군 췌 애 기
활성화된 단핵구 및 대식세포의 항원제시기농에 대한
Kaempferitrin
의 조절 효과
김병훈 • 조동하 • 조재열^ 강원대학교 BT학부대학생명공학부 (Received October 9, 2007: Revised October 29, 2007)
Modulatory Effect of Kaempferitrin, a 3,7-Diglycosylflavone, on the LPS-Mediated Up-regulation of Surface Co-stimulatory Molecules and CD29-Mediated
Cell-cell Adhesion in Monocytic- and Macrophage-like Cells
Byung Hun Kim, Dong Ha Cho and Jae Youl Qk/
School of Bioscience and Biotechnology, Kangwon National University, Chuncheon 200-701, Korea
Abstract —— Kaempferitrin, isolated from Kenaf (Hibiscus cannabintis), was examined to evaluate its modulatory effects on antigen-presenting cell functions of macrophages/monocytes such as phagocytosis of foreign materials, up-regulation of co
stimulatory molecules (CD40, CD80 and CD8 6), adhesion molecule activation, and antigen processing and presentation.
Kaempferitrin strongly blocked up-regulation of CD40, CD80 and CD8 6, but not pattern recognition receptor (PRR) (e.g., TLR2). It also suppressed functional activation of CD29 (pl-integrins), as assessed by cell-cell adhesion assay, required for T cell-antigen-presenting cell (APC) interaction. Furthermore, this compound did not block a simple activation of CD29, as assessed by cell-fibronectin adhesion assay. However, the compound did not diminish phagocytic uptake, an initial step for antigen processing, and ROS generation in RAW264.7 cells. In particular, to understand molecular mechanism of kaempfer- itrin-mediated inhibition, the regulatory role of LPS-induced signaling events was examined using immunoblotting analysis.
Interestingly, this compound dose dependently suppressed the phosphorylation of kB a, Src, Akt and Syk, demonstrating that it can negatively modulate the activation of these signaling enzymes. Therefore, our data suggested that kaempferitrin may be involved in regulating APC function-relevant immune responses of macrophages and monocytes by regulating intra
cellular signaling.
Keywords □ kaempferitrin, macrophages, monocytes, antigen-presenting cell function, co-stimulatory molecules, adhe
sion molecules
단핵구및대식세포는내재성 면역반응을담당하는대표적인 면역세포다. 이들세포들은외부로부터침입한세균틀에서 유래 된세포구성 성분들이나체내다른면역세포에서분비한IFN-y 와같은사이토카인에 의해 활성화되어 종양괴사인자(tumor necrosis factor-a)와 같은 사이토카인이나 일산화질소(nitric oxide)나활성산소(reactive oxygen species) 등과같은득성물질, 그리고프로스타그란딘류와같은염증매개물질의 분비를조절 한다.« 특별히, 이들세포는세균이나암세포등과같은면역유 발원을탐식하고탐식된면역원을맵 드 로 가 공 주 조 직
본 논문에 관한문의는 저자세게로 (견화) 033-250-6562 (팩스:) 033-253-6560 (E-mail) [email protected]
적합성항원에공여함으로써, T cell의분열및분희를촉진시키 는항원제시 세포(Antigen-presenting cell[APC])로서의 역할을 수행하는것으로알려겨있다.^^ 이때대식세포흑은수지상세포
와T cell간의세포-세포간상호작용이펄수적이며이률조절하
는단백질은CD80, CD8 6 및CD40와같은co-stimulatory mol- ecule들과 pi- 흑은 p2-int:egrins와같은adhesion molecule로 알려져 있다 .^이 들 단백질의 활성및발현은외부자국에 반 응하는pattern-recognition receptor(PRR)과같은수용체의 매 개및이들수용체와결합된신호전달단백질복합체의 활성에 의해조절되는것으로알려져있다. 특별히관련신호전달단백 질은non-receptor type tyrosine kinases(Src, Syk 및JAK2), phosphoinositide-3-kinase(PI3K), Akt(protein kinase B) 및 mitogen-activated protein kinases(MAPK) 등으로구성되어 었
휠성화된 단핵구 및 대식세포의 항원제시기능에 대한 Kaempferitrin 의 조절 효과 483
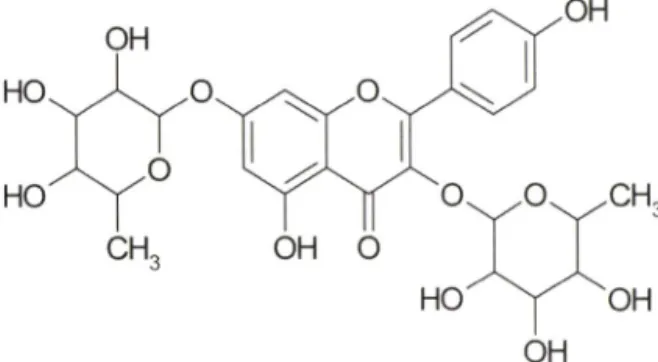
Fig. 1 - Chemical structure of kaempferitrin.
다.® 이들단백질의 활성은최중적으로 NF-kB 및 AP-1 과같은 transcription factor의활성과밀접한관련이 있으며, 이둘의 활 성은염증및내재성 면역반응관련단백질의 발현을매개하게 된다.6.7)
Kaempferitrin(Fig. 1)은남부아시아가원산지이며, 무궁회■속
에속하는Kenaf(Hibiscus 에서주줄한당함유폴라
보노이드게화합물이다.® 현재까지 kaempferitrin에관한약리작 용은많이보고되어 었지않다. 하지만 renal cellular membrane 의보호작용,®* glucose transporter 조절을통한항당뇨작용
및항산화작용,^ > 염증유발물질(종양괴사인자및일산화질소
등) 분비억제를통한항염증작용^' 등이 kaempferitrin에의해 서매개되는것으로보고되고있다. 본연구에서는대식세포및 단핵구에의해수행되는내재성면역반응에 관한 kaempferitrin 의조절효과를검증하고자하였다. 특히염증반응이후진행되 는 대식세포 및단핵구의 항원제시기눙(antigen presentation function)에대한 kaempferitrin의조절기능에초점을맞춰연구 하였기에이를보고하고자한다.
재료 및 방법
실험재료
이실험에서는마우스대식세포주인 RAW264.7 세포와인간
단핵구세포주인 U937 세포를사용하였다. 세포배양을위한배
양용배지 RPMI1640 media, fetal bovine serum(FBS) 그러고 항생제 (penicillin/streptomycin) 는 Hydone(Hydone, South Logan, UT, USA)사제품을사용하였다. Human fibronectin은 BD Biosciences(San Jose, CA USA)를 사용했고, CD80 (MEM-233, purified IgGl), CD86(BU63, purified IgGl), CD29(MEM lOlA, IgGl, ascites) 과 CD43(161-46, ascites, IgGl)는각각 V Horejsi와 R Villela로부터제공받았다. Src, Akt, JAK-2, Syk 및 MBa에대한 phospho-specific 항체는 Cell Sig
naling Technologies사제품을사용하였다. 그밖에 MTT(3-[4,5- dimethylthiazol-2-yl]-2,5-diphinyltetrazolium bromide)를포함한
화학시약은 Sigma 제품(St. Louis, MO, USA)을사용하였다.
세포배양
RAW 264.7 및 U937 세포들은 5% CO^, 37^C incubator에서 배양하였다. 배지는 penicillin(100lU/m/) 및 streptomycin(100
|ig/m/)파 10% FBS가포함된 RPMI 1640을■사용하였다.
세포 생존율 검정
세포성 면역활성 효파률 검증하기 위한 선행실험으로서, kaempferitrin 농도(0, 12.5, 25, 50, 100 및 200 |jM)벌로세포생 존에미처는영향을 MTT assay법을이용하늬분 석 하 였 다96- well plate에 I X 10®개의 RAW264.7 세포률 plating하고 kaemp- feritrin을농도벌로처리하였다. 37°C에서각면역실험 조건에 상응하는배양시간동안 C0 2 incubator에서배양하였다. 이후 10 MTT 용액 (stock concentration: 5 mg/m/)을첨가하고 3시간 동안추가반응을유도하였다. 반응종료및 formazan crystal의 용해를 위해 각 well에 100 |i/ MTT stopping solution(10%
sodium dodecyl sulfate in 0.01 M HCI)을분주하였다. 세포생 존율은 MTT가 formazan으로환원된양을 570 nm에서흡광도
률측정하셔 얻어진 0D 값을통해산출하였다.
Co-stimulatory molecules의 발현 정량
RAW 264.7 세포(IX 10® cells/m/)률 12-well plate에 1 m//well 썩분주후 18시간동안전배양하였다. 앞의실험방법에서와같 이 kaempferitrin 또는약물(각종단백질의 억제자)과 LPS를처 리하여 8시간배앙후, 세포률회수하여, 96 well round plate에 세포수를일정하게분주하였다. 그러고 FACS buffer(2% FBS in PBS)로 세척하였다. 그리고 10% rabbit serum(in PBS)으로 blocking하고, CD80 항체및각종항체률처리하여 45분간배양 후 FACS buffer로세척하였다. 이후, 3.7% formaldehyde로고 정하고 flow cytometery로형광정도를측정하였다.
라디칼 소거농 정량
RAW264.7 세포를 penicillin(100 lU/mi) 및 streptomycin (100 ng/m/pl- 10%의 FBS률함유하는 RPMI 1640 배지룰이 용해서 2xio«cells/m/의 농도로 조절한 후, 6 well plate에 Im/를분주하고, 5% CC>2 및 37>C에서 18시간동안 전배양하 였다.15) 전배양후배지률 제거하고, PBS 495 n/를분주하였다. 그리고 4배농도로조제된 DHR123(최중농도 20 ^M) 250 ^i/를 해당 well에분주후 10분배양하였다. 이후, 200배농도로조 제된 kaempferitrin 5 나/를처리후 30분배양하고, 4배농도로 조제된 SNP 250 |j/(최종농도 0.25 mM>률처러하식 20분배앙 하였다. 라디칼소거 정도는 flow cytometer률이용하여 죽정 하였다.
Vol. 51, No. 6, 2007
buffer에서반응시킨푸특정단백질에특이한 1차항체률상온 에서 1시간반응시켰다. 이후 2차항체를 1시간동안추가적으 로 처리한후, ECL chemiluminescence로빈r응정도률확인하 였다.
통계처리
각 data는얻어진결과는평균±표준편차로나타낸것이며, 그 룹간의통계적유의성은 SAS package를이용하여 P<0.05 수
준에서 Duncan의다중비교법에의해분석하였다.
결과 및 고찰
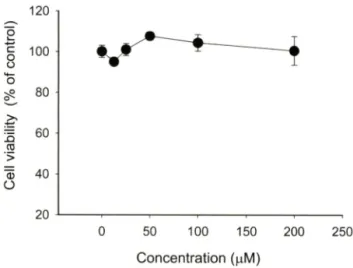
Kaempferitrin의세포독성에미처는효과
우선 kaempferitrin의세포득성 효과률 알아보기 위해 최대
200 nM 농도까지 대식세포및단핵구암세포주에 24시간동안
처리하였다. Fig. 2에서나타난바와같이 kaempferitirin은미우 스대식세포인 RAW264.7 세포에아무런득성효과를나타내지 않았다. 게다가, 사람의단핵구인 U937 세포에도동일조건에서 견혀세포득성 효과률유도하지 않았다(data not shown). 특별 히약리적으로효능이 강한폴라보노이드류에서강한세포득성 이나타난다는것과비교시/가상대적으로본화합물이우수하 고다양한면역약리작 보 일 경우, 세포득성측면에서다른 폴라보노이드류보다 약물학적인 장점을가질 것으로관단된 다. 따라서면역약리학적 효능을조사•하기 위해, 활성화된단핵 구및대식세포를이용하여 kaempferitirin의조절능을연구하여 보았다.
0 50 100 150 200 250
Concentration (|xM)
Fig. 2 - Effect of kaempferitrin on the viability of RAW264.7 cells.
RAW264.7 cells (1x10® cells/m/) were incubated with kaempferitrin for 24 h. Cytotoxicity of this drug was determined by conventional MTT assay as described in Materials and Methods. Data represent mean ± SEM of three independent observations performed in triplicate.
대식세포의 탐식작용 촉정
RAW264.7 세포를 2X0®cella/well로조정하식 6-welI plate에 분주하고 18시간동안견배양하였다. 견배앙푸 kaempferitrin을 처러하고 1시간동안 37X에서배양하였다. 그리고형광으로표 지된 particle FITC-dextran(2 mg/m/)을세포에 처러하고 빛을 차단하여 30분간배양하였다. Cold-PBS로세포를세척하여 미 탐식된 입자를 제거하고 3.7% formaldehyde로 고정 후 flow cytometer로형광정도를측정하였다.오5그
세포써I포간 유착반등 분석
Kaempferitrin을각농도로조제해 96-well plate에각각 10 \d
씩분*주하고, U937 세포(IX 10®cells/m/) 용액을 80 m/를처리하 여 37X에서 30분간배양하였다.^®^ 다시 여기에 10 나/의항체 (CD43 및 CD29 각 1 나g/m/)를분주하여시간벌로사진을촬영 하였다.
세포-fibronectin간의 유착반음 정량
우선 fibronectin(최중농도 50 lig/m/)을 96-well plate의해당 well에 100 m/를분주하고, 나머지 well에는 PBS를분주후, 4X 에서 18시간동안 견배양하였다. RPMI1640 배지로 세척 후 2 mg/m/ bovine serum albumin으로 blocking시켰다. 다른 plate 에여러가지처리농도로희석된 kaempferitrin 11 |i/률분주하 고, U937(5.6x 10*^cells/ml) 세포용액을 99|i/를처리하여 37T 에서한시간동안배양하였다. Blocking한 plate룹 RPMI1640 배지로세번세척후, kaempferitrin이처러된세포용액 100 |i/
윽 fibronectin-coated plate로옮겨 넣고, 4시간추가배양을실 시하였다. 세포-fibronectin 간유착 정도는 0.1% crystal violet 용액을첨가한후 15분동안반응시키고, 10% acetic acid를처
리한후 540nm에서흡광도를측정하였다.^타
표적 단백질의 발현량 검토
RAW 264.7 세포(5X10®cells/m/)틀 6-well plate의해당 well 에 Im / 분주하고, 배지 3m/을더한후 18시간동안견배양하 였다. 앞의 실험방법에서와같이 kaempferitrin을 30분전처리 하고, LPS룰 2분, 30분처리한후, 세포룰회수하였다. 회수된 세포는 lysis buffer(함유조성 : 20 mM Tris-HCl, pH 7.4, 2 mM EDTA, 2 mM ethyleneglycotetraacetic acid, 50 mM p-glyc- erophosphate, 1 mM sodium orthovanadate, 1 mM dithio- threitol, 1% Triton X-100, 10% glycerol, 10 |ig/m/ a protinin, 10 \i^ml pepstatin, 1 mM benzimide, and 2 mM hydrogen peroxide)를처러하여 total cell lysate를조제하였다. 동일한양 의단백질을 10% SDS-polyacrylamide gel에서 견기영동하고, wet-blotting transfer 방법을이용하여 PVDF membrane으로 단백질을 transfer하였다. Membrane을 5% BSA blocking
484 김병문 • 조동•하 • 조재열
0000020864{I0J1U
8 io/JA}!l!qe!>0
10
dectin-1 T LR 2 TLR 4 S R -A C D 11b
! C D 8 0 I C D 86 ] C D 40 I C D 69
0 • unl LPS (1 ng/ml) Kaempferitrin (100 uM)
Bfnl
20
0 - m i
FITC-dextran (1 mg/ml)
Kaempferitrin (fiM) 25 50 100
Fig. 4 - Effect of kaempferitrin on the phagocytic uptake of RAW264.7 cells. RAW264.7 cells (2x10^ cells/m/), pretreated with kaempferitrin, were stimulated with FITC-dextran (2 mg/m/) for 6 h. The extent of the phagocytic uptake was determined by flow cytometric analysis, as described in Materials and Methods.
LPS{1 ng/ml) Kaempferitrin (100 j.iM)
160
N DO SB SP PP2 LY Wort AG Pice LPS
Fig. 3 ~ Effect of kaempferitrin on the LPS-mediated up-regulation of co-stimulatory molecules. (A and B) RAW264.7 cells
( 2 X1 0^ cells/m/) were treated with kaempferitrin in the presence or absence of LPS (1 for 12 h. Surface levels of costimulatory molecules (A) or surface pattern recognition receptors (B) were analysed by flow cytometry, as described in Materials and Methods. (C) RAW264.7 cells (2 X10^ cells/m/) were treated with various inhibitors [UO (U0126. 40 ^M), SB (SB203580, 20 ^M), SP (SP600125, 10 PP2 (40 ^M), LY (LY294002, 40 ^M). Wort (Wort
mannin, 20 (iM), AG (AG490, 40 |iM), Picea (piceatannol, 40 laM)] in the presence or absence of LPS (1 \xg/ml) for
12 h. Surface levels of costimulatory molecules were analysed by flow cytometry, as described in Materials and Methods. Data represent mean ± SEM of three indepen
dent observations performed in triplicate. p<0.05 com
pared to normal group, and *; p<0.05 compared to control group.
Vol. 51 No. 6, 2007
Co-stimulatory molecule의 발현에 머치tz: Kaempferitrin의 효과
Co-stimulatory molecule(eg. CD40, CD80 및 CD86 등)은 활성화된대식세포가 T cell의분열을촉진시키는 항원제시 세 포(Antigen-presenting cell[APC])로서의 역할수행시 필수적으 로요구되는세포막단백질이다.^*^^ 실제로 LPS률처러하게되면 대식세포는활성화되면서정상수준보다증가된 CD40, CD80 및 CD86의막표면발현양태률보이게된다(Fig. 3A). 따라서, 본연 구에서는 Kaempferitrin이 LPS에의해활성화된대식세포에서 높게발현되는 CD40, CD80, 및 CD86의세포막단백질의 발현
수준에미치는영향을 FACS 실험을통해조사하였다. 흥미롭게
도, 이들증가된 CD40, CD80, 및 CD86 발현양은 kaempferitrin
처러시 감소하는 것을확인할 수가 있었다(Fig. 3A). 게다가
kaempferitrin은활성화된대식세포에서 초기에 발현되는 CD69 의상승정도도억제하는것으로나타나초기 대식세포활성 자 체률억제할가능성이 었는것으로관단되었다. 죽, 이는 CD69 가림프구, 단핵구, 혈소관의 초기활성화시 발현이증가된다고 보고되었기때문이다.고®^ 그러나 kaempferitrin은 LPS에의해증
가된 PRR의하나인 TLR-2의발현앙증가에는조절효과를나타
내지 않은 것(Fig. 3B)으로 보아, kaempferitrin의효과는 co
stimulatory molecule의발현앙조절에만관련되어 있는것으로 관단된다. 한편가장발현번화가큰 CD80을이용하여 발현증 가시 될수적인세포내 신호전달과정을조사하였다. Fig. 3C에 따르면, 특별히 Src kinase 저해제인 PP2, PISIVAkt 저해제인 wortmannin, 그리고 Syk 저해제인 piceatannol 처러군에서 CD80
의발현이유의성있게감소된것으로나타났다. 이는 LPS 유도
에의해 발현되는 CD80을 포함한 co-stimulatory molecule은 Src, Akt 및 Syk kinase 활성을필요로한다고사료된다. 실제로
칠성화된 단핵구 및 대식세i 의 항원제시기능게 대한 Kaempferitrin 의 조절 효과 485
y
w
o o
o o
o o
o o
5
0
5
0
5
0
5 3
3
2
2
1
1 {9nle>idiAOw으no아loEAJO;BlnLU!lslou oooooooo
8
6
4
2
{0nlB>Ids}
w^lnoeloLUAJ01elnE!JS-0u
김병훈 • 조동하 • 조재열
Kaempferitrin OxM)
고
25 50 100 200
월 삶 .스^^^율
^ HHHHH
■■■■■
C
Fig. 5 - Effect of kaempferitrin on CD29-mediated cell-cell adhesion. U937 cells were incubated with kaempferitrin in the presence or absence of pro-aggregative (activating) antibodies [antibodies (1 each) to CD29 (MEM lOlA) and CD43 (161-46)] for 2 h. The images of the cells in culture were obtained using an inverted phase contrast microscope attached to a video camera.
NF-kB률포함한이돌효소들이 LPS 매개에 의한대식세포의
pro-inflammatory 단백질생합성에중요한신호전달매개단백
질로알려져 있다.2®>
식세포작용에 미치는 kaempferitrin의효과
내재성 면역반응에서보여지는중요한현상이 식세포(대식세 포와호중구) 매개에의한식세포작용으로알려져있다.2« 특벌 히이과정은세균들을제거하기위한과정이면서동시에 APC 의항원가공 및 제시 기능을 위해 될수적인 전단계 이므로 kaempferitrin의조절작용을조사하여보았다. 그러나 Fig. 4에서 확인되듯이, kaempferitrin은마우스대식세포주인 RAW 264.7
세포에서보여지는 FITC-dextran의식세포작용에이■무런영향
도미처지않는것으로나타났다(Fig. 5).
세포쎄포간 및 세포-m atrix proteii£} 유척빈해 I 미처는 kaempferitrin의효과
세포긔1포간유착반응은 APC 기능수행시보여지는대표적인 반응중하나의다. 죽대식세포혹은수지상세포와 T cell간의상 호작용은 T cell의분열과정조절시 될수적22>이기 때문이다. 특 벌히 이들 과정을조절하는 단백질은 CD18(p2-integrins) 및 CD29(pi-integrins) 등과같은 adhesion molecule들로알려져있 으며, 실제로이들단백질의기능을선택적으로저해하는특이 항체처리시 APC 기능매개에의한 T cell 증식과정이억제된것
으로 보고되었다™ 따라서 APC 기능수행에 필수적인 C 0-
stimulatory molecule의막표면발현을매우효과적으로억제한 kaempferitrin이 adhesion molecule의활성조절은어떤 영향을 나타내는지조사•하고자하였다.
본연구자는 adhesion molecule인 CD29의기능적활성을검 증하기위해서 U937 cell 및 CD29 활성유도항체를이용하쉬, CD29 활성매개성 세포^ 1포간유착모델을확립한바있다.*®
이둘모델은 CD29의기눙매개에의한세포간상호작용을확인
할수있는실험법이므로, 본연구에서 kaempferitrin의효과검 증에 적용하여보았다. Fig. 5에서볼수있듯이, kaempferitrin 은농도의존적으로 CD29 기눙을억제한것으로나타났다. 죽 kaempferitrin은 100 및 200 나M에서강력히 CD29 데개성세포 간유착현상을억제하였다. 그러나, CD43 매개에의한세포간 유착현상은억제하지 않았다(Fig. 5). 이는 kaempferitrin에의한 CD29 기눙억제는딘순히약물비특이적인억제가아닌, adhesion molecule 선택적인것으로사료되었다.
특별히세포간유착과정은세포질내신호전달과정의활성및
이률통한막표면 CD29의활성에서기인된유착과정 메개등으
로구성되어져 있다.고견' 따라서, kaempferitrin에의한세포간유 착과정억제가어떤과정의 간섭에서기인하는지확인하기 위해 먼저 CD29 단백질자체의 활성을측정할수있는세포-matrix protein(fibronectin)간의유착실험모델을이용하여 연구하였다. 동미롭게도, kaempferitrin은 U937 세포가 fibronectin에유착되 는현상을억제하지못한것으로나타났다(Fig. 6). 즉본결과는 kaempferitrin의세포간유착현상억제는 adhesion molecule인
50
0 ■!■'그* DHR123 (20 |.iM) - Kaempferitrin (fiM) v SNP (0.25 -
mM)
25 50 100
+ + +
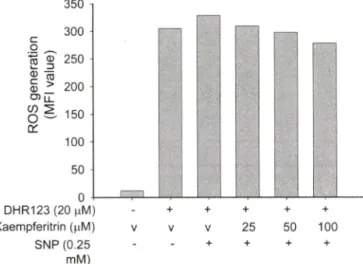
Fig. 7 - Scavenging effect of kaempferitrin on SNP-released radical generation. RAW264.7 cells (2 x 1 0^) were pretreated with kaempferitrin for 30 min and further incubated in the presence of absence of SNP (250 }iM) for 20 min. The level of radicals was determined by flow cytometric analysis as described in Materials and Methods.
Vol. 51, No. 6, 2007
20
0 50 100 150 200 250
C o nce ntration (|aM)
Fig. 6 - Effect of kaempferitrin on cell-fibronectin adhesion. U937 cells (5x10^ cells/well), pretreated with kaempferitrin, were seeded on fibronectin (50 |ag^m/)-coated plates and further incubated for 3 hours. The attached cells were determined by crystal violet assay, as described in Materials and Methods.
CD29 자체의단순한활성억제에서 기인되는것보다는 CD29의
칠성을유도하는세포내신호전달과정억제에의해매개되는것 으로관단된다.
Kaempferitrin의 라디칼 소거능
ROS(reactive oxygen species)는대개산소가필요한대사파 정중에서부산물로생성되는데, 세포의신호전달에서중요한역 할을수행하기도한다. 그러나환경적인스트레스로인해 ROS 가급격히증가되면, 라디칼이 발생되어 세포구조에심각한손
상을초래하여 세포사멸과 같은현상을유도하게 된다.2®' 반면 에면역빈r응에서는활성산소유래라디칼을이용하여 대식세포 나과립구등이 몸안으로침입한미생물이나중양세포를공격 하는 용수단으로사용한다. 병리적으로는이들활성산소중 유래 라디캄이 과량으로분비될경우노화나암 및염증질환의
발생과깊은관련이 있어 ROS의라디칼소거능을가지는폴라
보노이드류같은항산화제둘은관련질환의치료효능이 있는것 으로보고되고 있다.® 따라서, kaempferitrin이대식세포내에서 유도되는라티칼발생에 어떤 영향을미치는지 조사하였다. 그 러나, Fig. 7에서보여지듯이, kaempferitrin은 SNP에의해유러 되는라디칼을중화시키지못하는것으로나타났다. 반면에 대 조군으로사용된토코페롤은강력하게라디칼생성을억제하는 것으로나타났다(data not shown).
억제기전 연구
Fig. 3C에서보여주듯, CD80의막표면발현앙증가률위해서 는 Src kinase, PI3K 및 Syk kinase 활성이 요구된다는 것과, Fig. 5 및 6에서확인되었듯이, kaempferitrin의세포간유착과정
억제는 CD29 자체의 단순한구조적 활성보다는 CD29 활성을
유도하는세포내신호전달과정을억제할가능성등의있다는것
으로고려하여, kaempferitrin의약리작용에 대한억제 기전을
Western blot을통해 세포내신호전달파정측면에서 조사하였
30 min
LPS(1 ^ig/ml) - + + + + +
Kaempferitrin (mM) - 0 25 50 100 200
p-Akt 서 혈 mmmm ^mmm
p-kBa 행P혜T • -
p-Src 따
p-actin
2 min LPS{1 나 g/ml)
Kaempferitrin (나M)
+ + + + +
0 25 50 100 200
p-Syk 쪽* *
P-JAK2
p-actin 램mm mmm
Fig. 8 - Effect of kaempferitrin on LPS-mediated signaling enyme activation in RAW264.7 cells. RAW264.7 cells (5x10^ cells/
ml) were stimulated with kaempferitrin in the presence or absence of LPS (1 for 2 and 30 min. After immuno
blotting, the total or phospho-protein levels of Src, JAK-2, Akt, Syk, IkBcx, and p-actin were identified by their total protein- or phospho-specific antibodies. The results show one experiment out of three.
칠성화된 단핵구 및 대식세포의 항원제시기능에 대한 Kaempferitrin 의 조절 효과 487
000000
5
0
5
0
5
0 3
3
2
2
1
1
{anlB>ElAO
UOIlroJ^U이어woy
40
80
40
{I0JJU8 JO UOIS이I사ujp^uojclljill^o
김병훈 ■ 2동하 ■조재열
다. Fig. 8에서확인되듯이, kaempferitrin은: 농도의존적으로 IkBcx 및 Src 인산화과정을억제하였다. 또한 200|oM에서는 Akt 및 Syk kinase의인산화과정을억제한것으로나타났다. 이는 LPS 신호견달과정시 각신호전달단백질의 인산화과정이 이들단 백질의활성과비례한다는관점에서볼때본약물은 NF-kB 및 Src 활성을강력하게억제할것으로판단되며, 이외에도 Akt 및 Syk 등의활성도고농도에서는억제하는것으로추측된다. 반면 에, 25 및 5(HiM 농도에서는 Akt 인산화과정을증가시킨것으 로볼때, 저농도의 kaempferitrin은인산화과정을조절하는세 포내효소(에를돌면, phosphatase 등) 활성에 영향을즐수있 을것으로관단된다. 또다른한편으로, 당을함유한구조적특 징은효과가매우우수한모핵구조인 kaempferin과달리세포막 투과성을저해할것으로생각된다. 즉, 이경우 kaempferitrin은 세포내보다는세포HH 존 재 는 단백질의활성조절을통한세 포내신호견달과정에 참여할가능성이 더클것으로사료된다. 이들가능성은추가적인관련연구를통해구체적으로확인할 예정이다. 그럼에도불구하31 현재까지의결과로볼때, kaempfer-
itrin에의한내재성면역조절은활성화된면역세포들이 갖는신
호전달현상■조절을통해매개되는것으로관단된다. .
결 론
YjtmiiJiibiscus cawwoi/nMs)에서추출한 kaempferitrin은대식 세포의 T cell 분열 및분화조절시 필수적인 co-stimulatory molecule의막표면발현을억제하였으며, CD29-매개성 세포•세 포간유착현상을효과적으로억제하였다. 그러나본화합물은식 세포작용, 라디칼소거능은매우미약한것으로나타났다. 세포- fibronectin간유착과정 및 CD80 발현에대한 kinase 저해제처 리효과등의결과를고려해볼때, kaempferitrin의면역약리작 용은세포표면 단백질의 활성억제보다는 NF-kB, Src, Akt 및
Syk를포함하는신호전달과정억제를통해진행되는것으로사
료된다. 향후보다자세한신호전달억제기견에관한추가연구 를 진행하고자한다.
감사의 글
표준 kaempferitrin을제공해주신 강원대학교약학대학권용
수 박사및농업생명과학대학김명조박사님께 감사드린다.
참고문헌
1) Tetley, T. D. : Inflammatory cells and chronic obstructive pulmonary disease. Curr. Drug. Targets. Inflamm. Allergy 4, 607 (2005).
2) Edwards, J. R, Zhang, X., Frauwirth, K. A. and Mosser, D. M. ; Biochemical and functional characterization of three activated macrophage populations. J. Leukoc. Biol 80, 1298 (2006).
3) Murtaugh, M. E and Foss, D. L. : Inflammatory cytokines and antigen presenting cell activation. Vet Immunol Immunopathol
87, 109 (2002).
4) Caulfield, J. J., Fernandez, M. H„ Sousa, A. R., Lane, S. J., Lee, T. H. and Hawrylowicz, C. ML : Regulation of major histocompatibility complex class II antigens on human alveolar macrophages by granulocyte-macrophage colony-stimulating factor in the presence of glucocorticoids. Immunology 98, 104
(1999).
5) Fang, H., Pengal, R. A., Cao, X., Ganesan, L. R, Wewers, M. D., Marsh, C. B. and Tridandapani, S .: Lipopolysaccharide- induced macrophage inflammatory response is regulated by SHIP J. Immunol 173, 360 (2004).
6) Chen, E and Shi, X .: NF-kappaB, a pivotal transcription factor in silica-induced diseases. Mol Cell Biochem. 234-235, 169 (2002).
7) Dijkstra, G., Moshage, H. and Jansen, R L. : Blockade of NF- kappaB activation and donation of nitric oxide: new treatment options in inflammatory bowel disease? Scand J. Gastroenterol Suppl 37 (2002).
8) Urgaonkar, S. and Shaw, J. X : Synthesis of kaempferitrin. J.
Org. Chem. 72, 4582 (2007).
9) Yokozawa, T, Dong, E., Kawai, Y., Gemba, M. and Shimizu, M ,:
Protective effects of some flavonoids on the renal cellular membrane. Exp. Toxicol Pathol 51, 9 (1999).
10) Jorge, A. R, Horst, H., de Sousa, E., Pizzolatti, M. G, and Silva, E R. : Insulinomimetic effects of kaempferitrin on glycaemia and on 14C-glucose uptake in rat soleus muscle. Chem. BioL Interact 149, 89 (2004).
11) de Sousa, E., Zanatta, L., Seifriz, L, Creczynski-PSsa, T. B., Pizzolatti, M. G., Szpoganicz, B. and Silva, E R. : Hypoglycemic effect and antioxidant potential of kaempferol-3,7-0-(alpha)- dirhamnoside from Bauhinia forficata leaves. J. Nat Prod. 67,
829 (2004).
12) da Silva, K. L , Biavatti, M. W, Leite, S. N., Yunes, R. A., Delle Monache, E and Cechinel Filho, V : Phytochemical and pharmacognositc investigation of Bauhinia forficata Link (Leguminosae). Z Naturforsch [C] 55, 478 (2000).
13) Fang, S. H., Rao, Y. K. and Tzeng, Y, M. : Inhibitory effects of flavonol glycosides from Cinnamomum osmophloeum on inflammatory mediators in LPS/IFN-gamma-activated murine macrophages. Bioorg. Med. Chem. 13, 2381 (2005).
14) Cho, J. Y., Baik, K. IL, Jung, J. H. and Park, M. H .: In vitro anti
inflammatory effects of cynaropicrin, a sesquiterpene lactone, from Saussurea lappa. Eur. J. Pharmacol 398, 399 (2000).
15) Lee, Y. G., Byeon, S. E., Kim, J. Y., Lee, J. Y., Rhee, M. H.,
활성화된 단핵구 및 대식세포의 항원제시기능에 대한 Kaempferitrin 외 조절 효과
Hong, S., Wu, J, C„ Lee, H. S., Kim, M. J., Cho, D. H. and Cho, J. Y.; Immunomodulatory effect of Hibiscus cannabinus extract on macrophage functions. J. Ethnopharmacol 113, 62 (2007).
16) Cho, J. Y., Fox, D. A,, Horejsi, V, Sagawa, K., Skubitz, K. M., Katz, D. R. and Chain, B .: The functional interactions between CD98, betal-integrins, and CD 147 in the induction of U937 homotypic aggregation. Blood 98, 374 (2001).
17) Kim, A. R., Cho, J. Y., Zou, Y., Choi, J. S. and Chung, H. Y. : Flavonoids differentially modulate nitric oxide production pathways in lipopolysaccharide-activated RAW264.7 cells.
Arch. Pharm. Res. 28, 297 (2005).
18) Burastero, S. E. and Rossi, G. A. : Immunomodulation by- interference with co-stimulatory molecules: therapeutic perspectives in asthma. Thorax 54, 554 (1999).
19) Marzio, R., Mauel, J. and Betz-Corradin, S. : CD69 and regulation of the immune function. Immunopharmacol ImmunotoxicoL 21, 565 (1999).
20) Kang, J. L.. Lee. H. W, Kim, H. J., Lee, H. S., Castranova, V, Lim, C. M. and Koh, Y. : Inhibition of SRC tyrosine kinases suppresses activation of nuclear factor-kappaB, and serine and tyrosine phosphorylation of IkappaB-alpha in lipopolysaccharide- stimulated raw 264.7 macrophages. J. Toxicol Environ. Health A 6 8, 1643 (2005).
21) Savina, A. and Amigorena, S. ; Phagocytosis and antigen presentation in dendritic cells. Immunol. Rev. 219, 143 (2007).
22) Stonehouse, T. J., Woodhead, V E., Herridge, P. S., Ashrafian, H., George, M., Chain, B. M. and Katz, D. R, : Molecular characterization of U937-dependent T-cell co-stimulation.
Immunology 96, 35 (1999).
23) Woodhead, V E., Stonehouse, T J., Binks, M. H., Speidel, K., Fox, D. A., Gaya, A., Hardie, D., Henniker, A. J., Horejsi, V, Sagawa, K., Skubitz, K. M., Taskov, H., Todd, R. E, 3rd, van Agthoven, A., Katz, D. R. and Chain, B. M. : Novel molecular mechanisms of dendritic cell-induced T cell activation. Int.
Immunol 12, 1051 (2000).
24) Longhurst, C. M. and Jennings, L. K. : Integrin-mediated signal transduction. Cell. Mol. Life Sci. 54, 514 (1998).
25) Migliaccio, E., Giorgio, M. and Pelicci, R G. : Apoptosis and aging: role of p6 6Shc redox protein. Antioxid. Redox. Signal. 8,
600 (2006).
26) Katiyar, S. K., Matsui, M. S., Elmets, C. A. and Mukhtar, H. : Polyphenolic antioxidant (-)-epigallocatechin-3-gallate from green tea reduces UVB-induced inflammatory responses and infiltration of leukocytes in human skin. Photochem. PhotobioL
69, 148 (1999).
Vol. 51, No. 6, 2007

![Fig. 5 - Effect of kaempferitrin on CD29-mediated cell-cell adhesion. U937 cells were incubated with kaempferitrin in the presence or absence of pro-aggregative (activating) antibodies [antibodies (1 each) to CD29 (MEM lOlA) and CD43 (161-46)] for 2 h](https://thumb-ap.123doks.com/thumbv2/123dokinfo/5022565.308606/5.884.84.809.97.466/kaempferitrin-mediated-incubated-kaempferitrin-aggregative-activating-antibodies-antibodies.webp)