
Chi36 ANNLGSKLLVGYWHNFDNGTGIIKL B. cereus 6E1 [26]
ChiL ATPATATYSTDSDWETGFQQKWTIK Bacillus strain MH-1 [16]
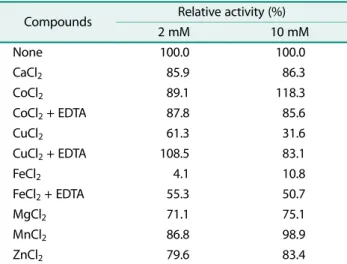
Fig. 3. Effect of pH and temperature on the activity and stability of the chitinolitic enzyme from Bacillus licheniformis GA9.
(A) optimum pH, (B) pH stability, (C) optimum temperature, (D) temperature stability.
최적반응 pH 및 pH 안정성 측정
조효소액과정제한키틴분해효소의최적반응 pH와 pH 안정성결과는 Fig. 3A와 Fig. 3B와같다. 조효소액과정제 한효소액모두 pH 5.0에서최대활성을나타내었으며(Fig.
3A), 조효소액은 pH 5.0−8.0 사이에서안정하였고정제한효 소액의경우 pH 5.0−6.0에서안정하였다(Fig. 3B).
조효소액과정제한효소액의키틴분해활성은알칼리조 건에서보다산성조건에서비교적높게나타내었다. Wang 등[25]이분리한 Bacillus amyloliquefaciens V656이생산한 두개의항진균성키틴분해효소 F1와 F2는각각 pH 7.0과
pH 6.0에서 최대 활성을 나타낸다고 보고하였으며, B.
licheniformis Mb-2가생산한키틴분해효소는 pH 6.0에서 최대활성을나타내었고 pH 4.0−11.0 범위에서효소활성이 70%이상유지되었다고보고하였다[23]. B. cereus CH가생 산하는키틴분해효소A, B1, B2, B3는 pH 5.0−7.5에서최대 활성을나타내었고[13], B. circulans No.41가생산하는키 틴분해효소는 pH 8.0일때최대활성을나타내었으며 pH 3.0과 pH 12.0에서는약 50%의키틴분해활성이소실된다 고보고하였다[28]. 대부분의 Bacillus 속(이) 생산하는키틴 분해효소들은 pH 5.0−8.0 사이에서최대활성을나타내는것 으로확인되었다.
최적반응 온도 및 온도 안정성 측정
조효소액과정제한키틴분해효소의최적온도와온도안 정성에대한측정결과는 Fig. 3C와 Fig. 3D에나타내었다. 조효소액과정제한효소액에대한온도의영향은 20℃부터 효소활성이점차증가하여 40℃에서최대활성을나타내었다 (Fig. 3C). 조효소액은 20℃에서 50℃사이에서온도에대한 안정성을나타내었으나 60℃이상부터는효소활성이급격히 감소하여 70℃이상에서는활성이소실되었다. 정제한효소 액은 60℃까지약 70% 이상효소활성이유지되었고 70℃이 상부터는효소활성이급격히감소하여소실되었다(Fig. 3D).
Wang 등 [25]이분리한 B. amyloliquefaciens V656이생 산한두개의항진균성키틴분해효소 F1와 F2는 40℃에서 최대활성을나타내었다고보고하였으며, B. licheniformis Mb-2가생산한키틴분해효소는 70℃에서최대활성을나타 내었으며 75℃이상부터는효소활성이감소하였다고보고하 였다[23]. B. licheniformis SK-1이생산하는키틴분해효소 는 55℃에서최대활성을나타내었고 50℃까지는매우안정 하며 60℃에서의효소활성은 90분마다 50%씩감소하였다고 보고하였다[10]. Bacillus 속(이) 생산하는키틴분해효소는 대부분 40−50℃사이에서최대활성을나타내었으며 60−70℃ 까지 온도안정성을 나타내었다. 따라서 B. licheniformis GA9이생산한키틴분해효소는기존의연구결과들과유사 한결과를나타내었다.
금속이온에 대한 영향
정제한키틴분해효소의금속이온에대한영향을조사하 기위하여각각의금속이온을최종농도가 2 mM과 10 mM
이되도록조정하여측정한결과는 Table 3에나타내었다.
10 mM Co2+금속이온존재시효소활성은 118.3%로증가하 였으며, 2 mM Fe2+과 Cu2+금속이온존재시효소활성은각 각 4.1%와 61.3%로감소하였고 10 mM의경우는각각 10%
와 31.6%로감소하였다.
B. licheniformis B-6839가 생산하는 키틴 분해효소는 1 mM의 Mn2+, Zn2+ 그리고 Co2+에의해키틴분해활성이약 70% 이하로감소하였으며 Mg2+와 Ca2+에의해서는효소활 성이 80−90% 감소를나타내었다[24]. 또한 Gomaa [6]에따 르면 B. licheniformis NM120-17이생성하는키틴분해효소 는 5 mM Cu2+이존재할때잔존활성이 98.9%로감소하였 으며금속이온농도가 10 mM로증가하였을때, 85.4%로더 감소하였다고 보고하였다. 또한, Fe2+ 존재시 84.7%에서
77.1%로금속이온의농도가증가할수록잔존효소활성이감
소한다고보고하였다. 정제한키틴분해효소활성에영향을 미치는금속이온과 EDTA를혼합하였을때, 효소활성이크 게감소되었던 Fe2+와 Cu2+는 EDTA 존재시효소활성이회 복되었으며, Co2+금속이온은 EDTA가존재할때, 증가되었 던효소활성이감소하는결과를나타내었다.
Km및 Vmax 측정
정제한키틴분해효소의기질친화도를측정하기위해다 양한농도의기질(0.1−0.4% (w/v) colloidal chitin)을이용해 Lineweaver-Burk 방법으로 plot하여기질별로 Km과 Vmax
Table 3. Effect of metal ion on the chitinolytic enzyme from Bacillus licheniformis GA9.
Compounds Relative activity (%)
2 mM 10 mM
None 100.0 100.0
CaCl2 85.9 86.3
CoCl2 89.1 118.3
CoCl2 + EDTA 87.8 85.6
CuCl2 61.3 31.6
CuCl2 + EDTA 108.5 83.1
FeCl2 4.1 10.8
FeCl2 + EDTA 55.3 50.7
MgCl2 71.1 75.1
MnCl2 86.8 98.9
ZnCl2 79.6 83.4
The enzyme was pre-incubated at various metal ions and EDTA in 10 mM sodium acetate buffer (pH 5.0) for 30 min at 37℃ and remaining activity was measured.