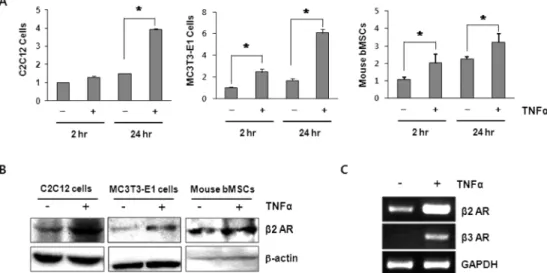
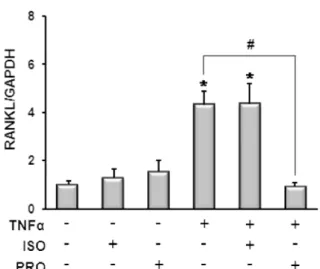
$TNF{\alpha}$ Increases the Expression of ${\beta}2$ Adrenergic Receptors in Osteoblasts
6
0
0
전체 글
(2)
(3)
(4)
(5)
(6)
수치

관련 문서
Primary hepatocytes were incubated with T090 for 24 h following Res pretreatment (1 h). Cells were fixed and stained with Oil red O and observed under a microscope. The Oil
The proliferations of MC3T3-E1 cell seeded on pristine PCL scaffold and surface modified scaffolds for 1, 3, and 5 days... ALP activity of the MC3T3-E1 cell seeded
There was a significant increase in new bone formation in the group in which toothash and plaster of Paris and either PRP or fibrin sealants were used, compared with the groups
mRNA expression of osteonectin, Runx2 and BSP(bone sialoprotein) in MC3T3-E1 cells cultured for 24 hours on 4 different titanium surfaces.. The mRNA were analyzed
MC3T3-E1 cell morphorologies on the (a) CP-Ti surface and (b) nano-mesh surface for 20 min culturing time.. MC3T3-E1 cell morphorologies on the (a) CP-Ti surface and
The untreated surfaces and AA plasma treated with 3D-PCL surfaces were also examined for their in vitro pre-osteoblast (MC3T3-E1) cells proliferation and
In the case of inherently conducting polymers like PANI or PPY, the conductivity values of the metal oxide based composites showed wide variations excepting in PPY-ZrO 2 , and
In other words, pomegranate extract has an effect of inhibiting the progression of alveolar bone loss due to periodontal inflammation by reducing the expression of COX-1 and
