Characterization of the Lsi1 Homologs in Cucurbita moschata and C. ficifolia for Breeding of Stock
Cultivars Used for Bloomless Cucumber Production
Jaemin Jung1†, Joonyup Kim3†, Bingkui Jin1, Youngmi Choi1, Chang Oh Hong2, Hyun Ho Lee2, Youngwhan Choi1, Jumsoon Kang1, and Younghoon Park1,3*
1Department of Horticultural Bioscience, Pusan National University, Miryang 50463, Korea
2Department of Life Science and Environmental Biochemistry, Pusan National University, Miryang 50463, Korea
3Life and Industry Convergence Research Institute, Pusan National University, Miryang 50463, Korea
*Corresponding author: [email protected]
Bloomless cucumber fruits are commercially produced by grafting onto the pumpkin stocks (Cucurbita moschata) to restricted silicon (SiO2) absorption. Inhibition of silicon absorption in bloomless stocks is conferred by a mutant allele of the CmLsi1 homologous to Lsi1 in rice.
In this study, we characterized the Lsi1 homologs in pumpkin (C. moschata) and its cold- tolerant wild relative C. ficifolia (‘Heukjong’) in order to develop a DNA marker for selecting a bloomless trait and to establish the molecular basis for breeding bloomless stock cultivars of C. ficifolia. A Cleaved amplified polymorphic sequence (CAPS) marker (CM1-CAPS) was designed based on a non-sysnonymous single nucleotide polymorphism (SNP, C>T) of the CmLsi1 mutant-type allele, and its applicability for Marker-assisted selection (MAS) was confirmed by evaluating three bloom and five bloomless pumpkin stock cultivars. Quantitative RT-PCR of the CmLsi1 for these stock cultivers implied that expression level of the CmLsi1 gene does not appear to be associated with the bloom/bloomless trait and may differ depending on plant species and tissues. A full length cDNA of the Lsi1 homolog [named CfLsi1(B+)]
of ‘Heukjong’ (C. ficifolia), was cloned and sequence comparison between CmLsi1(B+) and CfLsi1(B+) revealed that there exists total 24 SNPs, of which three were non-synonymous.
Phylogenetic analysis of CfLsi1(B+) and Lsi1 homologs further revealed that CfLsi1(B+) is closesly related to Nodulin 26-like intrinsic proteins (NIPs) and most similar to CpNIP1 of C.
pepo than C. moschata.
This work was supported by a grant (710001- 07-5) from the Vegetable Breeding Research Center through Agriculture, Food and Rural Affairs Research Center Support Program, Ministry of Agriculture, Food and Rural Affairs (MAFRA), Korea. Further, this research was supported by the National Agricultural Genome Program (Project No. PJ010438022015), Rural Development Administration.
HORTICULTURAL SCIENCE and TECHNOLOGY 35(3):333-343, 2017
URL: http://www.kjhst.org pISSN : 1226-8763 eISSN : 2465-8588
This is an Open-Access article distributed under the terms of the Creative Commons Attribution NonCommercial License which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
Copyrightⓒ2017 Korean Society for Horticultural Science.
OPEN ACCESS Received:
Accepted:
March 2, 2017 April 24, 2017
Abstract
Additional key words: cucurbitaceae, Lsi2, Low silicon rice 1, marker-assisted breeding, transporters, RACE PCR
Introduction
Cucumbers (Cucumis sativas L.), theCucurbitaceae, arefruitsandvegetableswithatotalannual productionof75milliontons (FAO, 2014). TotalproductionofdomesticcucumberinKoreais271,040
†The authors contributed equally to this manuscript.
tonsperyear (KOSIS, 2015), ofwhich28.4tonsisexportedtoJapan (UNI-PASS, 2015). FormarketabilityinJapan, cucumber fruitwithoutbloom (bloomlesscucumber) isinevitablebecauseitdisplaysacleanandshinyappearanceonthesurfaceofthe fruit (Choietal., 2013; Mitanietal., 2011). InmajorAsiancountriesincludingJapanandKorea, orientalpumpkin (C. moschata Duch.) isusedasastockingraftingofcucumberforpreventionofFursariumwiltdisease, maintenanceofvegetativegrowth, andimprovementofcoldtolerance. ForproducingcucumbersexportedtoJapan, therefore, aspecifictypeofpumpkinstock (bloomlessstock) thatpreventsthecucumberfruitfrombloomingmustbeused (Seoetal., 2004; Choietal., 2013). Theseedsfor bloomlessstockareimportedmainlyfromJapan, whilenodomesticstockcultivarsarecommerciallyavailabletodate.
Forproductionofwintergreenhousecucumber, awildtypepumpkincultivar ‘Heukjong’, (C. ficifolia Bouche.) iswidely usedasastockduetoitsnatureofcold-toleranceoverothernon-tolerantpumpkinstocks (Tachibana, 1988; Seoetal., 2004).
However, ‘Heukjong’ producesbloomingraftingofthecucumberandisinadequateforcultivatingbloomlesscucumbersinthe winterseason. Furthermore, despitethenecessityforcold-tolerantbloomlessstockcultivars, introgressionofthebloomlesstrait into C. ficifolia from C. moschata isnotstraightforwardduetotheinfeasibilityofcross-fertilizationbetweenthesetwospecies (RobinsonandDecker-Walters, 1997).
Thebloomofcucumberfruitisgeneratedwhenabsorbedsilicon (SiO2) isexudedwithwaterontotheskinandthendried (Yamamotoetal., 1989). Agenethatfacilitatestheuptakeofsilicon, Low silicon rice 1 (Lsi1), wasfirstidentifiedinrice (Maet al., 2006), anditshomologousgeneswerealsoidentifiedinmanyotherplantsincludingmaizeandbarley (Chibaetal., 2009; Mitanietal., 2009). Therearetwotypesoftransportersthatmediatesiliconabsorption, achannel-typetransporter (oraninflux transporter) andaneffluxtransporter (MaandYamaji, 2015). Inrice, the Lsi1 geneencodesachannel-typetransporterthat facilitatespassivetransportofsiliconacrosstheplasmamembranebetweenapoplastandtheplantcell (Yamajietal., 2008), whileaneffluxtransportismediatedbythe Lsi2 genethatbelongstoanuncharacterizedaniontransporterfamily (Maand Yamaji, 2015). Inresponsetosilicon, expressionof Lsi1 wasdown-regulatedinriceandcucumber (Maetal., 2006; Sunetal 2017), butwasunalteredforitshomologousgenesinmaizeandbarley (Chibaetal., 2009; Mitanietal., 2009). Recently, itwas reportedthatpumpkinhomologs, CmLsi1 and CmLsi2, areassociatedwiththetransportofsilicon (Mitanietal., 2011; Mitani- Uneoetal., 2011). Furthermore, abloomlesspumpkincultivar ‘Super-unryu’ possessesamissensemutation (P>L) atposition 242aminoacidthatislinkedtothetransportactivityofsilicon (Mitanietal., 2011).
DNAmarkershavebeenemployedtoidentifyDNApolymorphismsandanalyzegenotypesandgeneticlinkagebetween genesandtraits (LeeandChung, 2011; KwonandChoi, 2013). Inmodernbreedingstrategies, Marker-assistedselection (MAS) isthemostpopularapplicationoftheDNAmarker. MASshortensthetimerequiredforbreeding, isnotinfluencedby environmentalfactors, andcangreatlyincreasereliabilityandprecisionincomparisontophenotype-basedselections (Tanksley, 1983; CollardandMackill, 2008; Jonahetal. 2011). Althoughithasbeenreportedformutationaleffectof CmLsi1 relatedtothe bloomlesstrait (Mitanietal., 2011), aDNAmarkerthatcanbeusedtoanalyzeallelicormutationalvariationsofthe CmLsi1 geneisnotpubliclyavailable.
Here, wedevelopedamolecularmarkerforselectionofabloomlesstraitbasedonnucleotidepolymorphismsof CmLsi1. We clonedthehomologousgeneof Lsi1 in C. ficifolia toestablishthemolecularbasisforbreedingofcold-tolerantbloomlessstock cultivarsof C. ficifolia..
Materials and Methods
Plant Materials
Threecultivarsofbloompumpkinstocks [‘Heukjong’ (KyungNongSeed, Busan, Korea), ‘Odaetojwa’ (KyungNongSeed, Busan, Korea), and ‘Arirang’ (Koregon, Anseong, Korea)] andfivecultivarsofbloomlesspumpkinstocks [(‘OhmaiStock’ (KyungNongSeed, Busan, Korea), ‘OhmaiSummerStock’ (KyungNongSeed, Busan, Korea), ‘Union’ (Koregon, Anseong, Korea), ‘Nunbusyeo’ (Koregon, Anseong, Korea), ‘Newtype’ (Koregon, Anseong, Korea)] wereusedforthedeterminationof siliconuptakeandgeneanalysis.
Gene-Based DNA Marker
Extraction of DNA: GenomicDNAwasextractedfromyoungleavesofthetwo-trueleafstageoftheplant. Collectedleaf tissueswereplacedintoa1.5mLmicro-centrifugetubewithbeadsand600μLofDNAextractionbufferandsubjectedto Tissuelyser (TissueLyserII, QIAGEN, Venlo, Netherlands) forhomogenization. Sampleswerelysedina65°Cwaterbathfor45 minandfurtherincubatedbyadding200μLof7.5Mammoniumacetateonicefor15 - 20min. Incubatedsampleswere separatedbycentrifugationfor10minat14,240xg. Thesupernatantofcentrifugedsampleswasplacedina1.5mLtube containing5μL (5mg ∙ mL-1) ofglycogensolutionand600μLofisopropanolina1.5mLtubeandcentrifugedfor10minat 14,240xg. Afterdecantingthesupernatant, remainingpelletwaswashedusing300μLof 70% EtOH. Thewashedpelletwas resuspendedin200μLof 0.1MTris. ConcentrationofDNAwasmeasuredusingaNanodrop1000spectrophotometer (Thermo Scientific, Waltham, MA, USA) and20ng ∙ μL-1wasusedforPCRreactions.
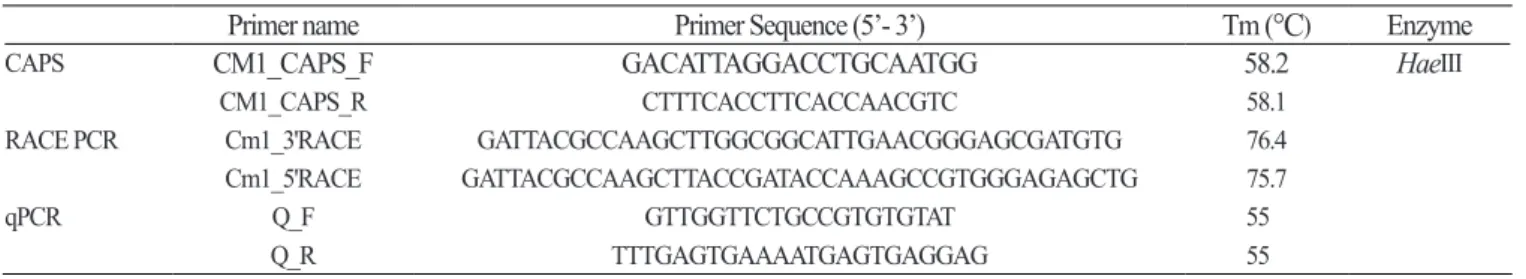
Development of CAPS Marker: CodingDNASequences (CDS) ofwild-typeallele CmLsi1(B+) (AB551949) andmutant- typeallele CmLsi1 (B-) (AB551950) ofthe C. moschata Lsi1 wereobtainedfromNCBIandalignedforcomparisonusing CLUSTALW (www.genome.jp/tools/clustalw) andconfirmedfortheSNPofthemutantallele (Fig. 1). IdentifiedSNPswere searchedforrestrictionsites (NEBcutterV2.0, NEB®, Ipswich, MA, USA) togenerateaCleavedAmplifiedPolymorphic Sequence (CAPS) markerusingPrimerpremier5 (PREMIERBiosoft, PaloAlto, CA, USA) (Fig. 1). PCRconditionsfor genotypingconsistedof1μLoftemplateDNA (20ng ∙ μL-1), 0.5μLofforwardandreverseprimer (10pmol), 0.1μLofTaq polymerase (5U ∙ μL-1, eTaqSolg ™, SolGent, Daejeon, Korea), 0.2μLofdNTPs (Solg ™, SolGent, Daejeon, Korea), 1μLof 10Xbuffer (Solg ™, SolGent, Daejeon, Korea) and6.7μLofddH2Oinatotalvolumeof10μL. Reactionswereincubatedat 95°Cfor2min, andcycled35timesasfollows: denaturationat94°Cfor15sec, annealingat58°Cfor30sec, extensionat72°C for1minusingaPCRcycler (T100 ™, BIO-RAD, Hercules, CA, USA). Afterthelastcycle, reactionswereincubatedat72°C for3min. RestrictiondigestionofPCRproductswasperformedbyadding0.3μLof Hae III (10,000U ∙ mL-1, Time-Saver ™, NEB®, Ipswich, USA), 1.5μLof10Xbuffer (CutSmart ™, NEB®, Ipswich, Massachusetts, USA), 3.2μLofddH2OtothePCR productandincubatedat37°Cfor1h. Digestedproductswererunona2% agarosegelat70Vfor40minandstainedwithEtBr, andcheckedfortheresultsusingGelImageAnalysisSystem (CoreBioi-MAXTM, Davinch-K, Seoul, Korea).
Analysis of CmLsi1 Gene Expression
RNA Extraction and cDNA Synthesis: Youngleavesandrootsfromatleastthreeplantspercultivarwerecollectedand
immediatelyfrozeninliquidnitrogen. TotalRNAwasextractedusingaSeed/FruitKit (Ribospin™, GeneAll®, Seoul, Korea) accordingtothemanufacturer’sinstructions. QuantitatedRNAusingNanodrop1000 (ThermoScientific, Waltham, MA, USA) wasdilutedto10ng ∙ μL-1andusedforcDNAsynthesiswithanRTPremixturekit (HyperScript ™, GeneAll®, Seoul, Korea) followingthemanufacturer’sinstructions.
Quantitative PCR (qPCR): Gene-specificprimers (Q- FandR) for CmLsi1 (B+) (AB551949) weredesignedusingPrimer premier5 (Table1). Quantitativereal-timeRT-PCR (qPCR) wasperformedinaReal-timePCRcycler (LightCycler®480, Roche, Basel, Switzerland) for ‘Heukjong’, ‘Odaetojwa’, ‘OhmaiStock’, and ‘OhmaiSummerStock’. Reactionswereperformed byadding1μLofcDNA (10ng ∙ μL-1), 0.5μLofeachforwardandreverseprimer (10pmol), 5μLof2XSYBRGreenIMaster (Roche, Basel, Switzerland), and3μLofddH2Oinatotalvolumeof10μL. CmACTIN wasusedasareferencegeneasdescribed byObreroetal. (2011) (Table1). Allreactionswereincubatedat95°Cfor5min, andcycledfor45timesasfollows: 95°C for10 sec, 55°Cfor20sec, 72°Cfor1min. Relativequantitationofthe Lsi1 transcriptto CmACTIN genewasperformedusingthe 2-ΔΔCTmethod (LivakandSchmittgen, 2011).
Cloning of Lsi1 Homolog in C. ficifolia
5’ and 3’ RACE: Thefull-lengthcDNAsequenceof CmLsi1 homologousgenefrom C. ficifolia wasobtainedusingRACE 5'/3' Kit (SMARTer®, Takara, Kusatsu, Japan). TotalRNAwasextractedasoutlinedaboveandRACEreactionswereperformed accordingtothemanufacturer’sinstructions. Gene-specificprimersweredesignedfrom CmLsi1 (B+) CDS (AB551949) with primerlengthsof23 - 28bp, GCcontentsof50 - 70%, Tm>70°Cand15bp5'- endprimeroverlapsequence (GATTACGCCAAGCTT).
Cloning and Sequencing of CfLsi1: RACEreactionswerecheckedonanagarosegelandpurifiedtheproductusingagel extractionkit (ExpinTM, GeneAll®, Seoul, Korea). PurifiedRACEproductwasclonedintoT-EasyVectorSystemI (pGEM, Progma, Madison, WI, USA) andverifiedthesequencebythedyeterminatormethod (Genotech, Daejeon, Korea).
Sequence Alignment and Phylogenetic Analysis
Thenucleotidesequenceof CfLsi1 wasusedasaquerysequencetoconductBLASTsearchestoidentifyhomologsofthe CfLsi1 fromNCBI (www.ncbi.nlm.nih.gov). Todeterminetherelationshipbetweensearchedsequences, aphylogenetictree (dendrogram) ofhomologswasgeneratedwithMEGA5 (Tamuraetal., 2011) usingtheNeighbor-Joining (NJ) algorithmand Tamura-Neiparametermodelwith1,000bootstrapreplicates.
Table 1. PCR primers used in this study.
Primer name Primer Sequence (5’- 3’) Tm (°C) Enzyme
CAPS CM1_CAPS_F GACATTAGGACCTGCAATGG 58.2 HaeIII
CM1_CAPS_R CTTTCACCTTCACCAACGTC 58.1
RACE PCR Cm1_3'RACE GATTACGCCAAGCTTGGCGGCATTGAACGGGAGCGATGTG 76.4
Cm1_5'RACE GATTACGCCAAGCTTACCGATACCAAAGCCGTGGGAGAGCTG 75.7
qPCR Q_F GTTGGTTCTGCCGTGTGTAT 55
Q_R TTTGAGTGAAAATGAGTGAGGAG 55
Measurement of Silicon Uptake
Seedsofeightcommercialcultivars (seeabove) weresowninBarokerpottingsoil (SeoulBio, Eumseong, Korea) ina50 - holedtray. After25daysofsowing, seedlingsweretransplantedinaplasticpot (15cm × 13cm × 19cm) andgrownfor30days inagreenhouseatPusanNationalUniversity (Miryang, Korea). Whiletransplanting, thesoilcollectedfromapaddyfieldofthe AgricultureResearchStationatPusanNationalUniversitywasair- driedandsifted, and2.4kgof thesoilwasfilledintheplastic pot. Threeplantsofeightcultivarsweregrowninacompletelyrandomizeddesignwiththreebiologicalreplicates. Bulkdensity ofsoilwas1.37g ∙ cm-1, and576mLofdistilledwaterwasaddedsothatthewatercontentinsoilwas60% oftheporevolume withaparticledensityof2.65g ∙ cm-1. Aftertransplanting, thesilicatefertilizer (NonepongEco, Nousbo, Suwon, Korea) was dissolvedin100mLoftapwaterandaddedattherateof0 (untreatedcontrol), 100and200mgSi ∙ kg-1ineachpot. Throughout thewholegrowth, nitrogen (12.8mg ∙ kg-1), phosphoricacid (8.75mg ∙ kg-1) andpotassium (12.7mg ∙ kg-1) andcompost (1g ∙ kg-1) weresupplementedwiththesameamount, andwaterwasaddedtocompensatethedecreasedamountbyweighingthepotonce everyfourdays. After30daysoftransplanting, wholeplantswerecollectedfromsoilanddriedinadryovenat70°Cfor72hand thenpulverized. Onegramofthepulverizedsamplewaslysedinthelysisbuffer (H2SO4:HClO:H2O = 5: 9:1, Samchun, Pohang, Korea) andthenthecontentofsiliconwasanalyzedwithICP-AES (GBCmodelX-100, GBC, Melbourne, Australia) .
Results and Discussion
Development of CmLsi1 Marker
CodingDNASequences (CDS) ofwild-typeallele (B+) andmutant- typeallele (B-) ofthe CmLsi1 wereusedforalignmentto confirmthenon-synonymousSNP (C>T) missensemutation (P>L). TodesignaCAPSmarkerfortheidentifiedSNPs, restriction enzymesiteswithinthesequenceweresearchedandrecognitionsitesof Hae IIIwasselectedforthestudy. APCRprimerset CM1-CAPS(Table1) wasdesignedspanningthe Hae IIIrestrictionsitetocuttheamplifiedproduct (201bp) into140bpand61 bpinthemutant-typeallele(Fig. 1). GenotypinganalysisoftheeightstockcultivarsusingtheCM1-CAPSmarkerrevealedthat bloomandbloomlesstraitswerecorrelatedwitheachgenotype (Fig. 2). ForabloomF1cultivar ‘Arirang’, markeranalysis indicatedthatitisheterozygousgenotypefor CmLsi1 (YamajiandMa, 2007), whichsuggeststhatoneoftheparentallinesof
‘Arirang’ wasabloomlesscultivarhomozygousforthemutantalleleof CmLsi1. Fromthis, weconfirmeduseoftheCM1- CAPSmarkerforMAS-assistedbreedingofbloomlesstrait.
Fig. 1.Coding DNA sequence (867 bp) of the CmLsi1(B+) (AB551949). PCR primers for CM1_CAPS marker (yellow) and quantitative real time RT-PCR (red) are color-coded. Restriction site used in this study for HaeIII is boxed and the SNP (C/T) to CmLsi1(B-) (AB551950) is red-colored.
769 846 ATAAAGGACTTTGGGTGTACTTTGTTGGGCC/TGGTTACAGGAACCCTATTAGGGGCATGGTCATATAAGTTCATAC
CAACCGATACCAAAGCCGTGGGAGAGCTGGCAGGTTTGGCAGTTGGTTCTGCCGTGTGTATCACATC 617 694 CATCTTGGCTGGACCCGTATCAGGTGGGTCGATGAACCCTGTGAGGACATTAGGACCTGCAATGGCAAGTGATAATT
GTGCCAGTGATAAACCTGTGCACTTAATTTCTCCTCACTCATTTTCACTCAAACTTCGAAGAATGTCAAGATCTGAC GTTGGTGAAGGTGAAAGATGA 857
Transcripts of CmLsi1 in Bloom and Bloomless Stock Cultivars
Expressionlevelsofendogenous CmLsi1 genewereexaminedinleafand roottissuesfrom bloom (‘Heukjong’ and
‘Odaetojwa’) andbloomless (‘OhmaiStock’ and ‘OhmaiSummerStock’) cultivarsusingQuantitativerealtimeRT- PCR (qRT- PCR) (Fig. 3). Fromthetissuestestedforrepresentativecultivars, relativeexpressionof CmLsi1 washigherinrootthanleafof thesecultivars (Fig. 3). Theexpressionpatternof Lsi1 wasconsistentwiththeresultfromrice (Oryza sativa) andcucumber (Cucumis sativus), whichshowedhigherexpressionof Lsi1 inrootthanleaf (Maetal., 2006; Sunetal., 2017). Amongcultivars ofpumpkins (C. moschata), ithasbeenshownthatexpressionof Lsi1 geneinabloomlesscultivar ‘Super-unryu’ washigherthan abloomcultivar ‘Sintosa’ (Mitanietal., 2011). Curiously, itwasalsoreportedthatexpressionof CmLsi1 inshootapexwas Fig. 2. Agarose gel (2%) image showing the result of CM1_CAPS genotyping for the CmLsi1 gene in eight stock cultivars.
Lane 1, ‘Heukjong’; 2, ‘Odaetojwa’; 3, ‘Arirang’; 4, ‘Nunbusyeo’; 5, ‘Ohmai Stock’; 6, ‘Ohmai Summer Stock’; 7, ‘Union’; and 8,
‘Newtype’.
Bloom Bloomless
2
1 3 4 5 6 7 8 M
200 bp
100 bp
Leat
Heukjong Odaetojwa Ohmai Ohmai Heukjong Odaetojwa Ohmai Ohmai
Stock Stock
Stock Stock
Summer Summer
Root
Relative to Heukjong Leaf
0 5 10 15 20 25 30