(40205) 경북 울릉군 북면 현포2길 127
4제주대학교 해양과학대학 해양의생명과학부 (63243) 제주특별자치도 제주시 제주대학로 102
Study on the Annual Reproductive Cycle of the Comb Pen Shell, Atrina pectinata (Linnaeus, 1767) (Mollusca: Bivalvia: Pinnidae)
on the Southern Coast of Ulleungdo Island, Korea
Hyun-Sung Yang
1*, Choong Hwan Noh
2, Sung Jin Yoon
3, Yun-Bae Kim
3, Kwang-Sik Choi
4, and Do-Hyung Kang
11Jeju International Marine Science Center for Research and Education, KIOST Jeju 63349, Korea
2East Sea Environment Research Center, KIOST Uljin 36315, Korea
3Ulleungdo-Dokdo Ocean Science Station, KIOST Ulleung 40205, Korea
4School of Marine Biomedical Science (BK21 PLUS), Jeju National University Jeju 63243, Korea
Abstract : Gametogenesis of the comb pen shell, Atrina pectinata (Linnaeus, 1767) (Bivalvia: Pinnidae) on the southern coast of Ulleungdo Island, Korea was assessed monthly (November 2013 to October 2014) using histology. Gametogenesis commenced in January when the surface water temperature was 12.6oC and pen shells evidenced an early development phase with small oogonia from January to April, although few females exhibited ripe eggs in their follicular epithelium. In April, the oocyte diameter increased rapidly, and fully mature eggs appeared in May. First spawning males and females were observed in June as the surface water temperature reached 19.3oC and July (23.2oC) respectively. The spawning activity continued until the end of September. Histology indicated that the spawning peak of the females in Ulleungdo Island was July to August. During October to January, most of the pen shells were in spent and resting stages. Our data suggested that A. pectinata is a summer spawner, and their annual gametogenesis is closely associated with the seasonal variation in the surface water temperature. The present study is the first provided fundamental information on the life history of A. pectinata in Ulleungdo Island, and this can be put to good use in the management of this pen shell in the study area.
Key words : gametogenesis, spawning, comb pen shell, Atrina pectinata, Ulleungdo Island
*Corresponding author. E-mail : [email protected]
1. 서 론
키조개(Atrina pectinata)는 해양이매패류 중 크기가 가 장 큰 여과섭식자 중 하나이며, 태평양, 인도양 및 대서양 의 얕은 조하대의 부드러운 저질에 서식하는 것으로 알려 져 있다. Schultz and Huber (2013)에 따르면, 전세계 온 대와 열대 지역에서 55종의 키조개과에 속하는 종들이 서 식하고 있으며, 우리나라에서는 5종의 키조개과에 속하는 종들이 보고된 반면, 일본과 중국에서는 각각 14종과 9종 이 서식하는 것으로 보고되었다(NFRDI 1997, 2009;
Okutani 2000; Min 2004; Zhongyan 2004). 국내에 알려 진 키조개(A. pectinata)는 남서해안의 수심 20−50 m 사 니질로 구성된 조하대에 주로 서식하는 것으로 보고되었 다(Kim and Hur 1998; Hong et al. 2002; Min 2004; An et al. 2012). 키조개는 상업적으로 매우 중요한 종 중 하나 이며, 키조개의 패주는 우리나라를 비롯한 아시아 여러 국 가에서 가장 인기있는 수산물 중 하나이다. 키조개는 남해 안(가막만, 여자만, 득량만 및 진해만)과 서해안(천수만, 보령 및 군산 등지)이 주요 생산 지역으로 알려져 있다.
통계청 국가통계포털 수산 통계자료에 따르면, 국내 키조 개 생산량은 1990년 15,300톤이 생산되던 것이 1996년 837톤으로 급감하고, 이후 매년 생산량이 증가와 감소를 반복하는 경향을 보이고 있는데(Statistics Korea 2015), Ryu et al. (2001)은 서식지 감소와 남획을 이 같은 현상의 원인으로 꼽고 있다.
울릉도(37o45'−37o55'N, 130o80'−130o95'E)는 우리나라 동해안에서 120 km 떨어져 있고 화산 활동에 의해 생성 된 섬이다. 울릉도의 해양환경은 주변해역 대부분이 조간 대부터 조하대까지 가파른 암반 절벽으로 이루어져 있으 며, 연중 높은 파도의 영향을 받는 것으로 알려져 있다. 이 러한 독특한 지형학적 특징에도 불구하고, 울릉도 주변 해 역에 서식하는 해양생물에 대한 연구는 많지 않다. 2013년 울릉군 보고에 따르면, 울릉도 남쪽 해역 수중 조사를 실 시한 결과, 낮은 수심의 조하대에서 높은 종다양성의 식물 상과 동물상을 보고한 바 있으며(Ulleung-Gun 2013), 이 조사에서 수심 25−30 m에 서식하는 키조개(A. pectinata) 군락이 발견되었다(Yang et al. 2015). Choe et al. (1994) 이 1989년부터 1992년까지 울릉도 주변 해역에 서식하는 연체동물을 조사한 바에 따르면, 울릉도에는 총 33종의 이매패류가 서식하는 것으로 보고하였다. 키조개(A.
pectinata)가 이전 조사에서 발견되지 않았는데, Yang et al. (2015)에 따르면 울릉도 남쪽해역의 키조개 발견 지역 이 수심이 깊고 접근하기 어려워 이전 수중 조사활동에서 관찰되지 않았던 것으로 보여진다. 이번 키조개 개체군 발 견에 따라 울릉군에서는 수중조사 해역을 더 확대할 예정 이며, 발견된 해양생물들에 대한 개체군 보호 방안을 마련
중인 것으로 알려졌다.
해양 이매패류의 번식주기를 이해하는 것은 이들의 생 산 관리 및 수산 양식 발전에 있어 가장 중요한 일 중 하 나이다(Menzel 1991; Gosling 2003). 해양 이매패류의 번 식은 생식세포 생성, 생식소 성장, 산란과 수정 그리고 유 생 발달 등으로 구분된다(Mackie 1984; Olive 1985). 이 와 같은 번식과정은 내부적으로는 유전적 또는 진화적인 요소들에 의해 조절되고, 외부적으로는 수온, 염분, 가용 먹이 등의 환경적 요소들에 의해 결정된다(Hofmann et al.
1992; Kang et al. 2000; Gosling 2003; Drummond et al.
2006). 일반적으로 해양 이매패류의 배우자형성 과정은 조 직학적 방법을 이용하여 생식소 성숙 정도, 난모세포 크기 및 이미지 분석 프로그램을 활용한 여포 내 알 또는 정자 면적을 측정하여 나타낸다(Heffernan and Walker 1989;
Kang et al. 2003, 2007; Fabioux et al. 2005).
국내에서 키조개(A. pectinata)와 관련된 연구들을 살펴 보면, 생식소 발달에 따른 번식 주기(Chung et al. 2006, 2012; Lee et al. 2006), 부위별 생화학 조성분 분석(Baik et al. 2001; Lee et al. 2015), DNA 마커를 이용한 자연산 키조개와 양식산 키조개 유전자 비교(An et al. 2011), 먹 이에 따른 성장 및 성숙유도(Moon et al. 2009), 밀도에 따른 성장과 생존율(Kim et al. 2008), 자연채묘에 관한 연구(Son et al. 2005) 등이 이뤄졌다. 그러나 앞서 언급한 연구들은 남·서해안에 서식하는 개체군에 대한 것인 반면 금번 연구는 동해안 북부 해역에 서식하는 키조개 군락의 번식주기에 관한 최초의 연구로 이 지역 키조개의 배우자 형성 과정 및 번식 주기를 조사함으로써 울릉도 해역 키 조개의 생활사를 이해하며 타 지역과 비교를 통한 번식학 적 특성을 이해하기 위하여 수행되었다.
2. 재료 및 방법
채집 지역 및 시료 채집
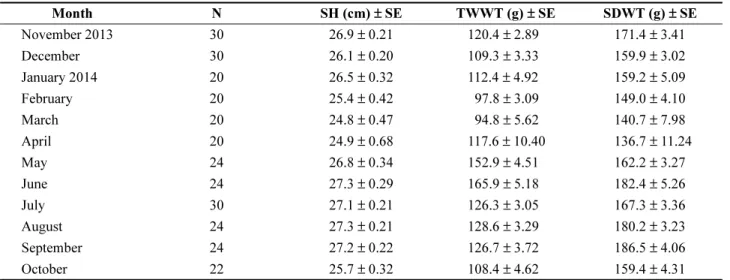
실험에 사용된 키조개는 울릉도 남쪽 수심 25−30 m 해 역(37o45'N, 130o85'E)(Fig. 1)에서 스쿠버 다이빙을 통해 2013년 11월부터 2014년 10월까지 매달 20−30 개체씩 채집하였다(Table 1). 키조개는 저질 바닥이 사질로 구성 된 약 600 × 700 m의 평평한 지역에 평균 11개체/m2의 밀 도로 분포하고 있었으며, 주변에는 키조개 은신처 역할의 잘피(Zostera caulescens)가 함께 서식하고 있었다(Yang et al. 2015). 채집한 키조개는 즉시 실험실로 옮겨 각 개체 별로 각장(Shell Length, SL), 각폭(Shell Width, SW) 및 각고(Shell Height, SH)를 측정한 후, 육질부와 패각으로 분리하였다. 분리한 육질부는 다시 내장낭이 포함된 생식 소, 폐각근 그리고 잔여조직의 세 부분으로 분리하여 각각 의 습중량(Tissue Wet Weight, TWWT)을 측정하였다. 비
만도[Condition Index, CI = 총 건중량(g)/건조 패각무게 (g)](Walne 1976; Mann and Glomb 1978) 및 생식소 중량 지수[Gonadosomatic Index, GSI = 생식소 건중량(g)/건조 패각무게(g)](Lee et al. 2015)를 산출하기 위해 동결건조 후 건중량을 재측정 하였다. 키조개 패각 또한 비만도와 생식소 중량지수를 산정하기 위해 실온에서 건조하였고, 이 연구에 사용된 키조개의 월 평균 각 체조직 습중량 및 패각 건중량 측정 자료는 Table 1과 같다.
조사 지역의 연중 수온 및 염분 데이터는 국립해양조사 원(Korea Hydrographic and Oceanographic Administration, KHOA)의 실시간 해양관측 자료를 활용하였다. 채집지역 과 가장 가까운 울릉도 관측소 자료를 이용하였고, 일별 측정 자료를 월별 평균값으로 계산해 사용하였다. 연중 수 Fig. 1. Map showing the sampling location
Table 1. Biometric measurements of sampled individuals of the Atrina pectinata population in this study. N, number of pen shell analyzed, SE, standard error, SH, shell height, TWWT, tissue wet weight in gram, SDWT, shell dry weight in gram
Month N SH (cm)± SE TWWT (g)± SE SDWT (g)± SE
November 2013 30 26.9± 0.21 120.4± 2.89 171.4± 3.41
December 30 26.1± 0.20 109.3± 3.33 159.9± 3.02
January 2014 20 26.5± 0.32 112.4± 4.92 159.2± 5.09
February 20 25.4± 0.42 097.8± 3.09 149.0± 4.10
March 20 24.8± 0.47 094.8± 5.62 140.7± 7.98
April 20 24.9± 0.68 117.6± 10.40 136.7± 11.24
May 24 26.8± 0.34 152.9± 4.51 162.2± 3.27
June 24 27.3± 0.29 165.9± 5.18 182.4± 5.26
July 30 27.1± 0.21 126.3± 3.05 167.3± 3.36
August 24 27.3± 0.21 128.6± 3.29 180.2± 3.23
September 24 27.2± 0.22 126.7± 3.72 186.5± 4.06
October 22 25.7± 0.32 108.4± 4.62 159.4± 4.31
Fig. 2. Seasonal changes in the surface water temperature and salinity at the sampling site from November 2013 to October 2014
온 및 염분 변화 범위는 각각 10.2−23.6oC, 32.2−33.7 psu 로 나타났다(Fig. 2).
조직슬라이드 제작 및 생식소 관찰
키조개 조직으로부터 분리된 생식소의 무게 측정이 끝 난 뒤, 생식소를 포함하는 2 mm 두께의 조직 단편을 절취 하여, Davidson’s 고정액에 48시간 고정하였다. 이 후 에 탄올을 이용하여 탈수 과정을 거친 후 파라핀으로 포매하 였고, 포매된 조직 시료는 마이크로톰을 이용하여 6 μm 두께로 박절한 후, Harris’ hematoxylin과 Eosin Y로 비 교·염색하였다. 키조개의 생식소 발달단계는 Lee et al.
(2006)의 방법을 일부 수정하여 휴지기(Resting stage), 초기발달기(Early developing stage), 후기발달기(Late developing stage), 완숙기(Ripe stage), 부분산란기(Partially spawned stage)와 산란후기(Spent stage)의 총 6단계로 구 분하였다. 난경(Oocyte Diameter, OD)은 이미지 분석 프 로그램(Image Pro-Plus Version 2.0, Media Cybernetics, USA)을 이용하여 측정하였다. 각 개체당 5곳을 임의로 지 정하여 여포의 디지털 이미지를 확보하고 여포 내 난 크 기를 계측하여 월별 분포양상 및 평균값을 산출하였다.
3. 결 과
키조개의 각고, 습중량 및 패각 건중량
이번 연구에 사용한 키조개의 월 평균 각고(SH), 습중 량(TWWT) 및 패각 건중량(SDWT)을 표준오차(Standard Error, SE)와 함께 Table 1에 정리하였다. 월 평균 각고 24.8± 0.47−27.3 ± 0.29 cm의 개체들이 이번 연구에 사용 되었다. 조직 습중량과 패각 건중량은 각각 94.8 ± 5.6−
165.9± 5.2 g과 136.7 ± 11.2−186.5 ± 4.1 g의 범위였다. 습 중량은 2014년 5월과 6월 사이에 급격히 증가하였고, 7월 부터 급격히 감소하는 것을 확인할 수 있었다.
생식소중량지수(GSI)와 비만도(CI)의 월 변화
울릉도 키조개의 생식소중량지수와 비만도의 월 변화 양상을 Fig. 3에 나타내었다. 생식소중량지수와 비만도는 각각 9.9−35.1 및 11.9−18.3의 범위였다. 생식소중량지수 는 2014년 1월(10.6)부터 서서히 증가하기 시작해 6월 (35.1) 최고값을 나타내었고, 7월(20.4)부터 8월(17.7)까지 급격히 감소한 뒤 지속적으로 낮아지는 추세를 보였다. 비 만도 역시 생식소중량지수와 유사한 변화 양상을 보이고 있었다. 2014년 3월(13.4)부터 5월(18.3)까지 급격히 증가 하는 추세를 보인 후, 6월(17.5)에서 8월(12.3) 사이에 급 격히 낮아졌고 이후 연중 가장 낮은 비만도를 나타냈다.
이와 같이 생식소중량지수와 비만도의 증가와 감소는 조 직학적 관찰 결과, 생식소 발달 및 산란 패턴과도 밀접한
관련이 있었다.
생식소 발달 변화
Fig. 4와 5는 울릉도 키조개의 수컷 생식소와 암컷 생식 소 발달 변화를 나타낸 그림이다. 휴지기 상태의 키조개 조직에서는 암·수 모두 생식세포가 관찰되지 않았고, 여포 발달도 안되어 있는 상태를 보이고 있었다(Fig. 4A, Fig.
5A). 초기 발달기 상태의 키조개 조직에서는 수컷의 경우, 정원세포(Spermatogonia, SG)가 여포를 가득 채우고 있었 고, 일부 여포에서는 발달한 정모세포(Spermatocyte, SC) 들이 여포 한 가운데 자리하고 있었다(Fig. 4B). 암컷 키 조개 조직에서는 여포가 점차 확장되고 있는 것을 확인할 수 있었고, 확장되고 있는 여포벽을 따라 약 10 μm 크기 의 난원세포(Oogonia, OG)들이 분포하고 있는 것이 관찰 되었다(Fig. 5B). 후기 발달기로 접어 들면서 키조개의 암·수 생식소는 급격히 발달하는 것을 확인할 수 있었다.
여포의 면적은 초기 발달기에 비해 넓어졌으며, 암·수 생 식소 역시 여포를 가득 채워가고 있었다. 수컷 조직에서는 정원세포(SG), 정모세포(SC), 정자(Spermatozoa, SZ)가 여포 바깥에서부터 차례로 자리잡고 있는 것을 관찰할 수 있었고(Fig. 4C), 암컷 조직에서는 초기 발달기에 비해 좀 더 성숙한 난원세포(OG)들이 여포 안쪽 벽을 따라 자리하 고 있었으며, 일부 난황형성난포세포(Vitellogenic Oocyte, VO)가 함께 관찰되었다(Fig. 5C). 완숙기에 접어 들면서 여포는 생식소 발달 기간 중 가장 크게 확장되었다. 수컷 조직에서는 확장된 여포 안쪽에 성숙한 정자(SZ)가 가득 채워져 있었고, 여포 외벽을 따라 정원세포(SG)와 정모세 포(SC)가 감싸 안은 형태를 보이고 있었다(Fig. 4D). 암컷 조직에서도 팽창된 여포 내에 완숙한 알(Mature Oocyte, MO)들이 가득 자리 잡은 것을 확인할 수 있었으며, 일부 발달중인 난원세포(OG)들도 여포 바깥부분에서 관찰되었 다(Fig. 5D). 부분 산란기 상태의 생식소는 방정 및 산란 으로 인한 여포 내 빈 공간이 관찰되었다. 수컷 생식소는 Fig. 3. Seasonal variations in gonadosomatic index (GSI)
and condition index (CI) of Atrina pectinata
방정으로 인해 빈공간이 관찰되고 방출되지 않은 정자들 이 잔존하고 있는 것을 관찰할 수 있었다(Fig. 4E). 암컷 생식소 또한 산란으로 인해 여포 내 빈 공간과 더불어 일 부 성숙난들이 남아 있는 것을 확인할 수 있었다(Fig. 5E).
산란 후기 상태의 암·수 생식소는 여포의 크기가 급격히 줄어 들었으며, 줄어든 여포 안에 방정 후 남은 여분의 정 자(Relict Spermatozoa, RSZ)(Fig. 4F) 및 산란 후 흡수되 는 알과 잔존난(Relict Oocyte, RO)이 관찰되었다(Fig. 5F).
연중번식주기
울릉도에 서식하는 키조개의 연중번식주기 패턴을 Fig.
6(A-male, B-female)에 나타내었다. 수컷과 암컷 키조개의 생식소는 1월 수온 12.6oC일 때 초기생식세포가 처음 만 들어지는 것으로 판단되며, 이때 초기발달단계의 생식소 비율은 수컷과 암컷 각각 28.6%와 37.5%였다. 후기 발달 기 상태의 생식소는 수컷의 경우, 2월부터 관찰되기 시작 하여 6월까지 나타난 반면, 암컷은 4월과 6월 사이에만 관 찰되었다. 이때는 이 지역의 수온이 서서히 증가하는 기간 으로 수온 범위는 10.2−19.3oC였다. 암수 모두 성숙한 생 식소는 6월 각각 66.7%와 73.3%로 나타났고, 이 시기의 생식소는 방정과 산란이 임박했음을 유추해 볼 수 있었 다. 방정과 산란은 수컷과 암컷 키조개 모두 7월부터 동시 Fig. 4. Photomicrographs of testis. (A) Resting stage: not observed any sperm in the follicle. (B) Early developing
stage: spermatogonia (SG) fill in the follicle. (C) Late developing stage: columns of spermatogonia (SG), spermatocyte (SC) and spermatozoa (SZ) move to the central part of growing testes. (D) Ripe stage: mature testes filled with spermatozoa (SZ). (E) Partially spawned stage: partly spawned testes with vacated space. (F) Spent stage: partly devoid follicle with relict spermatozoa (RSZ). Scale bar: 50 µm
에 일어나는 것을 확인할 수 있었고, 9월까지 지속되었 다. 이 기간 동안, 8월부터 9월까지 수컷은 73.7−100%의 방정을 하였고, 암컷은 57.1−76.9%의 산란이 이루어졌으 며, 수온은 22.9−23.6oC의 범위로 연구 기간 중 가장 높은 시기였다. 수온이 20.7oC로 낮아지는 10월부터 수컷과 암 컷의 생식소는 거의 대부분 산란후기와 휴지기 상태를 보 였으며, 이러한 생식소 상태는 12월까지 지속되는 것을 관찰하였다.
월 별 난경 변화 패턴
암컷 키조개의 난경 변화 양상을 Fig. 7에 나타내었고, 월 평균 난경의 크기는 11.6−54.5(Nov−Jun) μm 범위였
다. 초기 발달기 상태가 처음 관찰된 1월은 평균 13.4 μm 로, 대부분 20 μm 미만의 난들이 90% 이상을 차지하고 있었다. 후기 발달기 상태가 관찰되기 시작한 4월 평균 난경 크기는 28.0 μm로 증가하였고, 20−40 μm 크기의 난들이 약 90% 정도 분포하고 있었다. 이후 평균 난경 크 기는 급격히 증가하기 시작하였으며, 대부분 완숙기 상태 의 개체들이 관찰된 6월 평균 난경은 54.5 μm로 연구기 간 중 가장 높게 나타났다. 이 시기의 난들은 10−100 μm 크기 범위에서 다양하게 관찰되었으나, 40−70 μm 크기의 난들이 약 90% 정도 차지하고 있었다. 8월 산란 후기의 평균 난 크기는 34.6 μm로 감소하였다가, 9월 다시 54.5 μm로 증가한 후, 10월 28.6 μm로 낮아졌다. 9월 평 Fig. 5. Photomicrographs of ovaries. (A) Resting stage: not observed any gonad in the follicle. (B) Early developing
stage: ovary containing small oogonia (OG) and early vitellogenic oocytes occur along the follicle wall. (C) Late developing stage: growing ovary with vitellogenic oocytes (VO). (D) Ripe stage: mature ovaries (MO) packed with ovaries. (E) Partially spawned stage: partly spawned ovaries with loosely packed ova and vacated space.
(F) Spent stage: relict oocyte (RO) and empty space showed in the follicle. Scale bar: 50 µm
균 난경 크기가 증가한 이유는 8월에 잔존했던 30−50 μm 크기의 난들이 산란을 위해 다시 성숙했기 때문이라 사료 된다.
4. 고 찰
이번 연구에서 확인된 울릉도 키조개의 생식소중량지수 (GSI) 및 비만도(CI)의 연중변화 양상은 번식패턴과 일치 하는 경향을 보이고 있다. 급격히 감소하는 생식소중량지 수는 키조개의 산란을 의미하며, 2014년 6월 35.1%의 최 고치를 보인 후 급격히 감소하는데, 이로 미루어 보아 울
릉도 지역에 서식하는 키조개는 6월부터 8월 사이 연중 1회 산란하는 종으로 파악된다(Fig. 3). Lee et al. (2015) 은 남해 득량만에 서식하는 키조개의 산란 주기를 관찰한 결과, 이 지역에 서식하는 키조개의 생식소중량지수 최고 치는 2010년과 2011년 각각 30.6%(7월)와 32.6%(5월)를 보인다고 보고하였다. 다른 연구에서도 마찬가지로 다양 한 이매패류들이 산란 직전 생식소중량지수가 최고치를 보인 후 산란과 함께 감소하는 경향을 나타내었다. 남해에 서식하는 참굴(Crassostrea gigas)의 경우, 생식소무게가 전체 건중량의 40% 전후가 되면 산란이 일어난다고 알려 져 있다(Kang et al. 2003). Choi et al. (1993)은 미국 텍 Fig. 6. Percentage of Atrina pectinata in various reproductive stages over 12 months. (A) male comb pen shell, (B) female
comb pen shell
사스 Galveston Bay에 서식하는 굴(C. virginica)의 번식 량을 측정한 결과, 생식소 중량이 굴 전체 무게의 20%가 되면 산란이 시작된다고 보고하였다. Park and Choi (2004)와 Uddin et al. (2012)은 바지락의 생식소중량을 효 소면역학적방법을 이용해 측정한 결과, 바지락 생식소가 전체 중량의 25% 전후가 되면 산란이 일어난다는 연구결 과를 발표한 바 있다. Park et al. (2005)은 남해 거제에서 채집된 개조개(Saxidomus purpuratus) 알에 대한 항체를 개발해 생식소중량지수를 측정한 결과, 개조개의 산란은 생식소가 전체 무게의 27%를 차지할 때 일어난다고 보고 하였다. 따라서, 이번 연구결과와 남해안에 서식하는 키조 개의 생식소중량지수를 비교해 볼 때, 키조개는 생식소무 게가 전체 무게 중 30−35%를 차지할 때 산란이 일어난다 고 볼 수 있다.
다양한 해양 이매패류 연구에서 비만도(CI)는 생식소 발달 및 산란과 밀접한 연관이 있음을 발표한 바 있다 (Kang et al. 2007; Joaquim et al. 2008; Kim et al. 2010;
Limpanont et al. 2010). 일반적으로 해양 이매패류의 연중 최고 비만도는 생식소가 완숙 상태인 산란 직전인 것으로 알려져 있고, 비만도가 가장 낮은 시기는 산란 후기로서 비만도는 생식소 성숙의 연중 변화를 가늠하는데 유용하 게 사용되어 진다(Kim et al. 2005; Moura et al. 2008).
이번 연구에서 울릉도 남쪽에 서식하는 키조개의 비만도 는 11.9−18.3의 범위를 보이며, 2014년 5월 18.3으로 최고 치를 보인 후 7월 급격히 감소하는 것을 확인할 수 있었 다(Fig. 3). 비만도가 급격히 감소하는 7월과 8월, 조직관 찰 결과와 비교해 보면 이 시기의 키조개는 대부분 방정 과 산란이 일어나는 것이 확인되었다. Lee et al. (2015)의 동일 방법으로 산정한 연구결과를 보면, 득량만에 서식하 는 키조개의 비만도는 10−25의 범위를 나타내었고, 산란 직전 최고치를 보인 후 감소하는 패턴이 이번 연구 결과 와 유사하였다. 다른 이매패류에서도 유사한 연구 결과들 이 보고되었다. Uddin et al. (2010)은 선재도에 서식하는 바지락(R. philippinarum)의 연중 비만도 관찰 결과를 발 표하였다. 이 지역에 서식하는 바지락은 수온이 낮고 가용 먹이원이 적은 2−3월 사이 가장 낮은 비만도를 나타내다 가 서서히 증가하여 산란 직전 가장 높은 값을 보인 후 다 시 낮아진다고 보고하였다. 포르투갈에 서식하는 이매패 류인 Spisula solida에서도 유사한 현상이 관찰되었다.
Gaspar and Monteiro (1999)는 S. solida의 비만도 변화를 연중 관찰한 결과, 생식소 발달 주기와 밀접한 관련이 있 으며 산란 직전 생식소가 발달하는 시기에 증가하고 산란 기에 감소한다고 보고한 바 있다. 득량만에 서식하는 키조 개의 비만도는 10−25의 범위를 보였는데(Lee et al.
2015), 이번 연구에서 나타난 결과(11.9−18.3) 보다 높은 수치이다. Baek et al. (2014)은 곰소만에 서식하는 바지락
의 비만도를 관찰한 결과, 이들의 먹이원인 chlorophyll 함 량이 높은 만 입구에 서식하는 바지락의 비만도가 상대적 으로 chlorophyll 함량이 낮은 만 안쪽에 서식하는 바지락 보다 높게 나타나는 것을 보고하였다. 이와 같이 울릉도 지역의 키조개에서 낮은 비만도를 보이는 이유는 이 지역 의 먹이 조건이 득량만 보다 낮은 것으로 사료된다.
수온, 염분 및 가용 먹이와 같은 환경 인자는 해양 이매 패류의 성장, 배우자형성 및 체내 생화학 조성분 함량 등 에 영향을 미친다(Giese 1959; Okumus and Stirling 1998; Saxby 2002). 또한, 이와 같은 해양환경인자의 변화 는 해양 이매패류의 산란을 유발할 수 있는 물리적 환경 요소로 고려되어 진다(Mann 1979a; Dohmen 1985). 이 중에서도 수온은 해양 이매패류의 생식소 발달에 영향을 미치는 주요 인자로 알려져 있다(Ruiz et al. 1992). 이번 연구에서도 수온이 증가함에 따라 생식소가 발달하는 것 이 관찰 되었는데, 수온이 증가하는 시기에 평균 난경 크 기도 증가하고 수온이 낮아지는 시기에 난경 크기 또한 감소하였다. 이러한 현상은 키조개의 배우자형성 및 생식 소 발달에 있어 수온이 중요한 역할을 하고 있음을 짐작 할 수 있으며, 수온 증가는 난경 성장에 있어 필요한 조건 이라 판단된다. 이번 연구지역인 울릉도에 서식하는 암컷 과 수컷 키조개의 생식소 형성은 수온이 가장 낮은 기간 인 1월(12.6oC)부터 시작되는 것을 확인할 수 있었다.
Baik et al. (2001) 또한 서해안에 서식하는 키조개의 생식 소 역시 수온이 가장 낮은 시기인 11월부터 형성되기 시 작한다고 보고한 바 있다. 다른 이매패류 연구에서도 유사 한 결과들이 관찰되었다. Ke and Li (2013)는 중국 산동 반도의 북쪽 해안에 서식하는 Mactra veneriformis의 번식 주기를 관찰한 결과, 수온이 가장 낮은 시기인 12월 (3.8oC)에 생식소가 형성되기 시작한다고 발표하였다. 이 밖에 수온이 가장 낮은 시기에 생식소가 형성되는 현상 등은 중국, 포르투갈을 비롯한 국내에 서식하는 다양한 이 매패류들에서도 관찰되었다(Park et al. 2001; Joaquim et al. 2008). 그러므로 이와 같이 유사한 결과들로 미루어 볼 때, 겨울철 수온은 특정한 해양 이매패류들의 저장물질을 증가시키고, 이 저장물질에 의해 배우자 생성을 유발할 수 있는 것으로 판단된다. 울릉도 해역에 서식하는 키조개의 산란과 방정은 7월부터 11월까지 관찰되었는데, 주 산란 은 수온이 가장 높은 기간인 7−8월(23.6oC) 한차례 일어 났다. 이번 연구결과와 같이 수온이 가장 높은 시기에 산 란이 일어나는 현상은 호주에 서식하는 C. gigas와 중국 에 서식하는 M. veneriformis 등에서도 관찰이 되었다(Li et al. 2009; Ke and Li 2013).
이번 연구에서 수행된 키조개 생식소의 조직학적 관찰 결과, 울릉도 키조개의 산란기는 7월부터 9월까지 계속되 며, 주 산란기는 7−8월인 것으로 판단된다. Shim et al.
(2008)은 울릉도 주변해역의 식물플랑크톤 농도를 측정하 였는데, 이들 연구 결과에 따르면 식물플랑크톤 농도는 8월이 가장 높은 것으로 조사되었다. 이와 같은 현상은 울 릉도에 서식하는 키조개가 산란 시기에 좋은 먹이조건을 갖게 된다는 것을 의미한다. Kang et al. (2004)은 제주도 시흥리에 서식하는 바지락의 산란시기를 분석하였다. 시 흥리에 서식하는 바지락의 주 산란기는 7월과 8월로 식물 플랑크톤 번성시기와 밀접한 관계가 있다고 보고하였고, 이번 연구 결과와 유사하였다. 앞서 언급한 바와 같이 수 온과 해수 중의 가용먹이는 배우자 형성과정과 산란에 있 어 중요한 환경인자로 알려져 있다(Mann 1979b; Baron 1992). 이처럼 먹이조건이 충분한 시기에 산란하는 주된 이유는 생태학적으로 수정 후 발생한 플랑크톤 유생에게 알맞은 영양공급이 확실히 보장되어야 하기 때문이라 해 석된다(Shafee 1989; Hadfield and Anderson 1998).
난경 크기 분포를 관찰하는 것은 다양한 해양 이매패류
의 생식세포 형성 과정을 연구하는데 활용되어 왔으며, 생 식소 성숙에 있어 계절적 변화나 산란 빈도 측정 시 난경 의 월 변화 양상으로부터 추정된다(Southgate and Lee 1998; Park and Choi 2004; Chávez-Villaba et al. 2007;
Kang et al. 2009; Kim et al. 2010). 울릉도 키조개는 생 식소가 발달하면서 난경 또한 증가하였는데, 산란 직전인 5월과 6월은 40−60 μm 크기의 난들의 분포가 높게 관찰 되었다(Fig. 7). 산란이 시작된 7월부터 50 μm 이상의 난 들이 점점 감소하였으며, 8월에는 30−40 μm 사이의 난들 이 높은 비율을 차지하고 있었다. 반면, 9월에는 50 μm 이상의 난들이 75% 이상을 차지하고 있었는데, 이는 산란 후 잔존 난(RO)과 흡수되는 난들이 주로 관찰되었기 때문 이라 사료된다(Fig. 5F). 10월의 난경 분포 비율은 20 μm 이하의 난들이 90% 이상 차지하고 있는 것으로 보아 산 란이 종료된 것으로 판단되며, Fig. 6B에서 확인할 수 있 듯이 대부분의 개체들이 휴지기와 소모기 상태인 것으로 Fig. 7. Distribution of egg diameter of Atrina pectinata from November 2013 to October 2014
확인되었다. 이처럼 이번 연구에서 관찰된 조직발달변화 결과(Fig. 6)와 비교해 볼 때, 난경의 연중 변화 양상은 생식세포 변화 양상과 매우 밀접하게 연관되어 있고 생 식소 발달 과정을 잘 반영하고 있는 것을 확인할 수 있 었다.
5. 결 론
키조개와 관련된 대부분의 연구들은 남서해안에 서식하 는 개체군 위주로 진행되어 왔었다. 이번 연구는 울릉도 연안에서 발견된 키조개 군락 및 동해안 북부 해역에서 서식하는 키조개의 번식생태학적 연구와 관련된 첫 보고 이다. 조직학적 관찰, 생식소중량지수(GSI) 및 비만도(CI) 등의 결과들을 종합해 볼 때, 울릉도 남해역에 서식하는 키조개의 산란은 7월부터 9월까지 관찰이 되지만, 주 산 란시기는 7월부터 8월 사이 한차례인 것으로 사료된다.
그러나, 서해 보령에 서식하는 키조개의 산란은 6−7월 (Lee et al. 2006), 남해 득량만에 서식하는 키조개의 산란 은 같은 지역에 서식함에도 불구하고 6−7월, 8−9월 사이 로 해마다 다른 산란 패턴을 보이고 있었다(Lee et al.
2015). 이렇듯 같은 종임에도 불구하고 공간적·시간적으로 산란 시기가 다른 이유는 앞서 언급한 바와 같이 수온 및 먹이원 등 환경적인 차이일 것으로 판단된다. 이번 연구를 통해, 울릉도 남쪽 해안에 서식하는 키조개의 번식생리학 적 특성을 규명하였다. 이번 연구 결과는 울릉도 연안에 서식하는 키조개 군락지에 대한 자원 관리와 합리적이고 안정적인 생산을 위한 기초자료로 유용하게 사용될 것으 로 판단된다.
사 사
이 연구는 2013년 교육과학기술부 재원 한국연구재단 기초연구사업(No. 2013R1A1A2062015) 및 한국해양과학 기술원 ‘기후변화와 인위적 교란에 따른 울릉도 해양생태 계 반응(울릉도독도해양과학기지 지원)(PE99395)’ 과제의 지원에 의해 수행되었으며, 지원에 깊은 감사를 드립니 다. 또한, 시료채집에 도움을 주신 울릉군청 관계자 분들 에게 깊은 감사를 표합니다.
참고문헌
An HS, Lee JW, Dong CM (2012) Population genetic structure of Korean pen shell (Atrina pectinata) in Korea inferred from microsatellite marker analysis. Genes Genom 34:681−688
Baek MJ, Jee YJ, Choi KS, Lee WC, Park HJ, Kwak JH,
Kang CK (2014) Physiological disturbance of the Manila clam, Ruditapes philippinarum, by altered environmental conditions in a tidal flat on the west coast of Korea. Mar Pollut Bull 78(1−2):137−145
Baik SH, Kim KJ, Chung EY, Choo JJ, Park KH (2001) Seasonal variations in biochemical components of the visceral mass and adductor muscle in the pen shell, Atrina pectinata. Fish Aquat Sci 4(1):18−24
Baron J (1992) Reproductive cycles of the bivalve mollusks Atactodea striata (Gmelin), Grafrarium tumidum (Roding), and Anadara scapha (L.) in New Caledonia. Aut J Mar Freshwater Res 43(2):393−402
Chávez-Villaba J, Villelás-Avila R, Cáverse-Martinez C (2007) Reproduction condition and mortality of the Pacific oyster Crassostrea gigas (Thunberg) in Sonora, Mexico.
Aquac Res 38(3):268−278
Choe BL, Kim W, Lee JR, Yoon SH (1994) Pteriomorphia (Mollusca: Bivalvia) from Ullŭng Island, Korea. Korean J Syst Zool 10(1):61−83
Choi KS, Lewis DH, Powell EN, Ray SM (1993) Quanti- tative measurement of reproductive output in the American oyster, Crassotrea virginica, using enzyme- linked immunosorbent assay (ELISA). Aquac Res 24(3):
299−322
Chung EY, Baik SH, Ryu DK (2006) Reproductive biology of the pen shell, Atrina (Servatrina) pectinata on the Boryeong coastal waters of Korea. Korean J Malacol 22(2):143−150
Chung JS, Chung EY, Lee CH (2012) Gametogenic cycle and the number of spawning seasons by quantitative statistical analysis, and the size at 50% of group sexual Maturity in Atrina (Servatrina) pectinata (Bivalvia:
Pinnidae) in western Korea. Korean J Malacol 28(4):
363−375
Dohmen RM (1985) Gametogenesis. In: Verdonk NH, Van der Biggelaar JAM (eds) The mollusca (development).
Academic Press, London, pp 1−37
Drummond L, Mulcahy M, Cullory S (2006) The reproductive biology of the Manila clam, Ruditapes philippinarum, from the North-West of Ireland. Aquaculture 254:324−
340
Fabioux C, Huvet A, Souchu PL, Pennec ML, Pouvreau S (2005) Temperature and photoperiod drive Crassostrea gigas reproductive internal clock. Aquaculture 250(1−2):
458−470
Gaspar MB, Monteiro CC (1999) Gametogenesis and spawning in the subtidal white clam Spisula solida, in relation to temperature. J Mar Biol Assoc UK 79(4):
753−755
Modeling oyster populations. III. Critical feeding periods, growth and reproduction. J Shellfish Res 11(2):399−416 Hong SH, Lee JB, Im YJ, Hwang HJ, Yeon IJ, Park YC,
Ko TS, Ma CW, Park KY (2002) Population ecology of pen shell, Atrina pectinata, in the western waters of Korea-I. Geographic distribution and biomass. J Korean Soc Fish Res 5:12−23
Joaquim S, Matias D, Lopes B, Arnold WS, Gaspar MB (2008) The reproductive cycle of white clam Spisula solida (L.) (Mollusca: Bivalvia): implications for aquaculture and wild stock management. Aquaculture 281(1−4):43−
48
Kang, CK, Park MS, Lee PY, Choi WJ, Lee WC (2000) Seasonal variation in condition, reproductive activity and biochemical composition of the Pacific oyster, Crassostrea gigas (Thunberg) in suspended culture in two coastal bays of Korea. J Shellfish Res 19:771−778
Kang DH, Ahn IY, Choi KS (2003) Quantitative assessment of reproductive condition of the Antarctic clam, Laternula elliptica (King & Broderip), using image analysis. Invertebr Reprod Dev 44(1):71−78
Kang DH, Hyun CY, Limpanont Y, Choi KS (2007) Annual gametogenesis of the Chinese Anapella clam Coecella chinensis (Deshayes 1855) at an upper intertidal sandy beach on the east coast of Jeju, Korea. J Shellfish Res 26:433−441
Kang DH, Ahn IY, Choi KS (2009) The annual reproductive pattern of the Antarctic clam, Laternula elliptica from Marian Cove, King George Island. Polar Biol 32:517−
528
Kang DH, Ngo TTT, Koh KJ, Choi KS (2004) Seasonal changes in gonadal development of Manila clam, Ruditapes philippinarum from Shi-Heung-Ri, Jeju, Korea. J Aquacult 17(1):81−88
Kang SG, Choi KS, Bulgakov AA, Kim Y, Kim SY (2003)
Kim CW, Hur SB (1998) Distribution of the bivalve resources in Deukryang Bay. J Aquacult 11:249−260 Kim DH, Yoon HS, An YK, Lee SD, Choi SD (2008)
Density dependent growth and survival rates of Atrina pectinata in Duekryang bay, Korea. Korean J Malacol 24(2):137−142
Kim SK, Rosenthal H, Clemmesen C, Park KY, Kim DH, Choi YS, Seo HC (2005) Various methods to determine the gonadal development and spawning season of the purplish Washington clam, Saxidomus purpulatus (Sowerby).
J Appl Ichthyol 21:101−106
Lee YJ, Choi KS, Lee DS, Lee WC, Park HJ, Choy EJ, Kim HC, Kang CK (2015) The role of the adductor muscle as an energy storage organ in the pen shell Atrina japonica (Reeve, 1858). J Mollus Stud 81:502−
511
Lee SJ, Min KS, Kim BG, An CM, Ju SM, Lee JS (2006) Gonad development and gametogenic cycle of the pen shell, Atrina pectinata (Bivalvia: Pinnidae). J Kor Fish Soc 39(5):398−403
Li Y, Qin JG, Li X, Benkendorff K (2009) Monthly variation of condition index, energy reserves and antibacterial activity in Pacific oysters, Crassostrea gigas, in Stansbury (South Australia). Aquaculture 286:64−71
Limpanont Y, Yang HS, Won SH, Han SJ, Lee JB, Lee BG, Choi KS (2010) First report on the annual reproductive cycle of Burchardi’s cockle, Acrosterigma (=Vasticardium) burchardi Dunker 1877 (Bivalvia: Cardiidae) on a subtidal sand flat off southern Jeju Island, Korea. Invertebr Reprod Dev 54(1):27−34
Mackie GL (1984) Bivalves. In: Tompa AS, Verdonk NH, Van Den Biggelaar JAM (eds) The mollusca, vol 7.
Reproduction. Academic Press, London, pp 351−402 Mann R (1979a) Some biochemical and physiological
aspects of growth and gametogenesis in Crassostrea
gigas and Ostrea edulis grown at sustained elevated temperature. J Mar Biol Assoc UK 59:95−110
Mann R (1979b) The effect of temperature on growth, physiology, and gametogenesis in the Manila clam, Tapes philippinarum. J Exp Mar Biol Ecol 38:121−133 Mann R, Glomb SJ (1978) The effect of temperature on
growth and ammonia excretion of the Manila clam Tapes japonica. Estuar Coast Shelf Sci 6:335−339
Menzel W (1991) Estuarine and marine bivalve mollusk culture. CRC Press, Boca Raton, 362 p
Min DK (2004) Mollusks in Korea. Hanguel Graphics, Busan, 566 p
Moon TS, Jo PG, Kim BH, Park KY, Ku HD, Shin YK, Lym YS (2009) Induction of sex maturation and growth in comb pen shells, Atrina pectinata per microalgae classes. Korean J Malacol 25(2):105−112
Moura P, Gasper MB, Monteiro CC (2008) Gametogenic cycle of the smooth clam Callista chione on the southwestern coast of Portugal. J Mar Biol Assoc UK 88:161−167
NFRDI (1997) Studies on the development of techniques on pen shell natural spat collection. NFRDI, 148 p (in Korean)
NFRDI (2009) Research and development on artificial seed production of Comb Pen Shells, Atrina pectinata. NFRDI, TR-2010-AQ-008, 56 p (in Korean)
Okumuş I, Stirling HP (1998) Seasonal variations in the meat weight, condition index and biochemical composition of mussels (Mytilus edulis L.) in suspended culture in two Scottish sea lochs. Aquaculture 159:249−261 Okutani T (2000) Marine mollusks in Japan. Tokai University
Press, Tokyo, 1173 p
Olive PJW (1985) Physiological adaptations and the concepts of optimal reproductive strategy and physiological constraint in marine invertebrates. In: Laverack MS (ed) Physiological adaptations of marine animals. Symposia of the Society for Experimental Biology XXXIX. The Company of Biologists Ltd, Cambridge, pp 267−300 Park KI, Choi KS (2004) Application of enzyme-linked
immunosorbent assay for studying of reproduction in the Manila clam Ruditapes philippinarum (Mollusca: Bivalvia) I. quantifying eggs. Aquaculture 241:667−687
Park KI, Choi JW, Choi KS (2005) Development of a Saxidomus purpuratus (Mollusca: Bivalvia) egg-specific antibody for the quantification of eggs using an enzyme- linked immunosorbent assay. J Shellfish Res 24(4):1079− 1085
Park MS, Kang CK, Lee PY (2001) Reproductive cycle and biochemical composition for the ark shell Scapharca
broughtonii (Schrenck) in a southern coastal bay of Korea. J Shellfish Res 20(1):177−184
Ruiz C, Abad M, Sedano F, Garcia-Martin LO, Sanchez Lopez JL (1992) Influence of seasonal environmental changes on the gamete production and biochemical composition of Crassotrea gigas (Thunberg) in suspended culture in El Grove, Galicai, Spain. J Exp Mar Bio Ecol 155(2):249−262
Ryu DK, Baik SH, Park KH, Chung EY (2001) Age and growth of the Pen Shell, Atrina (Servatrina) pectinata japonica (Reeve), on the west coast of Korea. Korean J Malacol 17(2):71−78
Saxby S (2002) A review of food availability, sea water characteristics and bivalve growth performance at coastal culture sites in temperate and warm temperate regions of the world. Fisheries Research Division WA Marine Research Laboratories, Western Australia, Fisheries Research Report, vol. 132, 42 p
Schultz PWW, Huber M (2013) Revision of the worldwide Recent Pinnidae and some remarks on fossil European Pinnidae. Conchbooks, Bahnhofstr, 164 p
Shafee MS (1989) Reproduction of Perna picta (Mollusca:
Bivalvia) from the Atlantic coast of Morocco. Mar Ecol- Prog Ser 53:235−245
Shim JM, Yun SH, Hwang JD, Jin HG, Lee YH, Kim YS, Yun SC (2008) Seasonal variability of picoplankton round Ulleung Island. J Environ Sci 17(11):1243−1253 Son PW, Ha DS, Lee CH, Jang DS, Kim DK (2005) Study
on the natural spat collection of the pen shell, Atrina pectinata. Korean J Malacol 21(2):113−120
Southgate PC, Lee PS (1998) Hatchery rearing of the tropical blacklip oyster Saccostrea echinata (Quoy and Gaimard). Aquaculture 169(3−4):275−281
Statistics Korea (2015) Korean statistical information service.
http://kosis.kr/eng Accessed 1 Dec 2015
Uddin MJ, Yang HS, Choi KS, Kim HJ, Hong JS, Cho MJ (2010) Seasonal changes in Perkinsus olseni infection and gametogenesis in Manila clam, Ruditapes philippinarum, from Seonjaedo Island in Incheon, off the west coast of Korea. J World Aquac Soc 41(S1):93−101
Uddin MJ, Yang HS, Park KJ, Kang CK, Kang HS, Choi KS (2012) Annual reproductive cycle and reproductive efforts of the Manila clam Ruditapes philippinarum in Incheon Bay off the west coast of Korea using a hitology- ELISA combined assay. Aquaculture 364−365:25−32 Ulleung-Gun (2013) Ecological survey of marine fauna and
flora in coastal Ulleungdo Island. Ulleung-Gun, BSPG48060- 10360-3, 132 p (in Korean)
Walne PR (1976) Experiments on the culture in the sea of