Introduction Introduction
Mesenchymal stem cells (MSCs) are adult stem cells residing in the organ-specific supporting tissue and regarded to con- tribute to the organ repair from the injury [1,2]. MSCs display the differentiation potential toward several lineages including adipocytes, osteoblasts, chondrocytes, and organ-specific adult cell types [3]. Dental pulp stem cells (DPSCs) are MSCs residing in dental pulp and can be differentiated to odontoblast and restore the damaged dentin [4,5]. Though the recent lin- eage tracing experiments and the single cell analysis revealed the developmental origin and the constituting cell types of dental pulp [6-8], the exact nature of DPSCs still remains elu-
sive.
Organoid culture has been replacing the sphere culture, both of which have provided the platform for the long-term cul- ture of adult stem cells [9]. In the sphere culture, serum-free minimal condition drives the formation of three-dimensional (3D) cell aggregates in the suspension resulting in the gradual increment of stem cell population during serial passaging whereas the heterogeneity and cellular arrangement of in vivo origin are well-preserved in the organoid culture [10]. Organoid is advantageous in studying the morphogenesis and the loca- tion of adult stem cells, but sphere culture provides the edge in enriching the stem cell pool for the further analysis [11]. As the major types of organoids have been derived from epithelial Int J Oral Biol 45:197-203, 2020
pISSN: 1226-7155 • eISSN: 2287-6618 https://doi.org/10.11620/IJOB.2020.45.4.197
Increased SOX2 expression in three-dimensional sphere culture of dental pulp stem cells
Eun Jin Seo
1and Il Ho Jang
1,2*
1
Dental and Life Science Institute, Pusan National University School of Dentistry, Yangsan 50612, Republic of Korea
2
Department of Oral Biochemistry, Pusan National University School of Dentistry, Yangsan 50612, Republic of Korea
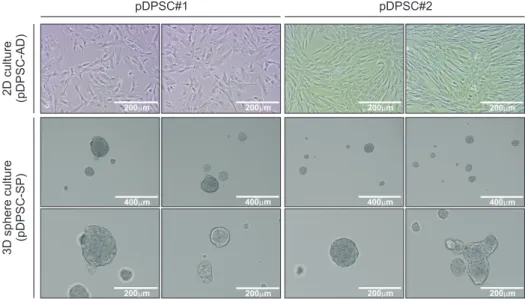
Mesenchymal stem cells in the dental pulp exhibit a tendency for differentiation into various dental lineages and hold great potential as a major conduit for regenerative treatment in dentistry. Although they can be readily isolated from teeth, the exact characteristics of these stem cells have not been fully understood so far. When compared to two- dimensional (2D) cultures, three-dimensional (3D) cultures have the advantage of enriching the stem cell population.
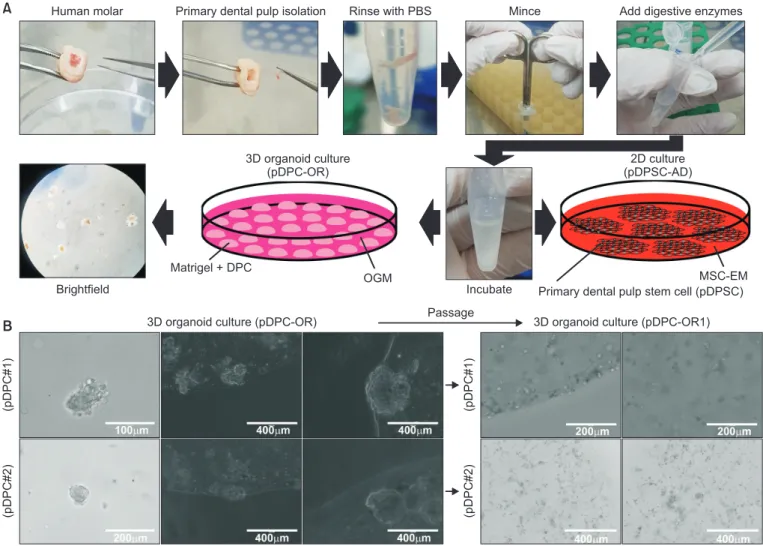
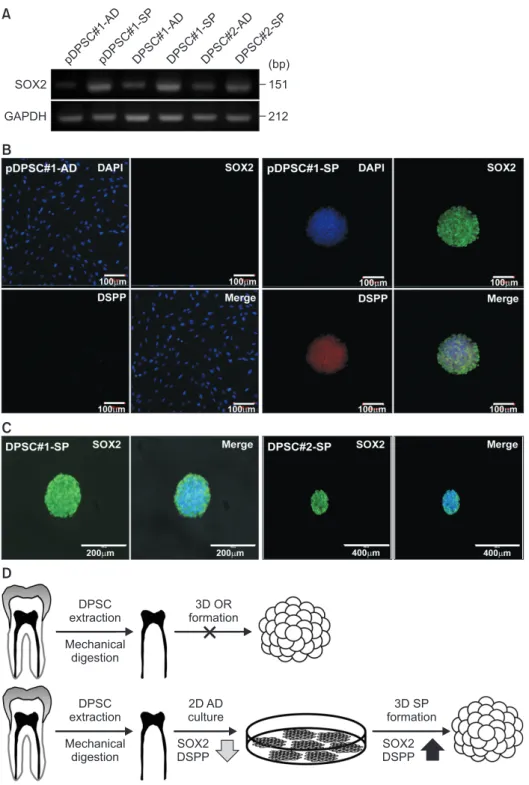
Hence, 3D-organoid culture and 3D-sphere culture were applied to dental pulp cells in the current study. Although the establishment of the organoid culture proved unsuccessful, the 3D-sphere culture readily initiated the stable generation of cell aggregates, which continued to grow and could be passaged to the second round. Interestingly, a significant increase in SOX2 expression was detected in the 3D-spheroid culture compared to the 2D culture. These results indicate the enrichment of the stemness-high population in the 3D-sphere culture. Thus, 3D-sphere culture may act as a link between the conventional and 3D-organoid cultures and aid in understanding the characteristics of dental pulp stem cells.
Keywords: Dental pulp stem cell, SOX2, Three-dimensional culture, Sphere, Organoids
Received November 30, 2020; Revised December 15, 2020; Accepted December 15, 2020
*Correspondence to: Il Ho Jang, E-mail: [email protected] https://orcid.org/0000-0002-0820-3035 Copyright © The Korean Academy of Oral Biology
CC