멜론 체세포배의 조직학적 특징
최필선*, 권석윤1
남부대학교 한방제약개발학과 약용식물형질전환연구소, 1한국생명공학연구원 그린바이오연구센터
Histological Characteristics of Somatic Embryos in Melon (Cucumis melo L.)
Pil Son Choi* and Suk Yoon Kwon1
Medicinal Plant Transformation Center, Department of Oriental Pharmaceutical Development, Nambu University, Gwangju 506-824, Korea
1Green Bio Research Center, Korea Research Institute of Bioscience and Biotechnology, Daejeon 305-806, Korea
Abstract - Hypocotyls explants of melon seedling were cultured on Murashige and Skoog’s (MS) medium supplemented with 1 mg/L 2,4-dichlorophenoxy acetic acid (2,4-D) and 0.5 mg/L benzyl aminopurine (BA) for 6 weeks to produce somatic embryos. In somatic embryos produced through intervening bright yellow friable (BYF) from the explants, somatic embryos with two-cotyledon (26%) and horn-type cotyledon (74%) were observed. The procambial strand of cotyledons was originated from circular procambial tissues of lower hypocotyls. The circular procambial independently divided into two procambial strand at the edge of cotyledonary-node, and then connected to each cotyledon to form somatic embryos with two- cotyledon. When cotyledon was horn-type, the circular procambial strand in lower hypocotyls would continuously remain connected to the cotyledon. However, somatic embryos with two or horn type cotyledon formed an abnormal shoot apex without the tunica-corpus structure or dome shape in the inter-cotyledonary area. These results demonstrated that the variation of cotyledon in somatic embryos was closely related to procambial tissue differentiation and shoot apical formation.
Key words -Apical meristem, Fused cotyledon, Histological characteristics, Melon, Somatic embryo, Two- cotyledon
*교신저자(E-mail) :[email protected]
서 언
멜론은 박과 작물에 속하는 일년생 초본식물로서 세계적 으로 중요한 경제 작물 중 하나이다. 식물조직배양을 통해 기관발생이나 체세포배 발생이 가능하고, 다른 식물 종과 마찬가지로 조직절편으로부터 직접 또는 캘러스 형성을 거 쳐 식물체를 생산할 수 있다(Tabei et al., 1991; Gray et al., 1993; Choi et al., 1994). 멜론 조직절편 배양과정에 서 호르몬 조합이나 배양절편에 따라 체세포배가 형성될 수 있으며, 이러한 체세포배의 형태와 기능은 자연상태의 접합자배와 유사한 것으로 여겨져 왔다(Ammirato, 1987).
그러나 최근 일부 연구에서 체세포배와 접합자배는 형태와 기능적으로 많은 차이가 있고(Choi et al., 2005), 그 원인
은 탄수화물의 종류와 농도(Soh et al., 2001), 2,4-D, ABA 및 BAP와 같은 식물 호르몬(Choi et al., 1994; Lee and Soh, 1993; Liu et al., 1993) 등이 중요한 원인으로 것으로 알려져 있다.
체세포배는 구형기를 거쳐 심장형기로 발달할 때 구형배 선단부에 있는 하표피(sub-epidermis) 시원세포가 분화 되어 자엽이 발달되고(Ammirato, 1987), 조직학적으로 심장형기 체세포배의 하배축으로 부터 전형층 조직이 분화 된 후 자엽 절과 자엽으로 이어진다는 연구결과에 따라 자 엽형성에 전형층조직의 중요성이 강조된 바 있다(Choi et al., 2005). 특히 자엽의 비정상적인 형태 형성은 전형성층 수의 분화와 밀접한 상관관계가 있어 하배축 부위에서 자 엽 절 부위에 이를 때 자엽의 수와 동일한 수의 전형층 조 직이 형성되는 것으로 알려져 있다(Choi et al., 2005). 한
Original Research Article
편 비정상형태의 자엽을 갖는 체세포배는 유경 조직이 흔 적 또는 미 분화된 상태로 남아 있거나 혹은 2개의 유경 조 직이 발달되어(Choi et al., 2005) 식물체로의 전환에 중 요한 영향을 미치는 것으로 알려져 있다(Soh et al., 2001;
Lee, 1993). 특히 이러한 비정상 형태의 체세포배는 식물 체 재생율을 감소시키거나 높이는 경향을(Gray and Mortensen, 1987; Soh et al., 2001)을 보이기 때문에 체 세포배를 이용한 식물체 대량생산이나 유전자 도입을 통한 형질전환연구를 수행하기 위해서는 체세포배의 발생학적 이해가 선행 되어야 할 것이며, 이는 체세포배에서 전형성 층 조직의 분화와 유경조직의 발달에 대한 조직학적 이해 가 필수적이라 할 수 있다.
따라서 본 연구에서는 멜론의 식물체 대량생산을 위한 기초 학술적 연구로서 기내배양과정에서 형성된 체세포배 를 형태에 따라 분류하고, 이들의 자엽 형성 초기와 유경조 직의 발생에 대한 조직학적 변화를 조사하였다.
재료 및 방법
체세포배 유도
멜론(Cucumis melo L.)종자를 70% 에탄올에 1분, 1%
sodium hypochlorite용액에 15분간 처리하여 표면 살균 한 후 멸균수로 3-5회 수세하였다. 이들 종자는 멸균 여과 지에서 물기를 제거한 후 MS(Murashige and Skoog, 1962)기본배지가 25 ml씩 분주된 페트리디쉬에 9개씩 치 상 하여 7-8일 동안 암조건으로 무균 발아 하였다. 길이가 5 cm정도 된 유식물체로부터 배축을 5 mm크기로 잘라 배 양재료로 사용하였다. 배발생 캘러스 및 체세포배를 유도 하기 위하여 1 mg/L 2,4-D와 0.5 mg/L BA가 조합 첨가 된 MS배지에 배축 절편을 치상하여 6주 정도 암 배양 하였 다. 배양 6주 정도 경과 하였을 때 배축 절편으로부터 형성 된 캘러스중 노란색의 단단한 배발생 캘러스를 해부 현미 경하에서 선발하였으며, 동일배지에서 4주 간격으로 계대 배양하면서 증식하였다. 배발생캘러스로부터 체세포배를 유도하기 위하여 호르몬이 첨가되지 않은 MS기본배지에 옮겨 3주간 명조건(cool-white형광등, 약 1,000 lux, 16 시간 광주기)에서 배양하였다. 배양 3주 후 형성된 체세포 배 형태를 해부 현미경 하에서 관찰 하였으며, 특히 자엽의 수와 모양에 따라 분류하여 기록하였다.
조직학적 특징
멜론 체세포배에서 자엽의 발생과 전형성층 조직의 분 화, 유경 조직의 발달을 조직학적으로 관찰하기 위하여 기 내 배양과정에서 얻은 2개 자엽을 갖는 정상 체세포배와 합생 자엽을 갖는 비정상 체세포배를 해부현미경하에서 조 심스럽게 선별하였다. FAA에 24시간 이상 고정 시킨 후 부탄올 시리즈로 탈수하여 파라플라스트에 포매 하였다.
회전식 마이크로톰으로 10 um 두께의 절편을 만들어 헤마 톡실린으로 염색한 후 광학 현미경하에서 각 절편을 연속 관찰 하였다(Choi et al., 2005).
결과 및 고찰
배양 1주째에 배축 절편으로부터 캘러스 형성은 거의 없 으나, 약간의 신장현상을 보였다. 배양 2주째부터 배축 절 단면으로부터 연한 노란색과 진한 노란색의 캘러스가 느리 게 생장되기 시작하였고, 점차 배양기간이 경과 될수록 이 들 캘러스는 연한 노란색의 부드러운 캘러스가 증식되기 시작하였다. 배양 4주 후 연한 노란색의 캘러스로부터 표 면에 구형기의 체세포배가 형성되어 배발생 캘러스로 간주 하였다. 이러한 캘러스를 해부현미경하에서 조심스럽게 선 별한 후 동일배지상에서 4주 간격으로 배양한 결과 2달 경 과 후부터 부서지기 쉬운 연한 노란색의 캘러스가 왕성하 게 증식되었고, 일부에서 구형 또는 심장형기의 체세포배 가 형성되기도 하였다. 이러한 캘러스는 1년 이상 체세포배 생산능력을 유지시킨 상태로 안정하게 계대배양 할 수 있 었으며, 식물호르몬이 첨가되지 않은 배지에 옮겨 약 2주 정도 배양하였을 때 많은 자엽기의 체세포배를 얻을 수 있 었다. 체세포배중에는 정상적으로 2개의 자엽을 갖는 것 (26%)과 자엽이 융합된 나팔형 (74%)의 체세포배도 관찰 되어 전자보다 후자가 약 3배정도 높은 빈도로 나타났다 (데이터 미제시). Jelaska(1974)에 의해 박과작물의 체세 포배 발생연구가 시작된 이래 호박, 오이 등과 같이 멜론에 서 배발생캘러스 유도는 오옥신 단독처리보다는 약간의 사 이토키닌의 조합 첨가가 효과적인 것으로 알려져 있다 (Kageyama et al., 1990). 본 연구에서도 배발생캘러스 유도는 2,4-D와 BA혼합 처리하였을 때 얻어졌으며, 이는 Choi(1994) 등의 보고와도 일치하였다. 또한 멜론 배양과 정에서 얻어지는 연한 노란색의 캘러스는 호박류(Jelaska, 1974)와 오이(Orezyk and Malepszy, 1985)에서도 언급
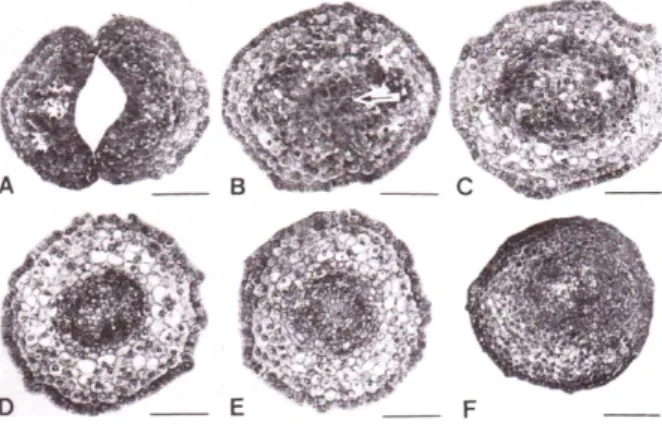
Fig. 1. Transverse sections of early heart‐shaped embryo with two cotyledons in tissue cultures of Cucumis melo L. were made from radicle to cotyledons. Procambial tissues in the radical (D-F), lower hypocotyl (C) and cotyledonary-node (B, arrow for plumule) are transformed into two procambial tissues, which are run into two cotyle- dons (A). All bars = 70 µm.
Fig, 2. Transverse sections of somatic embryo with fused cotyledons in tissue cultures of Cucumis melo L. Circular procambial tissues (C‐E) in lower hypocotyls were run into the cotyledon (A) through cotyledonary-node (B, arrow for plumule). All bars = 75 µm.
한바 있으며, 체세포배 발생 능력도 유사하였다. 또한 박과 작물에서 형성된 대부분의 체세포배는 외부 형태적으로 비 정상적이며, 식물체 전환율도 매우 낮은 것으로 알려져 있 다(Ziv and Gadasi, 1986). 본 연구에서도 멜론 체세포배 중 약 74%이상이 비정상적인 형태의 특징을 갖는 것으로 주로 자엽이 융합된 나팔형인 것으로 나타나 기 연구에서 와 유사한 경향을 보였으며, 이는 자엽형성 초기과정에서 오옥신의 극성이동(Liu et al., 1993), 사이토카닌(Soh et al., 2001) 및 sucrose (Kageyama et al., (1990) 등의 영 향에 의하여 생긴 결과라 판단된다.
멜론 체세포배중에서 2개 및 합생 자엽을 갖는 체세포배 를 해부 현미경하에서 취하여 횡단면으로 관찰한 결과 2개 의 자엽을 갖는 정상적인 형태의 체세포배에서는 하배축에 서 원형모양의 전형성층 조직이 분화된 후 상배축 및 자엽 절 부위로 이동될수록 점차 2개의 독립적인 전형층 조직으 로 나뉘어져 자엽에 이르면 완전히 2개의 독립된 전형층조 직으로 분화되는 것으로 관찰되었다(Fig. 1). 반면 합생 자 엽을 갖는 체세포배의 경우에는 하배축에서 형성된 원형의 전형성층 조직이 상배축, 자엽 절 및 자엽 부위에 이르기 까지 원형으로 이어지고 있는 것으로 관찰되었다(Fig. 2).
이와 같이 2개의 자엽을 갖는 정상 체세포배나 합생자엽을 갖는 비정상적인 형태의 체세포배의 하배축에서는 동일하 게 원형의 전형층조직이 형성되어 있으나 자엽 절과 자엽 으로 갈수록 정상과 비정상 자엽을 갖는 체세포배의 전형
층조직의 분화가 다름을 보여주고 있었다. 배지에 첨가되는 여러 가지 요인 중 오옥신과 탄소원은 비정상 형태의 자엽 발생에 원인이 될 수 있으며(Soh et al., 2001; Kageyama et al. 1990), 이는 구형기의 체세포배에서 내재 오옥신의 극성이동을 교란시켜 나타나는 현상으로 알려져 있고(Liu et al. 1993), 특히 비정상적인 형태의 자엽 발생은 심장형 기 체세포배에서 전형성층조직의 분화와도 밀접한 상관관 계가 있는 것으로 보고되어 있다(Raghavan, 2004; Choi et al., 2005). 본 연구에서도 초기 체세포배의 하배축에서 전형성층 조직이 원형으로 분화된 후 자엽과 유근이 발생 될 부위로 점차 분화되어 체세포배의 외부 형태적 특징을 형성하는데 중요한 역할을 하고 있는 것으로 나타났다.
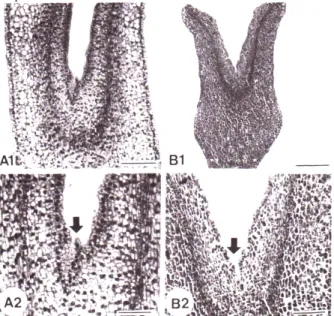
정상 체세포배와 합생 자엽을 갖는 비정상 형태의 체세 포에서 유경조직의 특징을 조사하기 위하여 종단면으로 관 찰한 결과 정상 체세포배나 비정상 체세포배 모두 다른 식 물의 체세포배에서 보여지는 유경조직의 특징인 세포질이 충만해 있는 작은 세포들로 구성되어 있거나 tunica- corpus층을 이루고 있는 전형적인 dome모양은 관찰할 수 없었고, 단지 몇 개의 세포들로 구성된 빈약한 구조를 형성 하거나 거의 흔적만 남아 있는 유경조직 특징을 보였다 (Fig. 3).
체세포배의 자엽 수와 형태적 변이는 식물체로의 재생율 을 감소시키는 중요한 요인이 될 수 있으며(Buchheim et al., 1989), 그 원인은 비정상적인 유경 조직을 형성하거나
Fig. 3. Longitudinal sections of somatic embryos with struc- turally abnormal cotyledons formed from the tissue cultures of Cucumis melo L. A1, A2: Two cotyledonary embryo with aborted plumule (arrow, bar = 90 µm); B1, B2:
Fused cotyledonary embryo with aborted plumule.
빈약한 구조가 형성되어 나타난 현상으로 이해하고 있다 (Soh et al., 2001; Dos Santos et al., 1983; Gray and Mortensen, 1987). 본 연구에서는 정상 체세포배 및 합생 자엽을 갖는 체세포배 모두 빈약한 유경조직을 형성하는 것으로 나타나 다른 식물의 합생 자엽을 갖는 체세포배의 유경조직의 특징과는 유사한 결과를 보였으나(Soh et al., 2001) 2개의 자엽을 갖는 정상적인 형태의 체세포배의 경 우에는 다른 경향을 보였다.
결론적으로 멜론 배축 절편으로부터 배발생캘러스 유도 는 2,4-D와 BA의 혼합첨가가 필요하며, 안정적인 배발생 캘러스의 증식을 위해서는 연한 노란색의 캘러스를 선발하 여 동일배지에서 계대배양하는 것이 매우 중요하다 할 수 있다. 한편 발생된 체세포배중에서 합생 자엽을 갖는 비정 상적인 체세포배의 발생 빈도가 전체 체세포배의 빈도에서 상당부분 차지하고 있을 뿐 아니라 그러한 체세포배의 유 경조직의 발달이 비정상적으로 형성되어 있으므로 식물체 로의 전환율을 감소시키는 주요 원인임을 알 수 있었다. 향 후 배발생 캘러스로부터 체세포배 발생을 통한 형질전환실 험을 성공적으로 수행하기 위해서는 정상적인 체세포배 발 생 및 전형적인 dome구조의 유경조직 형성에 대한 배양조 건 규명에 대한 연구가 선행되어야 할 것으로 판단된다.
적 요
멜론으로부터 체세포배를 얻기 위하여 유식물체의 배축 절편을 1 mg/L 2,4-D와 0.5 mg/L BA가 혼합첨가된 MS 배지 치상하여 6주 동안 배양하였다. 배축절편으로부터 연 한 노란색의 부드러운 캘러스 형성을 거쳐 형성된 체세포 배중에는 2개의 자엽을 갖는 정상적인 형태의 체세포배 (26%)와 합생 자엽을 갖는 나팔형의 비 정상적인 체세포배 (74%)가 관찰되었다. 정상 체세포배의 하배축 부위에서는 원형의 전형성층 조직이 발달되었고, 자엽 절 부위로 이동 되면서 점차 독립적으로 나뉘어져 자엽에서는 완전히 2개 의 전형성층 조직으로 분화하였다. 그러나 합생 자엽을 갖 는 나팔형의 체세포배에서는 하배축에서 발달된 원형의 전 형층조직이 자엽 절과 자엽부위에서도 원형의 전형성층 조 직으로 연결되어 있었다. 또한 2개의 자엽을 갖는 정상 및 합생 자엽을 갖는 비 정상 체세포배 tunica-corpus구조를 볼 수 없었으며 전형적인 dome구조도 관찰할 수 없었다.
이러한 결과는 체세포배에서 전형성층 조직의 분화는 자엽 형성 및 유경조직 형성과 밀접한 관계가 있는 것으로 추측 할 수 있다.
사 사
본 연구는 차세대 바이오그린21사업 (PJ008103)의 지원 하에 수행되었다.
인용문헌
Ammirato, P.V. 1987. Organizational events during somatic embryogenesis. In Green C.E. (ed.), Plant Tissue and Cell Culture, Alan R Liss, New York, USA. pp. 57-81.
Buchheim, J.A., S.M. Colburn and J.P. Ranch. 1989. Maturation of soybean somatic embryos and the transition to plantlet growth. Plant Physiol. 89:768-775.
Choi, P.S., W.Y. Soh, D.Y. Cho and J.R. Liu. 2005. Relation- ship of cotyledon number with procambium differentiation in somatic embryogenesis of Codonopsis lanceolata L.
Korean J. Plant Biotechnol. 32:135-138.
Choi, P.S., W.Y. Soh, D.Y. Cho and J.R. Liu. 1994. High frequency somatic embryogenesis and plant regeneration in seedling explants cultures of Melon (Cucumis melo L.).
Korean J. Plant Tiss. Cult. 21:1-6.
Dos Santos, A.V.P., E.G. Cutter and M.R. Davey. 1983. Origin and development of somatic embryos in Medicago sativa L.
(Alfalfa). Protoplasma 117:107-115.
Gray, D.J., D.W. McColley and M.E. Compton. 1993. High frequency somatic embryogenesis from quiescent seed cotyledon of Cucumis melo L., Cultivars. J. American Soc.
Hort. Sci. 118:425-432.
Gray, D.J. and J.A. Mortensen. 1987. Initiation and maintenance of long term somatic embryogenesis from anther and ovaries of Vitis longii “Microsperma”. Plant Cell Tiss. Org. Cult . 9:73-80.
Jelaska, S. 1974. Embryogenesis and organogenesis in pumpkin explants. Physiol. Plant. 31:257-261.
Kageyama, K., T. Komatsuda and K. Nakajima. 1990. Effects of sucrose concentration on morphology of somatic embryos from immature soybean cotyledons. Plant Tiss. Cult. Lett.
7:108-110.
Lee, K.S. and W.Y. Soh. 1993. Somatic embryogenesis and structural aberrancy of embryos in tissue cultures of Aralia cordata Thumb. Korean J. Plant Tiss. Cult. 20:77-84.
Liu, C.M., Z.H. Xu and N.H. Chua. 1993. Auxin polar transport is essential for the establishment of bilateral symmetry
during early plant embryogenesis. Plant Cell 5:621-630.
Murashige, T. and F. Skoog. 1962. A revised medium for rapid growth and bioassay with tobacco tissue cultures. Physiol.
Plant. 15:473-497.
Orezyk, W. and S. Malepszy. 1985. In vitro culture of Cucumis sativus L. stabilizing effect of glycine on leaf protoplasts.
Plant Cell Rep. 4:269-273.
Raghavan, V. 2004. Role of 2,4-dichlorophenoxyacetic acid (2,4-D) in somatic embryogenesis on cultured zygotic embryos of Arabidopsis: cell expansion, cell cycling, and morphogenesis during continuous exposure of embryos to 2,4-D. Amer. J. Bot. 91:1743-1756.
Soh, W.Y., P.S. Choi, D.Y. Cho and J.R. Liu. 2001. Plant regeneration from somatic embryos with anomalous cotyledons formed in cell cultures of Codonopsis lanceolata.
Phytomorphology Golden Jubilee Issue 327-336.
Tabei, Y., T. Kanno and T. Nishio. 1991. Regulation of organoge- nesis and somatic embryogenesis by auxin in melon (Cucumis melo L.). Plant Cell Rep. 10:225-229.
Ziv, M. and G. Gadasi. 1986. Enhanced embryogenesis and plant regeneration from cucumber (Cucumis sativus L.) callus by activated charcoal in solid/liquid double layer cultures. Plant Sci. 47:115-122.
(Received 11 March 2013 ; Revised 30 April 2013 ; Accepted 30 May 2013)