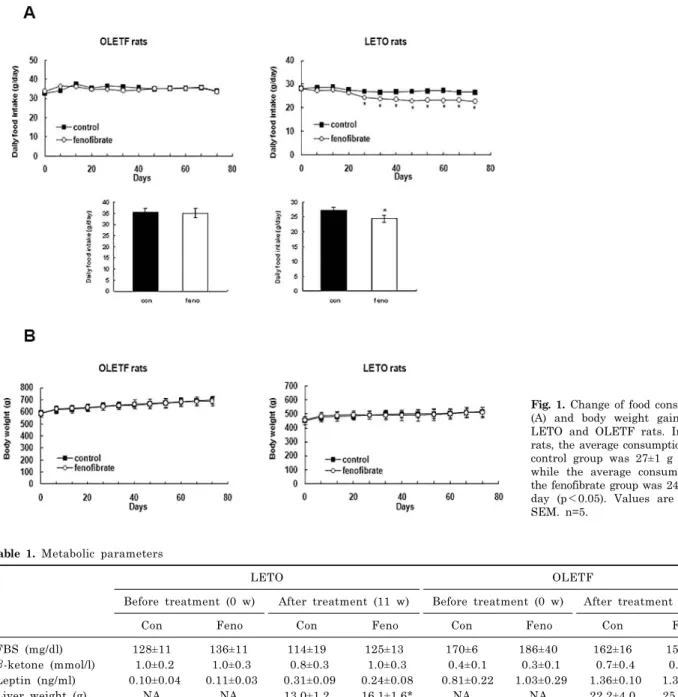
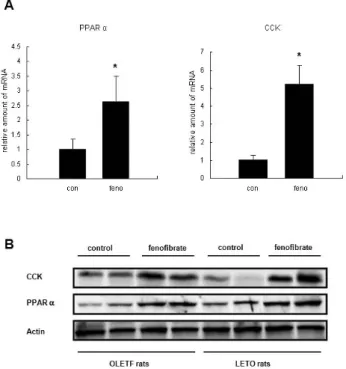
Reduction of Food Intake by Fenofibrate is Associated with Cholecystokinin Release in Long-Evans Tokushima Rats
6
0
0
전체 글
(2)
(3)
(4)
(5)
(6)
수치
관련 문서
In this study, it is shows that the stream of CO 2 is more effective in the reduction of unreacted TEGDMA and the increase of surface microhardness than that of N 2
Not only indigenous Korean food culture formed its traditional way in the latter part of the Chosun Dynasty, but also was the transition period from
Effect of caloric restriction on the expression of PGC-1 and PPARs mRNA in liver of Otsuka Long-Evans Tokushima Fatty Rats.. Long-lived growth hormone
Mean arterial plasma concentration-time profiles of 4-hydroxytamoxifen after an oral administration of tamoxifen (10 mg/kg) to rats in the presence or absence
Probiotic properties of lactic acid bacteria isolated from mukeunji, a long-term ripened kimchi. Journal of Food
영어 : Research on Recognition of Food Additives Contained in Processed Food by Elementary Students
Effect of mulberry leaf powder containing the lactic acid bacteria on body weight gain, food intake and food efficiency ratio in a constipation model
The conventional method of long distance face recognition uses reconstruction method turning face images of long distance with low resolution into
