K. B. Lee ( ) 조선대학교 생물학교육과
(Department of Biological Science Education, Chosun University) e-mail: [email protected]
식물의 생장과 발달에 영향을 미치는 슈트 정단분열조직의 체제와 기능
이규배
Organization and function of shoot apical meristem affecting growth and development in plants
Kyu Bae Lee
Received: 14 November 2014 / Revised: 9 December 2014 / Accepted: 20 December 2014
ⓒKorean Society for Plant Biotechnology
Abstract In plants, a shoot apex has a small region known
as the shoot apical meristem (SAM) having a group of dividing (initiating) cells. The SAM gives rise to all the groundabove structures of plants throughout their lifetime, and thus it plays important role in growth and development of plants.
This review describes theories to explain the SAM organization and function developed over the last 250 years. Since in 1759 German botanist C. F. Wolff has described firstly the SAM, in 1858 Swiss botanist C. Nägeli proposed the apical cell theory from the observation of a large single apical cell in the SAM of seedless vascular plants: however, this view was recognized to be unsuitable to seed plants. In 1868, German botanist J. Hanstein suggested the histogen theory: this concept subdividing the SAM into dermatogen, periblem, and plerome was unable to generally apply to seed plants. In 1924, German botanist A. Schmidt proposed the tunica-corpus theory from the examination of angiosperm SAM in which two parts show different planes of cell division: this theory was proved to be not suitable to gymnosperm SAM, not have stable surface tunica layer. In 1938, American botanist A. Foster described zones in gymnosperm SAM based on the cytohistologic differentiation and thus called it a cytohistological zonation theory. With works by E. Gifford, in 1954, this zonation pattern was demonstrated to be also applicable to angiosperm SAM.
As another theory, in 1952 French botanist R. Buvat proposed the méristème d’attente (waiting meristem) theory: however, this concept was confuted because of its negation of function
during vegetative growth phase to central initial cells. Rescent studies with Arabidopsis thaliana have found that formation and maintenance of the SAM are under the control of selected genes: SHOOTMERISTEMLESS (STM) gene forms the SAM, and WUSCHEL (WUS) and CLAVATA (CLV) genes function in maintaining the SAM; signaling between WUS and CLV genes act through a negative feedback loop.
Keywords Apical cell, Arabidopsis, Cytohistological zonation,
Histogen, Méristème d’attente (waiting meristem), Shoot apical meristem, Tunica-corpus
서 론
일반적으로 유관속식물은 접합자(수정난)가 분열하기 시 작하면서, 죽을 때까지 새로운 세포들이 만들어지고 새 로운 기관이 발달한다. 배발생의 초기에는 세포의 분열이 개체(胚) 전반에 걸쳐 일어나지만, 배가 독립생활을 하게 될 무렵에는 새로운 세포들이 점차 일정한 부위에 제한 된다. 이와 같이 분열하는 세포들이 모여 있는 부위를
‘분열조직(分裂組織, meristem)’이라고 한다(Evert 2006). 분 열조직이라는 용어는 스위스의 식물학자인 네겔리(Carl Nägeli)가 1858년에 발표한 그의 저서 “식물학 기고(Beiträge zur Wissenschaftlichen Botanik)”에서 처음으로 사용하였으 며(Nägeli 1858), 이 용어의 그리스어원은 ‘Gk. meristos’로 서 ‘분열하다(divide)’라는 뜻이다. 그래서 분열조직이라 는 용어는 기능적으로 ‘모든 조직들을 만드는 세포들’이 라는 뜻으로 쓰인다(Lyndon 1998; Tooke and Battey 2003).
분열조직에서 계속적으로 새로운 세포들을 만들어 낼 수 있는 원천으로서, 분열조직을 유지시킬 수 있는 세포를
DOI:http://dx.doi.org/10.5010/JPB.2014.41.4.180Review
ISSN 1598-6365
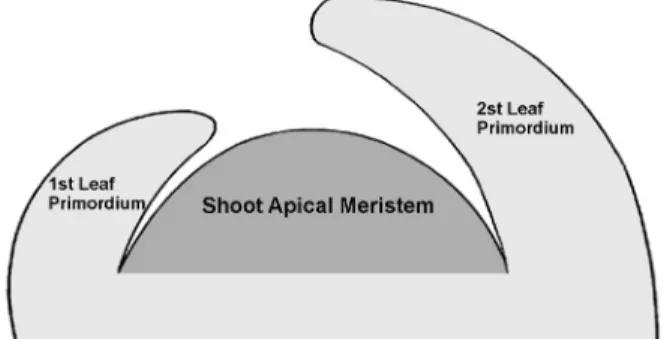
Fig. 1 Diagram of the shoot apical meristem (SAM) in plant.
The entire figure is the shoot apex consisting of leaf promordia and SAM, whereas the SAM is the dome-shaped, gray area, restricted at the tip of the shoot apex.
‘시원세포(始原細胞, initiating cells 또는 initials)’라고 한 다(Evert 2006). 고등동물에서 사용하는 ‘줄기세포(stem cell)’
라는 용어는 식물의 분열조직에 있는 시원세포에 해당하 며, 식물학자들도 시원세포를 줄기세포라고 하는 경우가 있다. 그러나 여기에서는 오래 전부터 쓰여 왔던 ‘시원세 포’라는 용어를 사용한다.
식물의 분열조직에서는 상반되는 두 가지 과정이 일어 난다. 즉, 시원세포들의 집단을 유지시키기 위한 시원세 포 자신의 재생(自己再生, self-renewal) 그리고 분열조직 에서 생긴 세포들을 그 주변에서 발달하는 기관으로 보 내는 일이다. 즉, 분열조직으로터 생긴 세포들이 분화하 여 발달하는 기관 형성에 쓰이도록 계속해서 기관 발생 장소로 이동함에도 불구하고, 분열조직에서 시원세포들 의 수는 놀라울만큼 일정하게 유지된다(Lyndon 1998; Laufs et al. 1998; Carles and Fletcher 2003). 이런 과정을 통해서, 새로 형성되는 시원세포의 수와 이로부터 엽원기 등으로 분화되어 사라지는 시원세포의 수가 거의 완벽하게 균형 을 이루게 된다(Laufs et al. 1998; Haecker and Laux 2001).
슈트(shoot) 정단분열조직을 유지시키고 기능하기 위해 서는 두 가지 주요 홀몬인 사이토키닌(cytokinin)과 옥신 (auxin)이 함께 작용하는 것으로 알려져 있다(Barton 2010).
사이토키닌은 정단분열조직을 유지시키고 세포분열의 특 성을 조절하는 데 중요한 역할을 하며, 정단분열조직에서 옥신이 축적되는 위치에서 잎과 같은 기관 발생이 이루 어진다(Haecker and Laux 2001; Vernoux et al. 2010; Murray et al. 2012). ‘슈트(shoot)’라는 용어를 ‘지상부’라고 하는 것이 그 정의에 가장 근접한 표기이지만, ‘땅속줄기(지하 경, rhizome)’를 갖는 식물에는 적용할 수 없다. 따라서
‘shoot’에 해당하는 정확한 한글 용어는 없으므로, 이를 외래어로 표기(‘슈트’)하는 것이 합리적이다.
식물의 슈트 정단분열조직으로부터 줄기, 잎, 꽃, 곁눈, 가시, 덩굴손 등과 같은 여러 가지 구조들이 형성된다.
따라서 슈트 정단분열조직은 그 구조들의 생장과 발달에 도 중요한 영향을 미친다. 슈트 정단분열조직은 크기가
작은 세포들이 모여 있는 조직으로서, Figure 1과 같이, 보통 반구형 또는 돔(dome) 모양이며 800 ~ 1,000여 개의 세포로 구성되어 있다(Medford 1992).
슈트 정단분열조직의 크기와 모양은 종에 따라서 그리고 발달 시기에 따라 다르다. 진정쌍자엽식물인 담배(Niconiana
tabacum)에서 슈트 정단분열조직의 폭은 약 100 μm이며 비교적 평평하여 높이가 낮다(Poething and Sussex 1985). 또 한 애기장대(Arabidopsis thaliana)에서 슈트 정단분열조직 의 크기(높이 x 폭)는 40 x 60 μm 정도로 비교적 평평하 며 높이가 낮은 모양이다(Barton 2010; Vernoux et al. 2010).
그러나 단자엽식물인 옥수수(Zea mays)에서 슈트 정단분 열조직의 크기(높이 x 폭)는 200 x 200 μm 정도로서(Barton 2010) 진정쌍자엽식물에 비하여 상당히 높게 솟아 있다 (Kessler et al. 2006; Brooks et al. 2009; Takacs et al. 2010).
식물의 슈트와 뿌리를 연결하는 주축의 양쪽 끝에 위 치하는 분열조직을 ‘정단(頂端)분열조직(apical meristem)’
이라고 한다. 식물은 배발생 이후의 거의 모든 생장이 슈 트의 맨 끝 부분에 있는 ‘슈트 정단분열조직(shoot apical meristem)’과 뿌리의 끝 부분에 있는 ‘근단분열조직(root apical meristem)’에서 일어난다. 그래서 일반적으로 이 두 가지의 정단분열조직으로부터 각각 식물의 지상부 및 지 하부의 구조가 형성된다. 이러한 정단분열조직은 식물의 발달을 위해서 절대적으로 필요하며, 지난 100여 년 동안 많은 연구의 주제가 되어 왔다(Dodsworth 2009; Barton 2010).
식물에서 슈트 정단분열조직은 배가 발생하는 동안에 형성되며, 그 후 생장하는 과정에서 슈트 정단분열조직 으로부터 줄기와 이의 부속구조(appendage)인 잎, 꽃, 꽃 차례, 곁눈, 가지, 가시 등이 발달한다. 이런 구조들은 최 초에 형성되었던 슈트 정단분열조직(끝눈)의 아래 쪽에 남게 되며, 이런 과정이 계속되어 슈트가 생장한다. 이와 같은 생장은 슈트 정단분열조직이 식물의 일생 동안 지 속적으로 유지되어야 정상적으로 이루어진다. 따라서 슈 트 정단분열조직은 부속 구조들의 생장과 발달에도 중요 한 영향을 미치므로, 슈트의 발달과 생장을 이해하기 위 해서는 우선적으로 슈트 정단분열조직의 체제를 파악해 야 한다.
지금까지 서구의 식물학자들에 의해서 제안된 슈트 정 단분열조직의 체제에 관한 이론들로서, 정단세포설(apical cell theory) (Nägeli 1858), 조직원설(histogen theory) (Hanstein 1868), 초층-내체설(tunica-corpus theory) (Schmidt 1924), 세 포조직학적 구역화설(cytohistological zonation theory) (Foster 1938; Gifford 1954), 대기(待機)분열조직설[méristème d’attente (waiting meristem) theory] (Bubat 1952) 등이 제안된 바 있 다. 이 설들은 이어지는 각론에서 자세히 논의된다.
또한 식물의 슈트 정단분열조직의 생장과 발달을 이해
하기 위한 모델 식물로서, 애기장대(Arabidopsis thaliana)
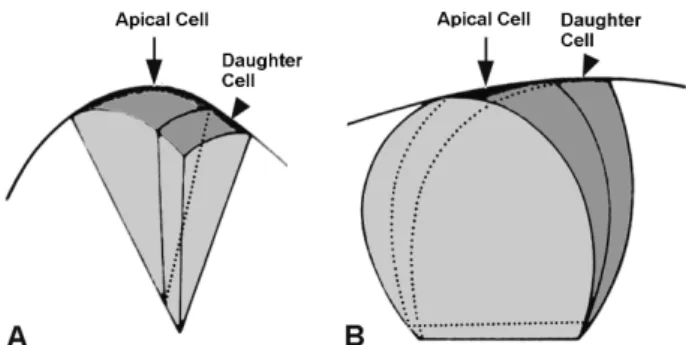
Fig. 2 Diagram of apical cells in a pteridophyte shoot apex. Two forms of apical cell (arrows): 4-sided or pyramidal shape (A) and 3-sided or lenticular shape (B). In (A) and (B), a daughter cell (arrowhead) or derivative is attached to the right side of the apical cell. Daughter cells produed by the subsequent cell division would be added to the both sides of the apical cell.
에서 슈트 정단분열조직의 체제가 밝혀진 이후(Vaughan 1952), 1990년대부터 이루어진 애기장대의 슈트 정단분열 조직을 형성하고 유지하는 작용을 조절하는 유전자의 분 자생물학적 연구를 소개할 것이다. 한편 국내에서는 저 자가 식물형태학 교재의 분열조직 단원에서 슈트 정단분 열조직의 체제에 관한 모형들을 소개한 바 있으나(Lee 2012), 그 이외에 이 주제를 대상으로 한 전문적인 연구 는 이루어진 바 없다.
이 연구의 목적은, (i) 식물학자들에 의해 제안된 슈트 정단분열조직의 체제에 관한 설(모형)들의 발전 과정을 기술하여, (ii) 식물의 생장과 발달에서 슈트 정단분열조 직의 기능에 관한 중요성을 이해하는 데 있다. 이러한 연 구를 통하여 식물의 해부학적 연구가 생리학, 발생학, 유 전학, 분자생물학 등 다른 분야의 발전에 기여할 수 있음 을 생각해 보고, 또한 식물학의 여러 분야에서 이루어지 는 연구에 도움이 되기를 기대한다.
정단세포설
1759년 독일의 식물학자 볼프(Kaspar Friedrich Wolff)는 후 성설(後成說, theory of epigenesis)을 지지하는 그의 박사 학위 논문 “Theoria Generationis”에 식물의 슈트 정단부에 서 조직과 잎들이 생기는 것을 최초로 기재하였다(Wolff 1759). 즉, 그는 ‘punctum vegetaionis’라고 지칭한 식물의
‘생장점(生長點)’을 발견하였다(Tooke and Battey 2003).
그 후 19세기에 들어서 광학 기구가 개량되어 식물을 더 자세하게 관찰하게 되었다. 1858년 스위스의 식물학자 네 겔리(Karl Nägeli)는 유관속식물(무종자식물)들의 슈트 정 단분열조직을 관찰한 결과, 형태적으로 커다란 하나의 정단세포(頂端細胞) (single apical cell)가 있다는 것을 발 견하여 “정단세포설(apical cell theory)”을 제안하였다[Nägeli 1858, Evert (2006)에서 인용하였음].
이 설에 의하면, 슈트 정단부에서 맨 끝의 중앙에 있는 커다란 하나의 정단세포는 4면이 있는 피라미드 모양 (Fig. 2A) 또는 3면이 있는 렌즈 모양(Fig. 2B)이다. 이런 세포가 분열하여 식물체를 구성하는 모든 세포들이 유래 된다(Bierhorst 1971; 1977). 즉, 정단세포가 식물의 전체 생장 과정을 조절하는 정단분열조직의 구조적 및 기능적 인 단위로 해석되었다.
예로서, 쇠뜨기속(Equisetum) 식물의 슈트 정단부의 중 앙에 하나의 커다란 정단세포가 있고, 이로부터 분열하 여 생긴 딸세포들이 납작한 벽돌을 쌓아 놓은 것처럼 정 단세포의 좌우 양쪽에 대칭으로 배열한다. 또한 줄고사 리속(Nephrolepis) 식물에서도 슈트 정단부의 한 가운데에 있는 정단세포가 표면에 대하여 수직으로 분열(수직분열) 하여 딸세포를 형성하며, 이어서 그 딸세포는 표면에 대 하여 수평으로 분열(수평분열)하여 세포수를 증가시킨다.
양치식물의 정단세포는 핵내재복제(endoreduplication)로 인하여 다배체의 빈도가 매우 높아서 정단세포의 유사분 열이 활발하게 일어나지 않을 것이다(Evert 2006). 그러나 슈트와 뿌리의 생장이 활발하게 일어나는 동안에는 정단 세포가 활발하게 분열한다(Gifford et al. 1979; Kurth 1981).
이 분야를 계속 연구한 학자들은, 양치식물 등의 하등 유 관속식물에는 하나의 정단세포가 존재하지만, 고등한 유 관속식물들에는 일반적으로 존재하지 않는다는 것을 증 명하였다. 따라서 정단세포설은 나자식물과 피자식물에 적용될 수 없게 되었으므로, 이 설은 식물체를 구성하는 서로 다른 부분들이 독립적으로 기원된다는 생각으로 바 뀌게 되었다.
조직원설
1868년에 독일의 식물학자 한스타인(Johannes Hanstein)은
“조직원설(組織源說, histogen theory)”을 제안하였다[Hanstein 1868, Evert (2006)에서 인용하였음]. 이 설에 따르면, 슈트 와 뿌리를 구성하는 세 가지의 1기조직들(표피조직, 기본 조직, 유관속조직)이 정단부에서 뚜렷하게 구별되는 원 표피(dermatogen), 원피층(periblem), 원중심주(plerome) 등의 세 가지 시원세포(들)로부터 각각 기원된다(Fig. 3). 3층으 로 배열되어 있는 시원세포(들)는 하나 또는 여러 개로 이루어져 있다(Cutter 1980). 조직원설에서 조직원으로부 터 분열되어 나온 딸세포들이 특별한 조직으로 분화되도 록 이미 결정되어 있다는 주장은 실제로 슈트 정단부에 적용되지 않는다(Foster 1939).
많은 동물의 배발생 과정에 있는 세포들의 운명처럼,
이 설은 식물에서 한 세포의 발생 운명도 그 세포의 계열
에 의해 조절된다고 생각한 것이다. 그러나 어떤 조직으
로 분화될 조직원으로 예정되어 있는 슈트 정단분열조직
Fig. 3 Diagram of the histogen organization of an angiosperm shoot apical meristem (SAM). The SAM has three initial layers:
dermatogem (d) at the upper surface layer, periblem (pb) at the middle layer, plerome at the lower layer. According to this concept, the epidermal, fundamental, and vascular tissues in a shoot are originated from the three initial layers, respectively. Arrows indicate the direction of cell division. LP1 and LP2, the first and second leaf primordia.
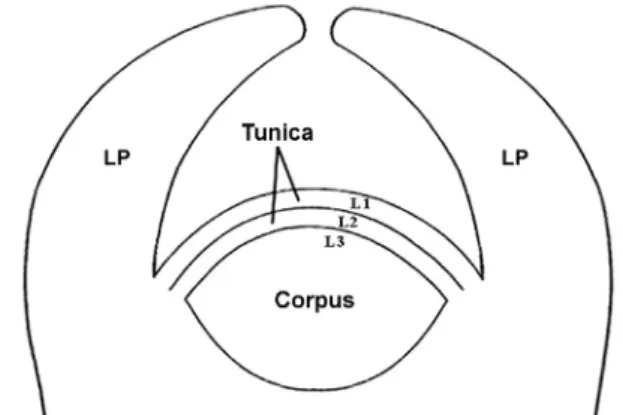
Fig. 4 Diagram of the tunica-corpus model of an angiosperm shoot apical meristem (SAM). The SAM is consisted of tunica (two layers) and corpus. Cells in the tunica layers would divide anticlinally, whereas cells in the corpus divide in all planes. The two tunica and corpus layers are represented by L1, L2, and L3, respectively. LP, leaf primordium.
부위를 외과적으로 잘라내어도 돔 모양의 슈트 정단분열 조직이 정상적으로 발달한다(Pilkington 1929; Snow and Snow 1951; Sussex 1952, 1964). 레이저를 이용하여 토마토의 슈 트 정단분열조직에서 중앙부(중앙구역)을 제거하더라도 며칠 후에 정단분열조직의 기능이 회복된다(Reinhardt et al. 2003). 이러한 결과들은 세포의 운명이 그 세포의 계 열에 의해서 결정되지 않음을 뜻한다. 예로서, 피층세포 는 분열조직 세포로부터 분화한다. 그 이유는 그 분열조 직 세포가 특이하기 때문이 아니라, 발생 과정에서 피층 세포의 대사작용을 변화시키도록 하는 신호를 받기 때문 이다(Moore et al. 1998). 따라서 분열조직을 원표피, 원피 층, 원중심주 등으로 나누는 것은 식물 전체에 일반화하 여 적용할 수 없다.
조직원설의 결정적인 오류는 식물체를 구성하는 서로 다른 부분들이 운명적으로 미리 결정된 3층의 시원세포 들로부터 독립적으로 분화되어 기원하리하고 추정한 것 이다(Tooke and Battey 2003; Evert 2006). 이 설은 슈트의 정단 분열조직 체제를 기재하기에는 적당하지 않으나, 뿌리의 분열조직을 기재하는데는 어느 정도 계속 사용되 고 있다(Evert 2006; Steeves 2006). 식물학자들은 슈트 정 단분열조직의 체체가 어떻게 이루어져 있는지를 설명하 기 위해서 새로운 설을 제안하게 되었다.
초층-내체설
정단분열조직의 구조에 관한 세 번째 설은, 1924년 슈미 트(A. Schmidt)가 피자식물의 슈트 정단부를 관찰하여 제 안한 “초층-내체설(鞘層-內體說, tunica-corpus theory)”이
다(Schmidt 1924)이다. 이 설에 의하면, 슈트의 정단분열 조직이 초층과 내체라고 하는 두 부분으로 구성된다(Fig.
4). 이 두 부분은 그 세포들의 분열면(分裂面)에 의해서 구분된다. 초층과 내체는 각각의 시원세포들에 의해서 형성되는데, 초층의 시원세포들은 중앙축의 위치에 있으 며, 내체의 시원세포들은 초총의 아래에 있다(Esau 1977).
일반적으로, 피자식물의 슈트 정단분열조직에서 2개의 초 층 가운데 가장 바깥쪽 및 바로 안쪽에 있는 ‘층(layer)’을
‘L’로 표시하여, 각각 L1 및 L2로 나타낸다. 초층-내체 구 조에서, 초층 아래에 있는 내체를 구성하는 세포들은 모 든 면(방향)으로 분열하여 부피를 증가시킨다. 이 내체를 L3으로 나타낸다(Fig. 4).
초층은 슈트 정단부의 맨 바깥층을 형성하며, 표면에 대 하여 오직 수직 방향으로만 분열(수직분열)한다. 즉, 이 분열은 분열조직에서 세포의 층 수를 증가시키지 않고 슈트의 정단부 표면을 덮는 세포층(표피)을 만든다. 대부 분의 피자식물들에서, 초층의 수는 종에 따라 1~8개로 다 양하다. 그러나 연구된 진정쌍자엽식물들의 반 수 이상에 서 초층의 수는 2개(L1~L2)이다(Gifford and Corson 1971).
또한 대부분의 단자엽식물에서 초층은 단 1개(L1)이나 (Carles and Fletcher 2003), 단자엽식물 가운데 김의털아과 (亞科) 및 기장아과 식물들에서 초층의 수는 각각 2개 및 1개인 것이 보통이다(Brown et al. 1957). 단자엽식물인 옥 수수(Zea mays)에서 초층의 수는 단 1개이다(Esau 1977).
바깥 쪽 층(L1)에서 표피조직이 기원되며, 그 안쪽에
있는 나머지 층에서 표피조직 이외의 조직들(기본조직
및 유관속조직)이 기원된다. 그러나 Evert (2006)는 다음
과 같이 기술하고 있다. 즉, L1층이 거의 완벽하게 수직
분열만 일어난다고 하더라도, 식물 종 및 초층의 수에 따
라서 L1 층의 세포들이 수평분열하는 경우가 생기며, 이
런 경우에는 안쪽에 위치하던 딸세포들이 L2 층으로 이
Fig. 5 Diagrams of the cytohistological zonation organization of seed plant shoot apices. The apices are represented by the layer and zonal models. Arrows indicate the direction of cell division. A:
Organization of a gymnosperm (Ginkgo biloba) shoot apical meristem (SAM). The SAM is consisted of apical-initial zone (aiz) at the outermost center, central mother-cell zone (cmcz) below the aiz, transitional zone (tz) extended radially from the cmcz, pheripheral zone (pz) derived from the tz and surrounding the cmcz, and rib zone (rz) below the tz. LP1 and LP2, the first and second leaf primordia.
B: Organization of an angiosperm SAM. The SAM is consisted of central zone [tunica-initial zone (tiz) plus corpus-initial zone (ciz)] at the cental surface, and pheripheral zone (pz) and rib zone (rz). The last two zones are derived from the dividsion of cells in both of the tiz and the ciz. LP, leaf primordium.
동되어 L2층에서 유래된 것처럼 분화한다. 그리고 이와 비슷하게 L2 층과 L3층의 딸세포들 사이에도 이동이 생 긴다. 그래서 기본분열조직 및 전형성층이 초층 또는 내 체, 또는 양쪽 모두에서 기원되기도 한다(Evert 2006). 이 러한 이유로 인하여, 내체로부터 어떤 조직이 유래되는 지 그 발생적 운명을 정확히 예측하는 데에는 초층-내체 모형이 적합하지 않은 것으로 판단된다. 그러나 Hanstein (1868)이 제안한 조직원의 체제와 다르게, 초층-내체 모 형은 피자식물에서 슈트 정단부의 특징을 해부학적으로 기재하기에는 적당하다(Moore et al. 1998).
초층-내체 모형은 피자식물의 슈트 정단부 체제를 설 명하기 위해 제안된 것으로써, 나자식물의 정단분열조직 의 특징을 설명하기에는 적당하지 않은 것으로 밝혀졌다 (Foster 1938; Johnson 1951; Gifford and Corson 1971). 특히, Gifford와 Corson (1971) 그리고 Cutter (1980)에 의하면, 매 마등속(Gnetum) 식물, 마황속(Ephedra) 식물, 그리고 다수 의 송백류식물 등을 제외한 나머지 나자식물들은 슈트 정단부에 수직분열을 하는 표면의 초층이 없다. 따라서 이런 나자식물들은 수직으로만 분열하는 안정적인 초층 을 갖고 있지 않으므로, 초층-내체 모형을 나자식물의 정 단분열조직에 적용하는 것은 적합하지 않은 것으로 해석 된다.
세포조직학적 구역화설
미국의 식물학자 포스터(Adriance Foster)는 은행나무(Ginkgo
biloba) 등 나자식물의 정단부에 있는 초층-내체에서 내체를 구성하는 세포들이 동일하지 않다는 사실을 확인하 였다. 또한 그는 초층 아래의 내체를 구성하는 세포들의 분열면, 세포학 및 조직학적 차이, 세포들의 분열조직 활 성 정도 등과 같은 특징을 바탕으로 “세포조직학적 구역화 설(細胞組織學的 區域化說, cytohistological zonation theory)”
을 제안하였다(Foster 1938, 1943). 그 후, 이 구역화 모형 은 다른 나자식물과 대부분의 피자식물에도 적용될 수 있다는 사실이 확인되었다(Gifford and Corson 1971).
많은 해부학자들(Cutter 1980; Mauseth 1988; Fahn, 1990;
Beck 2005; Evert 2006)에 의하면, Figure 5A에서처럼, 많은 나자식물의 정단부에서, 맨 바깥 층의 중앙에는 한 층의
‘정단시원세포구역(頂端始源細胞區域, apical-initial zone, aiz)’이 있다. 이 시원세포들이 수직분열 및 수평분열을 하여 주변의 모든 조직이 기원된다. 그 아래에 비교적 크 기가 크고 다소 액포화되어 있으며 느리게 분열하는 세 포들이 ‘중앙모세포구역(central mother-cell zone, cmcz)’을 이룬다. 이 구역은 ‘주변구역(pheripheral zone, pz)’으로 둘 러싸인다.
그래서 주변구역은 3차원적으로 중앙모세포구역 주위
를 둘러싸는 원통 모양을 형성한다. 주변구역의 세포들 은 크기가 작고 핵이 뚜렷하며 짙게 염색되는 세포들로 구성되며 분열을 활발하게 하는 것이 특징이다. 이 구역 으로부터 기본조직과 유관속조직, 그리고 잎 및 곁눈 등 이 기원된다. 중앙모세포구역의 세포들은 그 구역으로부 터 방사상으로 뻗어나오면서 배열되어 활발히 분열하는 세포들로 구성된 ‘전이구역(transitional zone, tz)’을 형성한 다. 전이구역의 아래에는 ‘늑(肋)구역(rib zone, rz)’ 또는
‘수(髓)분열조직구역(pith meritem zone)’이 있으며, 이 구 역의 세포들은 주로 횡으로 분열하기 때문에 그 딸세포 들은 종(縱)으로 열(列)을 이루어 배열된다. 이 구역은 수 (隨)로 분화되며, 이 세포들의 분열 활성으로 줄기에서 마디 사이가 확장된다.
대부분 피자식물의 슈트 정단부는 초층-내체의 체제
[층모형(層模型), layer model]화 함께 나자식물에서와 비
슷한 세포조직학적 구역화의 체제[구역(區域)모형, zonal
Fig. 6 Diagram of the méristème d’attente (waiting meristem) of an angiosperm (Cheiranthus cheiri) shoot apex. The shoot apical mesitem is consisted of (i) méristème d’attente (ma) (=
waiting meristem) at the center, anneau initial (ai) (= initiating ring) surrounding the ma, and méristème medullaire (mm) (=
pith meristem) below the ma. PL1 and PL2, the first and second leaf primordia [figure adapted from Gifford (1954)].
model]를 갖고 있다는 것이 잘 밝혀져 있다(Gifford 1950;
Popham and Chan 1950). Gifford (1954)는 쌍자엽식물의 슈 트 정단분열조직에서 Figure 5B와 같은 모형을 제시하였 다. 즉, 정단부에서 맨 바깥 층(초층) 중앙에 ‘초층시원세 포구역(tunica-initial zone, tiz)’이 있으며, 이 세포들은 크 고 밝게 염색되는 특징이 있다. 그 아래에는 나자식물의 중앙모세포구역과 비슷한 ‘내체시원세포구역(corpus-initial zone, ciz)’이 있으며, 이 세포들은 옆에 있는 주변구역의 작고 짙게 염색되는 세포들에 비해 밝게 염색되는 특징 을 보인다. 이 두 시원세포구역들을 ‘중앙구역(central zone, cz)’이라고 하며, ‘주변구역(peripheral zine, pz)’으로 둘러 싸여 있다. 주변구역 세포들은 부분적으로 초층의 바깥 층(L1) 및 안쪽층(L2) 그리고 내체(L3) 등에서 기원된다.
입체적으로 보면, 원통상의 주변구역 및 중앙구역 바로 아래에 ‘늑구역(rib zone, rz)’이 있다.
여러가지 방법으로 슈트 정단부의 서로 다른 부위에서 일어나는 유사분열의 빈도를 조사한 결과, 초층시원세포 구역과 내체시원세포구역으로 구성된 중앙구역의 시원 세포들은 다른 구역의 세포들보다 상대적으로 분열을 활 발하게 하지 않는 것으로 밝혀졌다(Corson and Gifford 1969;
Davis et al. 1979; Lyndon 1970, 1976, 1998). Moore 등(1998) 에 의하면, 옥수수(Zea mays)의 슈트 정단부 전체에서 유 사분열이 활발하게 일어나서, 세포들이 중앙구역에서 주변 의 분열조직 쪽으로 첨가된다. 그러나 해바라기(Helianthus
annuus)의 슈트 정단분열조직에서 유사분열 지수는 중앙구역 세포들보다 이를 원통 모양으로 둘러싸고 있는 주 변구역의 세포들에서 3.5배 이상 높다(Steeves 2006).
애기장대의 슈트 정단분열조직의 중앙구역에 있는 세 포들의 유사분열 지수는 주변구역에 비하여 약 50% 더 낮은 것으로 밝혀졌다(Laufs et al. 1998). 그러나 중앙구역 에 있는 시원세포의 수는 대단히 일정하게 유지된다. 이 런 현상은 돌연변이의 발생 빈도를 낮추어서 유전체를 변화 없이 보호하기 위한 것으로 설명될 수 있다(Irish and Sussex 1992). 이러한 생각은, DNA 복제 및 수선에 관여 된 효소들이 중앙구역(시원세포들)에서 평균 이상으로 많 다는 연구 결과에 의해 뒷받침된다(Dodsworth 2009).
DNA 복제가 오류없이 진행됨으로써 돌연변이가 증가 하는 것을 방지하고 슈트 정단분열조직에서 생기는 딸세 포들에서 DNA 이중가닥 절단(double strand break)을 방지 하는데 필수적이며, 그렇지 못할 경우 식물의 발달이 저 해될 것이다(Yadav et al. 2009). 그러나 이러한 중앙구역 에 이웃해 있는 주변구역에서는, 세포들이 빠르게 분열 하여 엽원기 등의 기관 발달에 필요한 수백만 개의 세포 들을 공급한다(Clark 2001; Reddy et al. 2004; Williams and Fletcher 2005; Reddy 2008). 이와 같이 분열조직의 중앙구 역에 있는 세포들은 활발하게 분열하지 않기 때문에, 유 전적으로 건강한 상태로 남아 있으며, 이것은 근단분열
조직의 ‘분열지연중심부(quiescent center)’와 비슷하다고 볼 수 있다(Evert 2006).
식물체의 슈트 정단부에서 세포조직학적 구역화가 이 루어지는 발달 시기는 종에 따라 다양하다. 선인장과의 일부 종에서는 발아하는 시기에 이미 구역화가 이루어져 있는 반면, 다른 종에서는 초층-내체의 체제만 이루어져 있으며, 잎이 30개 이상 발달할 때까지도 구역화가 이루 어지지 않는 종도 있다(Mauseth 1978). 꿀풀과의 콜레우 스속(Coleus) 식물의 슈트 정단부에서는 5쌍의 잎이 발달 할 때까지 구역화가 이루어지지 않으며(Saint-Come 1966), 토마토(Solanum lycopersicum)의 슈트 정단부에서는 구역 화의 유형을 확인할 수 없다(Sekhar and Sawhney 1985).
대기분열조직설
한스타인의 조직원설을 제외하고, 1940년대까지 제안되 었던 슈트 정단분열조직에 관한 설들은 일반적으로 세포 분열 양상을 기초로 한 구조를 기반으로 하였다. 그 후, 주로 세포분열의 빈도가 기능적으로 얼마나 중요한가를 확인하기 위한 연구를 하였다. 그래서 1952년에 프랑스 의 식물학자 뷔바(Roger Buvat)는 “대기(待機)분열조직설 [méristème d’attente (waiting meristem) theory]”을 제안하였 다. 이 설은 특히 뷔바를 중심으로 한 프랑스와 벨기에의 세포학자들에 의해 지지되었다(Buvat 1952; Wardlaw 1957;
Cutter 1959). Figure 6에서처럼, 대기분열조직은 바깥쪽의
2층(proméristème sporogéne(= sporogenous promeristem))과
안쪽 층(proméristème réceptaculaire(= receptacular promeristem)) 으로 구성된다. 이 두 부위는 앞서 언급된 초층-내체 모 형에서 각각 초층 및 내체의 위치에 해당하며, 또한 이 두 부위는 세포조직학적 구역 모형에서 중앙구역에 해당 한다(Kwiatkowska, 2008).
뷔바는 분열조직의 중앙구역에서 일어나는 세포분열 의 빈도를 대단히 중요시하였다. 그래서 유사분열의 빈 도, 세포학적, 조직학적, 미세구조적 연구 결과를 바탕으 로 하여 이 설을 제안하였다(Evert 2006). 즉, 배의 시기에 슈트 정단분열조직이 형성된 후, 영양생장 하는 시기 동 안에 정단부의 중앙구역에 있는 분열조직 세포들이 분열 을 활발하게 하지 않고 기다리고(待機하고) 있다. 그 후, 생식생장 하는 시기가 되면 분열조직의 활성이 회복된다.
영양생장 하는 시기 동안에 분열조직의 활성은 대기분열 조직을 둘러싸고 있는 원통형 구역인 ‘anneau initial (시원세 포환 (始原細胞環), initiating ring)’과 ‘méristème medullaire (늑 (肋)분열조직, rib meristem)’에서 주로 일어난다(Fig. 6). 시 원세포환은 앞서 언급된 모형에서 ‘주변구역’에 해당한 다(Kwiatkowska 2008).
이 설은 원래 쌍자엽식물에 적용되었으나, 그 후 단자 엽식물 그리고 일부의 나자식물 및 무종자 유관속식물에 까지 적용되었다(Steeves 2006). 영양생장 및 생식생장 하 는 시기에, 슈트 정단부의 중앙구역 세포들이 유사분열 을 하여 슈트의 모든 조직 및 기관을 형성하게 된다는 사 실이 많은 연구자들에 의해 여러 가지 방법으로 증명되 었다(Wardlaw 1957; Clowes 1959; Gifford et al. 1963; Irish and Sussex 1992). 따라서 이 설은 일부 연구자들에 의해 지지되었지만, 이전에 정단부의 시원세포들로 간주되었 던 중앙구역에 위치한 세포들이 영양생장 하는 시기 동 안에는 분열하지 않고 있다가 나중에 생식 구조의 발달 을 위한 시원세포들의 역할을 한다는 해석은 반대론자들 의 동의를 얻지 못하였다(Cutter 1980; Steeves and Sussex 1989).
애기장대의 슈트 정단분열조직
십자화과에 속하는 애기장대(Arabidopsis thaliana)는 (i) 세 대 시간이 짧고(약 5 ~ 6주), (ii) 비교적 작은 크기의 유전 체(약 125 Mb) (5개의 염색체에 약 27,000개의 단백질 암호 화 유전자)를 가지며, (iii) 식물체의 크기가 작고(20 ~ 25 cm 의 높이)(조직도 단순하여 해부학적 연구에 유용함), (iv) 많은 종자를 맺으며(개체 당 약 10,000개 이상), (v) 돌연 변이가 많아서 그 유전자의 기능을 쉽게 이해할 수 있는 등의 특징을 갖고 있다(Meinke et al. 1998; The Arabidopsis Genome Initiative 2000; Meyerowitz 2001; Page and rossniklaus 2002; Alonso et al. 2003; Lamesch et al. 2011). 이런 특징은
식물학의 여러 분야를 연구하는 데 있어서 커다란 장점 (이점)으로 작용하기 때문에 연구를 위한 모델 식물로서 널리 이용되고 있다. 특히 이 식물을 이용하여 식물의 생 장과 발달에 필요한 복잡한 과정들을 발생학, 생리학, 그 리고 분자 유전학적으로 이해하는 데 있어서 커다란 진 전을 이루었다.
배발생 시기에 슈트 정단분열조직의 발달
1990년대에 애기장대를 대상으로 한 분자유전학적 연구 가 이루어짐으로써, 슈트 정단분열조직의 기능은 물론, 정단분열조직이 어떻게 형성되며 유지되는가를 더 자세 히 이해하게 되었다(Barton 2010). Figure 7과 같이, 애기장 대에서는 비교적 늦은 배발생 시기에, 즉 자엽이 발달하 기 시작한 후에, 슈트 정단분열조직이 형성된다(Barton and Poething 1993).
애기장대의 배에 있는 슈트 정단분열조직에서, 세포들 을 미분화된 상태로 유지시키거나 분열을 계속하게 하는 유전자들이 확인되었다. 이 유전자들의 역할은 두 가지 의 돌연변이, 즉 분열조직의 크기를 감소시키거나 사라 지게하는 그리고 분열조직을 점차 확대시키는 돌연변이 에 의해 밝혀졌다. SHOOTMERISTEMLESS(STM)라는 유전 자는 슈트 정단분열조직을 형성하여 식물체에서 일생 동 안 분열조직을 유지시키는 작용을 한다. Figure 7에서처 럼, 이 유전자는 늦은 구형기 배(globular embryo)에 있는 1 ~ 2개의 세포에서 최초로 발현된다(Long et al. 1996; Long and Barton 1998). STM 유전자의 m-RNA는 중앙구역 및 주 변구역에서 나타나며 발달하는 엽원기에서는 나타나지 않는다(Long et al. 1996). 이 기능이 심하게 상실된 유전 자(stem)를 갖는 돌연변이는 정상적인 뿌리, 상배축, 자엽 등을 갖지만, 결국 정단분열조직이 형성되지 않는다 (Barton and Poething 1993).
정단분열조직 및 시원세포의 기능을 유지시키기 위해 필 요한 WUS (WUSCHEL homeobox)라고 하는 유전자는 16-세 포기 배에서 발현되기 시작한다(Fig. 7). WUSCHEL 유전 자의 이름(‘WUS’로 줄여서 씀)은 “곱슬곱슬한 모양”이라 는 뜻의 독일어 “wuschelig”에서 기원되었다. 이 낱말은 돌연변이 유전자(wus)가 발견되었던 애기장대가 지닌 겉 모양의 특징을 표현하기 위해 사용되었다.
성숙한 슈트 정단분열조직의 구조
1952년에 Vaughan은 애기장대의 슈트 정단분열조직에서
세포의 크기, 분열 빈도, 내용물의 밀도 등을 고려한 해
부학적 구조를 관찰하였다(Vaughn 1952). 즉, 중앙 종단면
에서 직경이 약 53 μm인 아주 작은 슈트 정단분열조직
은, 가운데에 ‘중앙시원구역(central initiation zone)’과 그
Fig. 7 Formation of shoot apical meristem (SAM) during Arabidopsis thaliana embryogenesis. Process of the embryogenesis: Division of the zygote produces two daughter cells: a small apical cell and a larger basal cell. The apical cell is the precursor of the embryo proper. The apical cell divides transversely and vertically, resulting in an 8-celled proembryo. From the upper four cells a SAM and cotyledons are originated, while from the lower four cells a hypocotyl is derived. The uppermost cell of the suspensor divides transversely and the upper cell becomes a hypophysis. The hypophysis produces the central cells of the root apical meristem and columella of a rootcap.
Gene expression in the SAM during the embryogenesis: The first indication of the SAM development is the initiation of WUS gene expression at the 16-cell stage embryo, long before the SAM is recognizable. Subsequently STM and CLV1 genes are initiated to express. The onset of STM expression is independent of WUS activity, and the initiation of CLV1 expression is independent of STM.
Bars indicate the stages at which the mRNA for each of the genes (WUS, STM, CLV1/CLV3) is detected [figure adapted from Lenhard and Laux (1999) and Evert (2006)].
좌우에 ‘측면분열조직(flank meristem),’ 그리고 중앙시원 구역 아래의 좌우에 각각 ‘형성층유사구역(cambium-like zone)’과 ‘늑(肋)분열조직(rib meristem)’ 또는 ‘종렬분열조 직(file meristem)’ 등으로 구성된다. 또한 Vaughan은 세포 조직학적 방법을 이용한 연구에서 중앙시원구역의 폭은 3 ~ 4개의 세포로 이루어진다고 하였다(Vaughn 1955). 그 러나 1990년대부터 애기장대를 이용하여 식물의 생장과 발달을 연구하는 분자 유전학자들은 Figure 8B에서처럼 초층-내체설에서 유래된 층(層)모형과 세포직학적 구역화 설에서 유래된 구역(區域)모형을 함께 적용하여 사용하 고 있다.
애기장대의 슈트 정단분열조직은 비교적 평평하고(Fig.
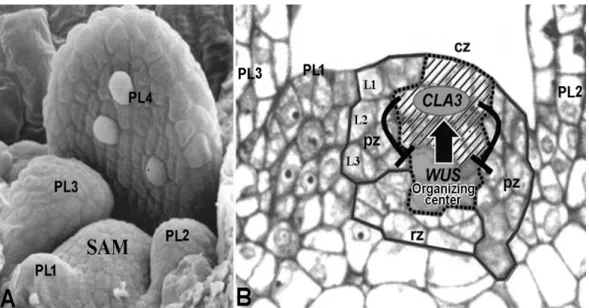
8A)(Bowman 1994), 직경은 약 53 μm이며(Vaughn 1952, 1955), 높이와 폭은 약 40 × 60 μm이다(Barton 2010). 그리고 이 식물의 슈트 정단분열조직은 약 500 여 개의 세포로 구 성되며, Figure 8B에서처럼 해부학적으로 뚜렷한 세포 층 (層)과 특정한 구역(區域)으로 나누어질 수 있다(Reddy 2008).
즉, 애기장대에서 영양생장 하는 슈트 정단분열조직은 중앙 종단면에서 3개의 층(L1, L2, L3)으로 구별되며, 2개 의 초층(L1, L2)과 그 아래층인 내체(L3)로 구성된다(Vaughn
1955, Medford et al. 1992). 정단부에서 바깥 쪽 층(L1)의 시원세포들은 수직분열만 하여 표피조직이 기원되며, 그 아래 층(L2)의 시원세포들도 거의 모두 수직분열을 하여 기본조직(잎에서는 엽육조직)을 형성하고, 맨 안쪽 층(L3) 의 시원세포들은 여러 방향으로 분열하여 잎과 줄기의 유관속 조직을 형성한다(Meyerowitz 1997; Weigel and Jurgens 2002; Williams and Fletcher 2005).
이러한 초층-내체의 체제는 앞서 설명된 피자식물의
슈트 정단부에서처럼, 3개의 세포 층(L1 ~ L3) 위에 기능
적으로 그리고 세포조직학적으로 구별되는 3개의 구역
으로 구별된다. 즉, Figure 8B에서처럼, 중앙구역(cz)에는
약 5개 세포의 깊이와 3 ~ 4개 세포의 폭으로 된 시원세
포들이 있으며, 짙게 염색된 세포들로 구성된 주변구역
(pz)에서 잎 등의 기관이 분화하며, 중앙구역 아래에 있
는 늑구역(rz)에서는 정단분열조직을 지지하는 줄기가 분
화한다(Lyndon 1998). 슈트 정단분열조직을 구성하는 몇 개
영역의 크기와 기능을 더 자세히 확인할 수 있는 방법
(live-imaging)을 이용한 최근 연구에 의하면, 애기장대에
서 중앙구역의 시원세포 수는 약 35개로 밝혀졌으며, 이
것은 이전에 세포학적 방법만으로 관찰했을 때 약 9개의
시원세포가 있을 것으로 예상했던 것과 비교했을 때 상
Fig. 8 Matue shoot apex of Arabidopsis thaliana. A: Scanning electron micrograph of the shoot apical meristem (SAM) surrounded by four leaf primordia (PL1, PL2, PL3, and PL4) (figure adapted from http://www.mun.ca/biology/desmid/brian/BIOL3530/DEVO_07/ch07f13.jpg).
B: Organization and molecular basis of formation and maintenance of the SAM. Median longitudinal section showing the SAM and three leaf primordia (PL1, PL2, and PL3). The SAM is composed of the central zone (cz) indicated by dashed line, peripheral zone (pz), and rib zone (rz). The cz is consisted of the initial cells in L1, L2, and L3 (correspond to the tunica-initial zone and the corpus-initial zone, respectively, in Fig 5B) and the underlying organizing center. CVL3 gene expression in the initial cells (shaded area of L1, L2, and L3,) and WUS gene expression in the organizing center (underlying of shaded area,) interact in a negative feedback loop (T bar), and thus the size of SAM is maintained constantly (figure adapted from http://www.mun.ca/biology/desmid/brian/BIOL3530/
DEVO_07/ch07f13.jpg).
당한 차이를 보인다(Stewart and Dermen 1970; Reddy and Meyerowitz 2005; Yadav et al. 2009).
슈트 정단분열조직의 형성과 유지를 위한
CLV-WUS 신호전달 회로
Figure 8B에서처럼, 발달된 슈트 정단분열조직의 중앙구 역에서 L3층[Fig. 5B에서 내체시원세포구역(ciz)] 바로 아 래에는 약 10개의 세포로 구성된 ‘형성중심부(organizing center)’라고 하는 부위가 있다(Dodsworth 2009). WUS 유전 자는 이 형성중심부에만 제한되어 발현되며, 슈트가 발 달하는 전 기간 동안 계속 발현된다(Mayer et al. 1998;
Vernoux et al. 2000). WUS 유전자의 기능이 상실된 돌연변 이(wus)에서는 슈트 정단분열조직의 성숙이 일찍 완료되 므로, 시원세포들이 그의 독자성(identity, 또는 특성)을 유 지하지 못하고 엽원기 등의 기관을 형성하는 데 쓰이고 만다(Laux et al. 1996; Mayer et al. 1998; Brand et al. 2000;
Schoof et al. 2000). WUS가 발현되면 필연적으로 시원세포 는 기관(예: 잎)이 발생할 곳으로 이동하기 때문에, WUS의 발현이 슈트 정단분열조직의 끝 부분(정단부)에서 반드 시 제한적으로 일어나야만 한다. 그래서, Figure 8B에서 처럼, WUS 유전자는 시원세포구역에서 발현되지 않으며, 이 유전자가 발현되기 위해서는 형성중심부에 있는 세포 들과 그 위에 있는 중앙구역의 시원세포들 사이에 신호
전달이 이루어져야 한다(Gallois et al. 2002).
WUS 유전자는 분열조직에서 L3의 깊숙한 곳에 위치한
형성중심부에서 발현된다. 형성중심부는 그 위에 자리하 고 있는 이웃 세포들에게 시원세포의 독자성을 지니게 하 는 반면, CLAVATA (CLV) 유전자들(CLV1/CLV3)로부터 발 생된 신호는 형성중심부에서 WUS가 발현되어 시원세포 가 형성되는 것을 음성적으로 저해한다(Fig. 8B). CLAVATA 유전자의 이름(‘CLV’로 줄여서 씀)은 애기장대의 열매(長 角果, silique)가 “방망이 또는 곤봉 모양”이라는 영어 “clavate”
에서 유래되었다. 야생형(CLV) 개체의 열매는 긴 방망이 모양인 반면, 돌연변이(clv)의 것은 짧은 방망이 모양이다.
그래서 CLV3는 WUS의 발현을 억제하는 신호로서 작 용하여, WUS와 더불어 ‘음성되먹임회로(negative feedback loop)’
를 형성한다(Lenhard and Laux 1999; Fletcher and Meyerowitz 2000, Boscá et al. 2011). 이러한 회로에 의해, 슈트 정단분열조 직의 크기를 일정하게 유지시킬 수 있다. 이와 같이 슈트 정단분열조직에서 WUS 유전자가 그 위에 있는 이웃 세 포들에게 시원세포의 특징을 지니도록 조절하기 때문에,
WUS 유전자가 발현되는 부위를 ‘형성중심부’라고 한다.위와 같이 STM 및 WUS 유전자들은 분열조직의 형성과
유지를 촉진하는 반면, CLV 유전자들(CLV1, CLV2, CLV3)
은 시원세포의 활성을 억제하여 분열조직의 크기를 조절
한다. 기능을 상실한 돌연변이(clv3)는 WUS의 발현 영역
이 확장됨에 따라 중앙구역에 미분화된 세포(시원세포)
들을 축적시켜서 슈트 정단분열조직의 크기를 증가시킨 다(Clark et al. 1995; Kayes and Clark 1998). clv3가 과잉으 로 발현되면, 슈트 정단분열조직의 성숙을 일찍 종료시 킬 수 있다(Mayer et al. 1998; Brand et al. 2000; Miwa et al.
2009). CLV3는 중앙구역의 L1 및 L2 층 그리고 약간의 L3 층 세포에 제한되어 발현되며, CLV1은 L3 층에 있는 대 부분의 세포들에서 발현된다.
STM은 분열조직 전체에 걸쳐 발현되며, CLV 및 WUS
와는 무관하게 작용하고, 분화가 일어나지 않도록 저해 하는 작용을 한다. 그래서 이 유전자는 슈트 정단분열조 직에서 세포들의 운명이 결정되지 않은 상태로 유지되도 록 한다(Endrizzi et al. 1996; Scofield and Murray 2006). STM 과 WUS는 서로 무관하게 작용하지만 슈트 정단분열조직 에서 상보적인 역할을 한다. 즉, STM은 시원세포의 분화 를 방지하는 반면, WUS는 일부의 세포를 시원세포들로 지 정한다(Lenhard et al. 2002; Williams and Fletcher 2005). 그 러므로 이 두 가지 유전자는 슈트 정단분열조직에서 시 원세포의 유지와 주변구역에서 엽원기 등의 생장 사이의 균형을 유지하는데 있어서 필수적이다.
슈트 정단분열조직으로부터 기관발생의 유전적 조절
슈트 정단분열조직의 중요한 기능 중의 하나는 줄기의 특정한 위치에 그리고 일정한 간격으로 잎이나 꽃과 같 은 부속 기관들을 형성하는 것이다. 따라서 슈트 정단분 열조직은 그 식물의 생장과 발달에 커다란 영향을 미친 다. 잎과 꽃 등의 부속 기관은 슈트 정단분열조직에서 세포 분열의 활성이 활발한 주변구역(peripheral zone)에서 발생된 다. 그래서 이 두 기관의 발생이 시작될 때 형성되는 구조를 각각 엽원기(leaf primordium) 및 화원기(flower primordium)라 고 하며, 이 기관들은 줄기의 측면에서 형성되므로 측재 (側在)기관(lateral organ)이라고 한다.
부속 기관의 하나인 잎의 발생은 처음에 옥수수에서 발견된 KNOTTED1 (KN1) 유전자들(Smith et al. 1992)의 하 향 조절에 의해서 시작된다(Brutnell and Langdale 1998;
van Lijsebettens and Clarke 1998; Sinha 1999). 옥수수에서 이 유전자는 엽원기가 발생될 자리에서 특이하게 하향 조 절된다. 애기장대속(Arabidopsis) 식물에서도 KNOTTED1의 종류에 속하는 KNAT1과 STM1 유전자들(Long and Barton 2000)이 엽원기의 발생 장소에서 하향 조절되는 것으로 밝혀졌다. 이런 유전자들의 무리를 KNOX (‘KNOTTED1 homeobox’에서 생긴 약자임) 유전자라고 한다(Leyser and Day 2003).
엽원기는 슈트 정단분열조직의 층모형(layer model)에 서 2개 또는 그 이상의 층에 걸쳐 분포되어 있는 일단의 세 포들에 의해서 발생이 시작된다. 애기장대속(Arabidopsis) 식물에서 엽원기를 형성하는 세포들은 30개 정도이며(Hall
and Langdale 1996), 엽원기 속에 형성된 전형성층 세포들 에서 PINHEAD (PNH) 유전자의 발현이 매우 높은 것으로 알려졌다(Lynn et al. 1999). 잎이 발생될 장소에서 PNH 유 전자는
STM 유전자가 하향 조절되기 전에 발현된다.잎차례(葉序, phyllotaxis)는 옥신(auxin)에 의해서 조절된 다(Vernoux et al. 2010; Murray et al. 2012). 애기장대속 식 물에서, pin-formed1 (pin1)라는 돌연변이 유전자를 갖고 있는 꽃차례(화서) 정단부에 IAA를 처리하면 꽃이 형성 된다. 정상형 PIN1 유전자 자체는 발달하는 엽원기에서 상 향 조절되는데(Vernoux et al. 2000), 이것은 세포의 확장과 엽원기 형성을 유도하기 위해서는 충분한 양의 옥신이 축적되어야만 한다는 것을 암시한다.
식물이 영양생장(vegetative growth)을 하는 동안에 슈트 정단부에서는 줄기, 잎, 뿌리 등이 발달한다. 낮의 길이나 온도 등의 환경 신호에 의해 슈트 정단부의 대사작용이 변화된다 . 따라서 영양기관이 발달하는 슈트의 정단부, 즉 영양분열조직(vegetative meristem)이 꽃을 형성하는 생식 정단부, 즉 꽃분열조직(floral meristem)으로 전환된다.
슈트 정단분열조직의 가장자리에서 꽃을 형성하기 위 해서는 다음과 같은 두 가지 단백질이 상호작용하는 것 으로 생각되고 있다. 즉, SHOOTMERISTEMLESS (STM) 유 전자 산물(단백질)과 다수의 CUP SHAPED COTYLEDON (CUS) 유전자 산물의 집단이 상호 작용하여 분열조직의 독자성(identity)을 결정한다(Vernoux et al. 2010). 그러므로 애기장대속(Arabidopsis) 식물에서 새로운 기관의 발생이 시작될 때, 위와 같은 분열조직의 독자성을 결정하는 유 전자들은 작동하지 않는다. 이때에 발달하는 원기(原基, primordium)에서 STM과 같은 분열조직의 독자성을 지속 시키는 인자를 억제시키는 ASYMETRIC LEAVES1 (AS1) 유 전자의 전사인자가 관여한다(Byrne et al. 2000).
슈트에 부속된 기관들의 독자성은 꽃을 발생시키는 유 전자인 LEAFY (LFY)의 활성에 의해 조절된다. LFY는 WUS 또는 AGAMOUS (AG)와 같은 유전자들의 전사인자들과 상호 작용하여, 식물과 꽃의 구조를 형성하는데 있어서 중요한 작용을 하는 것으로 밝혀졌다(Nilsson et al. 1998;
Lenhard et al. 2001; Lohmann et al. 2001). 꽃분열조직 자체
의 패턴은 주로 전사의 조절에 의해서 결정되는 것으로
알려졌다(Bowman and Floyd 2008). 이러한 조절에는 기관
의 독자성, 숫자, 대칭성 등을 조절하는 유전자들이 관련되
어 있다 . 예를들면, 애기장대속(Arabidopsis) 식물에서 APETALA1
(AP1), CAULIFLOWER (CAL), PISTILLATA (PI) SEPALLATA1 (SEP1), APETALA3 (AP3) 등과 같은 유전자들은 꽃잎이나꽃받침의 발달을 지시하는 것으로 알려져 있다(Vernoux et
al. 2010). 또한 PERIANTHIA 및 WIGGUM이라는 유전자들은
각 꽃에서 꽃잎과 꽃받침잎의 수를 조절하며, KANADI,
YABBY, 그리고 PHABULOSA-PHAVOLUTA- REVOLUTA 무리 등의 유전자들은 기관의 패턴 형성에 관여한다(Vernoux
et al. 2010).
꽃잎에서, 많은 유전자들이 세포증식과 세포확장 등에 영 향을 미쳐서 꽃잎의 생장을 조절한다(Anastasiou and Lenhard 2007). 즉, JAGGED, ANT, ARGOS 등과 같은 유전자들은 세포증식을 조절하여 꽃잎의 생장에 영향을 미친다(Hu et al. 2003; Dinneny et al. 2004). 반면에 BIGBROTHER, KLUH,
DA1 등의 유전자들은 세포증식의 기간을 조절하여 기관의 최종 크기를 조절한다(Anastasiou et al. 2007; Li et al.
2008).
애기장대속(Arabidopsis) 식물에서, 꽃차례(花序) 분열조 직(inflorescence meristem)을 유지하기 위해 필요한 많은 유전자들이 확인된 바 있다. 그 중의 하나가 TERMINAL
FLOWER 유전자의 산물이다(Conti and Bradley 2007). 이유전자 산물은 이동성 개화(開花) 신호와 관련되어 있다.
FT (애기장대에서 낮의 길이 변화에 반응하여 개화를 촉 진하는 단백질)(Turck et al. 2008), 그리고 TERMINAL FLOWER 단백질 등은 꽃차례 분열조직의 L3 속에 있는 한 장소에 서 분열조직의 위쪽에 위치하는 층으로 이동하는 것으로 보인다(Barton 2010).
결론적으로, (1) 무종자식물에서는 슈트 정단부에 있는 하나의 커다란 정단세포가 분열하여 슈트의 모든 부분들 이 발달되므로, 이 식물군의 슈트 정단부 구조는 정단세 포설로 설명될 수 있다. 그러나 나자식물과 피자식물에 적용될 수 있는 슈트 정단분열조직의 구조로서, 초층-내 체설에서 비롯된 층(層)모형(layer model)과 세포조직학적 구역화에서 제시된 구역(區域)모형(zonal model)이 함께 조 합된 구조가 가장 일반적으로 받아들여져 사용되고 있다.
(2) 어떤 면에서 보면, 슈트 정단분열조직을 구성하는 시원세포들은 기능적으로 동물의 발생에서 줄기세포들 (stem cells)에 해당한다고 할 수 있다. 그러나 식물에서는 배가 발달하기 시작하면서부터 형성된 슈트 정단분열조 직이 계속해서 유지되기 때문에, 배의 시기를 훨씬 지나 서도 여러 가지 구성 요소들로 이루어진 복잡한 식물체 를 계속해서 만들어 낸다. 이것이 식물의 슈트 정단분열 조직이 동물의 줄기세포와 다른 점이다. 또한 슈트 정단 분열조직에 의해서 복잡한 식물체가 지속적으로 발달하 는 현상은 식물체가 무한생장(無限生長, indetermnate growth) 을 할 수 있는 중요한 기작이라고 할 수 있다.
(3) 최근의 십자화과에 속하는 애기장애(Arabidopsis thaliana) 의 연구에서, 슈트 정단분열조직의 형성 및 유지를 조절 하는 유전자들이 밝혀졌다. 즉, STM 유전자는 슈트 정단 분열조직을 형성하며, WUS 및 CLV유전자들이 상호 작용 하여 슈트 정단분열조직의 크기를 조절하여 일정하게 유 지한다. 그 밖에도 다양한 유전자들의 상호 작용에 의해 서 잎이나 꽃과 닽은 기관의 발달이 조절된다. 이러한 슈 트 정단분열조직에 관한 분자 유전학적 연구는 식물학계 에서 식물의 생장과 발달에 관한 새로운 지식을 얻는 데
있어서 중요한 역할을 하고 있다. 이와 같은 분자 유전학 적 분석은 오래전부터 제시되어 발전되어 온 슈트 정단 분열조직의 해부학적 구조(층모형 및 구역모형)에 기초 하고 있다. 이 사실은 식물의 해부학적 연구가 생리학, 발생학, 분자 유전학 등 다른 식물학 분야의 학문적 발전 에 기여하고 있음을 시사한다.
사 사
이 논문은 2014년도 조선대학교 학술연구비의 지원을 받 아 연구되었습니다.
적 요
식물에서 슈트 정단부에는 분열하는 세포들로 이루어진 슈트 정단분열조직이라는 조그마한 부분이 있다. 슈트 정단분열조직은 그 식물의 일생을 통해서 땅 위의 모든 구조를 만들어 낸다. 이 연구의 목적은 지난 250년 동안 슈트 정단분열조직의 체제와 기능을 설명하기 위해 발전 해 온 이론들을 기술하여, 식물의 생장과 발달에 영향을 미치는 슈트 정단분열조직의 중요성을 이해하는 데 있다.
1759년 볼프(C. F. Wolff)가 최초로 슈트 정단분열조직을
기재한 이후, 1858년에 네겔리(C. Nägeli)는 무종자 유관속
식물의 슈트 정단분열조직에서 하나의 커다란 정단세포
를 관찰하여 정단세포설을 제안하였다. 그러나 이 설은
종자식물에 적용할 수 없는 것으로 확인되었다. 이어서
1868년 한스타인(J. Hanstein)에 의해 조직원설이 제한되
었으나, 이 설도 종자식물에 일반적으로 적용할 수 없었
다. 그 후, 1924년 슈미트(A. Schmidt)은 피자식물의 슈트
정단분열조직에서 세포의 분열면이 서로 다른 것을 관찰
하여 초층-내체설을 제안하였다. 이 설은 나자식물에 적용
할 수 없는 것으로 확인되었다. 1938년 포스터(A. Foster)는
세포조직학적으로 서로 다른 구역으로 이루어진 나자식
물의 슈트 정단부를 관찰하여 세포조직학적 구역화설을
제안하였다. 또한 1954년 기포드(E. Gifford)의 연구에 힘
입어 피자식물의 슈트 정단분열조직에도 세포조직학적
구역화설이 적용될 수 있다는 사실이 증명되었다. 또 다
른 설로서, 1952년 뷔바(R. Buvat)는 대기 분열조직설이 제
안되었으나, 영양생장 시기에 분열조직 중앙의 시원세포
들이 분열하지 않는다는 주장이 수용되지 않았다. 최근에
애기장대(Arabidopsis thaliana)를 이용한 연구에서, 슈트 정
단분열조직의 형성과 유지가 몇 개의 유전자들에 의해서
조절된다는 사실이 밝혀졌다. 즉, SHOOTMERISTEMLESS
(STM) 유전자는 슈트 정단분열조직을 형성하고, WUSCHEL
(WUS) 및 CLAVATA (CLV) 유전자들은 슈트 정단분열조직
을 유지시키며, WUS와 CLV 두 유전자들 사이의 신호전 달은 음성되먹임회로를 통해서 이루어진다.
References
Alonso JM, Stepanova AN, Leisse TJ, Kim CJ, Chen H, Shinn P, Stevenson DK, Zimmerman J, Barajas P, Cheuk R, Gadrinab C, Heller C, Jeske A, Koesema E, Meyers CC, Parker H, Prednis L, Ansari Y, Choy N, Deen H, Geralt M, Hazari N, Hom E, Karnes M, Mulholland C, Ndubaku R, Schmidt I, Guzman P, Aguilar-Henonin L, Schmid M, Weigel D, Carter DE, Marchand T, Risseeuw E, Brogden D, Zeko A, Crosby WL, Berry CC, Ecker JR (2003) Genome-Wide Insertional Mutagenesis of Arabidopsis thaliana. Sceince 301:653-656 Anastasiou E, Lenhard M (2007) Growing up to one’s standard.
Cur Opi Plant Biol 10:63-69
Barton MK (2010) Twenty years on: The inner workings of the shoot apical meristem, a developmental dynamo. Dev Biol 341:95-113
Barton MK, Poethig RS (1993) Formation of the shoot apical meristem in Arabidopsis thaliana: An analysis of development in the wild type and in the shoot meristemless mutant.
Development 1119:823-831
Beck C (2005) An Introduction to Plant Structure and Development.
Cambridge University Press, Cambridge, p 86
Bierhorst DW (1971) Morphology of vascular plants. Macmillian, New York
Bierhorst DW (1977) On the stem apex, leaf initiation and early leaf ontogeny in filicalean fern. Am J Bot 64:125-152 Boscá S, Knauer S, Laux T (2011) Embryonic development in
Arabidopsis thaliana: from the zygote division to the shoot meristem. Front Plant Sci 2:1-6
Bowman J (1994) Arabidopsis, An Atlas of Morphology and Development. Springer Verlag, New York
Bowman JL, Floyd SK (2008) Patterning and polarity in seed plant shoots. Annual Rev Plant Biol 59:67-88
Brand U, Fletcher JC, Hobe M, Meyerowitz EM, Simon R (2000) Dependence of stem cell fate in Arabidopsis on a feedback loop regulated by CLV3 activity. Science 289:617-619 Brooks L III, Strable J, Zhang X, Ohtsu K, Zhou R, Sarkar A,
Hargreaves S, Elshire1 RJ, Eudy D, Pawlowska T, Ware D, Janick-Buckner D, Buckner B, Timmermans MCP, Schnable PS, Nettleton D, Scanlon MJ (2009) Microdissection of shoot meristem functional domains. PLoS Genetics 5(5) e1000476 Brown WV, Heimsch C, Emery HP (1957) The organization of the
grass shoot apex and systematics. Am J Bot 444:590-595 Brutnell TP, Langdale JA (1998) Signals in leaf development. Adv
Bot Res 28:161-195
Buvat R (1952) Structure, évolution et fonctionnement du méristème apical de quelques dicotylédones. Ann Sci Nat Bot Biol 13:199-300
Byrne ME, Barley R, Curtis M, Arroyo JM, Dunham M, Hudson A, Martienssen RA (2000) Asymmetric leaves1mediates leaf
patterning and stem cell function in Arabidopsis. Nature 408:
967-971
Carles CC, Fletcher JC (2003) Shoot apical meristem maintenance:
the art of a dynamic balance. Trends Plant Sci 8:394-401 Clark SE (2001) Cell signalling at the shoot meristem. Nature Rev
Mol Cell Biol 2:276-284
Clark SE, Running MP, Meyerowitz EM (1995) CLAVATA3 is a specific regulator of shoot and floral meristem development affecting the same processes as CLAVATA1. Development 121:2057-2067
Clowes FAL (1959) Adenin incorporation and cell division in shoot apices. New phytol 58:16-19
Conti L, Bradley D (2007) TERMINAL FLOWER1 is a mobile signal controlling Arabidopsis architecture. Plant Cell 19:767-778 Corson GE Jr, Gifford EM Jr (1969) Histochemical studies of the
shoot apex of Datura stramonium during transition to flowering.
Phytomorphology 19:189-196
Cutter EG (1959) On the theory of phyllotaxis and histogenesis.
Biol Rev 34:243-263
Cutter EG (1980) Plant anatomy: Experimental and interpretation.
Part 2 Organs. Edward Arnold. p 56
Davis EL, Rennie P, Steeves TA (1979) Further analytical and experimental studies on the shoot apex of Helianthus annuus:
variable activity in the central zone. Can J Bot 57:971-980 Dinneny JR, Yadegari R, Fischer RL, Yanofsky MF, Weigel D
(2004) The role of JAGGED in shaping lateral organs.
Development 131:1101-1110
Dodsworth S (2009) A diverse and intricate signalling network regulates stem cell fate in the shoot apical meristem. Dev Biol 336:1-9
Endrizzi K, Moussian B, Haecker A, Levin JZ, Laux T (1996) The SHOOT MERISTEMLESS gene is required for maintenance of undifferentiated cells in Arabidopsis shoot and floral meristems and acts at a different regulatory level than the meristem genes WUSCHEL and ZWILLE. Plant J 10:967-979 Esau K (1977) Anatomy of seed plants. 2nd ed. Wiley, New York Evert RF (2006) Esau’s plant anatomy. Meristems, cells, tissues of
the plant body-Their structure, function, and development.
John Wiley & Sons, Inc, Publication
Fahn A (1990) Plant Anatomy. 4th ed. Butterworth-Heinemann, Oxford, p 61
Fletcher JC, Meyerowitz EM (2000) Cell signalling within the shoot meristem. Curr Opin Plant Biol 3:23-30
Fletcher JC, Brand U, Running MP, Simon R, Meyerowitz EM (1999) Signalling of cell fate decisions by CLAVATA3 in Arabidopsis shoot meristems. Science 283:1911-1914 Foster AS (1938) Structure and growth of the shoot apex in Gingko
biloba. B Torrey Bot Club 65:531-556
Foster AS (1939) Problems of structure, growth and evolution in the shoot apex of seed plants. Bot Rev 5:454-470
Foster AS (1943) Zonal structure and growth of the shoot apex in Microcycas calocoma (Miq.) A. DC. Am J Bot 30:56-73 Gallois J, Woodward C, Reddy GV, Sablowski R (2002) Combined
SHOOT MERISTEMLESS and WUSCHEL trigger ectopic organogenesis in Arabidopsis. Development 129:3207-3217
Gifford EM Jr (1950) The structure and development of the shoot apex in certain woody Ranales. Am J Bot 37:595-611 Gifford EM Jr (1954) The shoot apex in angiosperms. Bot Rev
20:477-529
Gifford EM Jr, Corson GE Jr (1971) The shoot apex in seed plants.
Bot Rev 37:143-229
Gifford EM Jr, Kupila S, Yamaguchi S (1963) Experiments in the application of H3-thymidine and adenine-8-C14 to shoot tips.
Phytomorphology 13:14-22
Gifford EM Jr, Polito VS, Nitayangkura S (1979) The epical cell in shoot and roots of certain fern: A reevaluation of its functional role in histogenesis. Plant Sci Lett 15:305-311
Groß-Hardt R, Laux T (2003) Stem cell regulation in the shoot meristem. J Cell Sci 116:1659-1666
Haecker A, Laux E (2001) Cell-cell signaling in the shoot meristem.
Curr Opin Plant Biol 4:441-446
Hall LN, Langdale JA (1996) Molecular genetics of cellular differentiation in leaves. New Phytol 132:533-553
Hanstein J (1868) Die Sheitzellgruppe in Vegetationspunkt der Phanerogamen. In: Festschr Friedrich Wilhelms Universtat Bonn. Niederrhein Ges Natur und Heilkunde, Marcus, Bonn, pp 109-134
Hu YX, Xie O, Chua NH (2003) The Arabidopsis auxininducible gene ARGOS controls lateral organ size. Plant Cell 15:1951-1961 Irish VF, Sussex IM (1992) A fate map of the Arabidopsis embryonic shoot apical meristem. Development 115:745-753 Johnson MA (1951) The shoot apex in gymnosperms. Phyto-
morphology 1:188-204
Kayes JM, Clark SE (1998) CLAVATA2, a regulator of meristem and organ development in Arabidopsis. Development 125:
3842-2851
Kessler S, Townsley B, Sinha N (2006) L1 division and differentiation patterns influence shoot apical meristem maintenance. Plant Physiol 141:1349-1362
Kurth E (1981) Mitotic activity in the root apex of the water fern Marsilea vestia Hook. And Grev. Am J Bot 68:881-896 Kwiatkowska D (2008) Flowering and apical meristem growth
dynamics. J Exp Bot 59:187-201
Lamesch P, Berardini TZ, Li D, Swarbreck D, Wilks C, Sasidharan R, Muller R, Dreher K, Alexander DL, Garcia-Hernandez M, Karthikeyan AS, Lee CH, Nelson WN, Ploetz L, Singh S, Wensel A, Eva Huala (2011) The Arabidopsis Information Resource (TAIR): improved gene annotation and new tools.
Nucleic Acids Res 40:D1202-D1210
Laufs P, Grandjean O, Jonak C, Kieu K, Traas J (1998) Cellular parameters of the shoot apical meristem in Arabidopsis. Plant Cell 10:1375-1389
Laux T, Mayer KFX, Berger J, Jurgens G (1996) The WUSCHEL gene is required for shoot and floral meristem integrity in Arabidopsis. Development 122:87-96
Lee KB (2012) Plant Morphology. 2nd ed, Life Science Publishing Co. Seoul, pp 49-51
Lenhard M, Laux T (1999) Shoot meristem formation and maintenance. Curr Opin Plant Biol 2:44-50
Lenhard M, Bohnert A, Jurgens G, Laux T (2001) Termination of
stem cell maintenance in Arabidopsis floral meristems by interactions between WUSCHEL and AGAMOUS. Cell 105:
805-814
Lenhard M, Jurgens G, Laux T (2002) The WUSCHEL and SHOOTMERISTEMLESS genes fulfil complementary roles in Arabidopsis shoot meristem regulation. Development 129:3195-3206
Leyser O, Day S (2003) Mechanism in plant development.
Blackwell Publishing
Li Y, Zheng L, Corke F, Smith C, Bevan MW (2008) Control of final seed and organ size by the DA1 gene family in Arabidopsis thaliana. Genes Dev 22:1331-1336
Lohmann JU, Hong RL, Hobe M, Busch MA, Parcy F, Simon R, Weigel D (2001) A molecular link between stem cell regulation and floral patterning in Arabidopsis. Cell 105:793-803 Long JA, Barton MK (1998) The development of apical embryonic
pattern in Arabidopsis. Development 125:3027-3055 Long JA, Barton MK (2000) Initiation of axillary and floral
meristems in Arabidopsis. Dev Biol 218:341-353
Long JA, Moan EI, Medford JI, Barton MK (1996) A member of the KNOTTED class of homeodomain proteins encoded by the STM gene of Arabidopsis. Nature 379:66-69
Lyndon RF (1970) Rate of cell division in the shoot apical meristem of Pisum. Ann Bot (London) 34:1-17
Lyndon RF (1976) The shoot apex. In: Yeoman MM (ed), Cell Division in Higher Plants. Academic Press, New York. pp 285-314
Lyndon RF (1998) The shoot apical meristem, its growth and development. Cambridge University Press. Cambridge.
Lynn K, Fernandez A, Aida M, Sedbrook J, Tasaka M, Masson P, Barton MK (1999) The PINHEAD/ZWILLE gene acts pleiot- ropically in Arabidopsis development and has overlapping functions with the ARGONAUTE1 gene. Development 126:
469-481
Mauseth JD (1978) An investigation of the morphogenetic mechanisms which control the development of zonation in seedling shoot apical meristems. Am J Bot 65:158-167
Mauseth JD (1988) Plant Anatomy. The Benjamin/Cummings Publishing Co., Inc., Menlo Park, California, p 88
Mayer KFX, Schoof H, Haecker A, Lenhard M, Jurgens G, Laux T (1998) Role of WUSCHEL in regulating stem cell fate in the Arabidopsis shoot meristem. Cell 95:805-815
Medford JI (1992) Vegetative apical meristems. Plant Cell 4:
1029-1039
Medford JI, Behringer FJ, Callos JD, Feldmann KA (1992) Normal and abnormal development in the Arabidopsis vegetative shoot apex. Plant Cell 4:631-643
Meinke DW, Cherry JM, Dean C, Rounsley SD, Koornneef M (1998) Arabidopsis thaliana: A Model Plant for Genome Analysis. Science 282:62-682
Meyerowitz EM (1997) Genetic control of cell division patterns in developing plants. Cell 88:299-308
Meyerowitz EM (2001) Prehistory and History of Arabidopsis Research. Plant Physiol 125:15-19
Miwa H, Kinoshita A, Fukuda H, Sawa S (2009) Plant meristems: