알파글루코시다아제 저해제 1-deoxynojirimycin을 생산하는 Bacillus 균주의 분리 및 동정
김현수2·이재연1·황교열1·조용석1,2·박영식1·강경돈1·성수일1,2
*
1(주)바이오토피아 부설생명과학연구소, 2수원대학교 생명과학과 Received: January 5, 2011 / Revised: February 18, 2011 / Accepted: February 19, 2011
Isolation and Identification of a Bacillus sp. producing α-glucosidase Inhibitor 1-deoxynojirimycin.
Kim, Hyun Su2, Jae Yeon Lee1, Kyo Yeol Hwang1, Yong Seok Cho1,2, Young Shik Park1, Kyung-Don Kang1, and Su Il Seong1,2*. 1R&D Center for Life Science, Biotopia Co., Ltd., Chuncheon 200-881, Korea,
2Dept. of Life Science, The University of Suwon, Hwaseong 445-743, Korea − Thirty Streptomyces sp. and 200 Bacillus sp. isolated from Korean soils and traditional foods were screened for their abilities to inhibit α-glu- cosidase and produce 1-deoxynojirimycin (DNJ). This screening identified a Bacillus sp. bacterium that strongly inhibited α-glucosidase and produced high levels of DNJ from Chungkookjang, a Korean traditional food. The bacterium was characterized in terms of its biochemical and molecular biological properties such as sugar utilization, cellular quinone composition, cell wall fatty acid composition, and 16S rDNA sequence. In addition, scanning electron microscopy was used to visualize the morphology of the bacterium. These analy- ses identified the bacterium as B. subtilis, a bacterium with Generally Recognized as Safe (GRAS) status. The selected strain was named B. subtilis MORI.
Key words: α-Glucosidase inhibitor, 1-deoxynojirimycin (DNJ), screening, bacterial identification, Bacillus subtilis MORI
서 론
우리 몸의 당대사와 glycoprotein의 생성에 관여하는 각종 glucosidase들에 대해 효소저해기능을 나타내는 polyhydroxylated alkaloid들의 항바이러스, 항암, 암전이 예방 등의 효과가 알 려짐으로써 이들 alkaloids 물질들의 산업적 응용이 보다 폭 넓게 기대되고 있다[11, 14]. 이러한 polyhydroxylated alkaloid 들은 주로 여러 종류의 식물과 미생물로부터 분리되고 있으며 대부분 단당의 구조 내에 수개의 수산기를 포함하며, 환 골격 내의 산소가 질소로 치환되어 있는 구조적 특징을 갖는다. 현 재까지 약 100여 종류가 넘는 polyhydroxylated alkaloids가 알려져 있으며 이들은 구조상의 차이에 의해 piperidines, pyrrolidines, indolizidines, pyrrolizidines, nortropanes의 다 섯 계열로 분류된다[1, 2]. Piperidine 계열은 monocycle의 6 탄당 구조를 가지며, 이 계열에 속하는 대표적인 물질로 1- deoxynojirimycin(DNJ)이 있다. DNJ는 최초로 L-sorbofuranose 로부터의 합성 또는 nojirimycin의 환원 등과 같은 화학적인 방법에 의해 얻어졌으나[7], 이후 Bacillus sp. 나 Streptomyces sp. 등과 같은 미생물의 배양액, 또는 뽕나무(Morus alba L.)
나 누에(Bombyx mori L.)의 추출물로부터도 정제되었다[3, 4, 6, 8, 10].
DNJ는 포유동물의 소장에 존재하는 α-glucosidase의 활성 을 저해함으로써 소장에서 혈액으로의 포도당 유입을 억제 하는데, 이런 특성 때문에 DNJ는 식후 고혈당증을 유발하 는 인슐린 비의존형 당뇨병 환자들의 혈당강하제로 이용되 고 있다[16]. 또한 DNJ는 α-glucosidase 활성 저해기능 외 에 간염바이러스나 AIDS 바이러스와 같은 외막 형성 바이 러스의 증식을 억제함으로써 이를 응용한 바이러스성 질병 치료제로의 이용가능성도 제기되고 있다[15]. 따라서 이와 같은 DNJ의 산업적 활용성을 감안할 때 만약 DNJ의 대량 생산이 가능하다면 이 물질의 산업적인 응용범위는 더욱 크 게 확대될 것으로 기대된다.
이에 본 연구진은 토양으로부터 분리한 Streptomyces sp.
30여 균주들과 한국전통식품에서 분리한 Bacillus sp. 200여 균주들을 대상으로 α-glucosidase의 저해활성이 우수하며 동 시에 DNJ의 생산이 높은 유용균주를 선발하고 나아가 이 균 주에 대한 생화학적 및 분자생물학적 동정을 통해 GRAS로 간주되는 B. subtilis 균주임을 확인하였다. 따라서 본 연구 를 통해 확보한 균주는 부가적인 대사공학적 연구를 통하여 건강기능성식품 소재나 의약품 소재 생산 등을 위한 DNJ의 대량생산 시스템 구축에 활용될 수 있을 것이다.
*Corresponding author
Tel: +82-31-220-2483, Fax: +82-31-220-2483 E-mail: [email protected]
재료 및 방법 균주 및 배양조건
토양으로부터 분리한 Streptomyces sp. 30여 균주들과 된 장, 청국장 등 한국전통식품들에서 분리한 Bacillus sp. 200 여 균주들을 사용하였다. 각각의 균주들은 1% glucose, 0.5% soybean meal, 0.3% yeast extract가 함유된 25 ml 액체배지에서 37oC, 150 rpm으로 5일간 진탕배양 하였고, 100oC에서 배양액을 5분간 열처리한 후 원심분리(9,200×g, 10 min)하여 얻어진 상등액을 α-glucosidase의 효소활성 저 해능 검토를 위한 시료로 사용하였다.
균주의 선발기준은 α-glucosidase의 효소활성을 저해함과 동시에 DNJ를 생산하는 균주로 하였으며, α-glucosidase inhibitor를 생산하는 표준균주로 Bacillus subtilis (KCCM 11314), Bacillus amyloliquefaciens (KCCM 40764), Streptomyces subrutilus (KCCM 40337), Streptomyces lavendulae 3균주 (KCCM 32799, KCCM 12294, KCCM 12293) 및 Streptomyces nojiriensis (KCCM 12307) 등을 사용하였다.
α-Glucosidase 효소원의 제조
도살 직후 적출된 돼지소장의 미세융모를 유리편을 이용 해 조심스럽게 긁어낸 후 0.1 M potassium phosphate buffer (pH 6.8)에 현탁한 후 원심분리(9,200×g, 10 min)하여 상등 액을 얻었다. 이 상등액에 80% ammonium sulfate를 첨가하 여 단백질들을 침전시킨 후 원심분리(9,200×g, 20 min)하 여 얻은 침전물을 투석막(M.W. 12,000 Da)(Sigma, USA)을 이용하여 4oC에서 12시간 동안 탈염한 후 -70oC에서 동결 건조하여 α-glucosidase 효소원으로 사용하였다. 효소원 내 의 단백질 함량은 Bradford의 방법[12]에 의해 정량하였다.
제조된 α-glucosidase 효소원의 효소활성 검토
돼지소장으로부터 얻어진 α-glucosidase 효소원을 전기 영동하여 단백질을 분리하고 기질-효소 반응을 통해 α- glucosidase의 존재를 확인하였다. 전기영동은 Davis 법[5]에 따라 Native-PAGE 하였으며, 7.5% polyacrylamide gel을 사용하였다. 전기영동 후 동일한 효소원을 대상으로 전개한 gel를 양분하여 한쪽은 Coomassie brilliant blue R-250으로 단백질 염색을 실시하였고, 다른 한쪽은 2.3 mM 4- methylumbelliferyl-α-D-glucose를 기질로 효소반응 시킨 후 ultraviolet transilluminator를 이용하여 α-glucosidase 존재여 부를 확인하였다.
α-Glucosidase 효소원 내의 maltase, sucrase, cellobiase 및 lactase 등 disaccharidase 들의 효소 활성은 TLC 분석을 통해 같은 방법으로 확인하였다. 먼저 0.1 M potassium phosphate buffer (pH 6.8)에 1 mg/ml이 되도록 녹인 α- glucosidase 효소원 100 µl와 2당류 기질로서 10 mM의
maltose, sucrose, cellobiose 혹은 lactose 등을 각 400 µl씩 가하여 37oC에서 45분간 효소반응 시킨 후 90oC에서 5분간 열처리로 반응을 정지시켰다. 각각의 반응액을 TLC plate (Merck, Silica gel F254)상에 10 µl씩 점적하고 butanol- propanol-acetic acid-water (5 : 3 : 3 : 1.5, v/v/v/v)의 혼 합용매에서 전개하였으며, 이 때 기질과 같은 농도의 D- glucose(Sigma, USA)를 표준물질로 함께 전개하였다. 전개가 끝난 TLC plate는 60oC oven에서 완전히 건조한 후 aniline- diphenylamine reagent (2 ml aniline, 2 g diphenylamine, 10 ml 85% phosphoric acid, 100 ml methanol)를 분사하 고 104oC에서 30분간 발색하여 glucose의 생성여부를 확인 하였다.
α-Glucosidase 효소원의 활성 분석
α-Glucosidase 효소원을 0.1 M potassium phosphate buffer (pH 6.8)를 이용하여 연속 희석액을 조제하고 각 연속희석 액에 12 mM p-nitrophenyl-α-D-glucopyranoside를 1 ml을 가하여 37oC에서 45분간 반응시킨 후 200 mM sodium carbonate 1 ml을 첨가하여 반응을 정지시켰다. 이 효소 반 응액의 흡광도(405 nm)를 측정하였으며, 효소역가는 1분 동 안 1 µM의 p-nitrophenol이 생성된 양을 1 unit으로 간주하 여 산정하였다. 이때 Molar extinction coefficient(ε)는 1.78
×104 M−1ㆍcm−1이었다.
Maltase 저해 우수 균주선발
돼지 소장으로부터 분리한 효소추출액을 0.1 M potassium phosphate buffer (pH 6.8)에 1 mg/ml가 되도록 용해시킨 후, 이 용액 100 µl에 10 mM maltose 400 µl, 각 균주들 의 열처리된 배양상등액 200 µl를 가하여 37oC에서 45분간 반응시킨 후 90oC에서 5분간 열처리로 반응을 정지시켰다.
이 반응액을 TLC plate (Merck, Silica gel F254)에 10 µl씩 점적하여 butanol-propanol-acetic acid-water(5 : 3 : 3 : 1.5, v/v/v/v) 혼합용매를 통해 전개한 후 aniline-diphenylamine reagent로 발색하여 glucose의 생성 정도에 따라 α-glucosidase 효소원내에 존재하는 α-glucosidase 중에서 maltase 저해능 이 우수한 균주를 선발하였다.
DNJ 생산 우수 균주선발
Maltose 분해 억제능이 우수한 균주들을 대상으로 각 균 배양액을 HPLC (ThermoQuest Co., U.S.A.)로 분석하여 균의 DNJ 생산 여부를 조사하였다. HPLC 분석을 위한 시료 조제 는 다음과 같이 준비하였다. 균 배양액 10 µl에 0.4 M borate buffer (pH 8.5) 10 µl와 9-flurenylmethyl chloroformate (FMOC-Cl) 20 µl를 넣어 20oC에서 20분간 반응시킨 후 0.1 M의 glycine 10 µl를 첨가하여 반응을 정지시켰다. 이 반응 액에 0.1% acetic acid 950 µl를 첨가하여 1 ml로 맞춘 후 column에 10 µl씩 주입하였다. Column은 Phenomenex
Luna C18 (4.60×250 mm I.D., 5-µm)을 사용하였고, 용매 로 acetonitrile-0.1% acetic acid (1 : 1, v/v)를 사용하여 용출하였으며, 용출속도는 1 ml/min, 검출기는 FL3000 fluorescence detector (excitation 254 nm, emission 322 nm), 분석 프로그램은 ChromQuestTM (ThermoQuest Co., USA)을 각각 사용하였다.
Polyhydroxylated alkaloids의 표준시료로서 DNJ는 Sigma 사로부터 구매한 표준품을 사용하였으며, 그 밖에 2-O-α-D- galactopyranosyl-1-deoxynojirimycin, 1,4-dideoxy-1,4-imino- D-arabinitol, 1,4-dideoxy-1,4-imino-(2-O-β-D-glucopyranosyl) -D-arabinitol, fagomine, 3-epi-fagomine, calystegin B2 그 리고 N-methyl-1-deoxynojirimycin 등은 농촌진흥청 농업생 물부 활성물질연구실로부터 분양받아 사용하였다[9].
DNJ 생산 균주의 동정
최종 선발된 균주는 다양한 생화학적 및 분자생물학적 방 법에 의해 균주 동정을 하였다. 생화학적 동정으로는 API 50 CHB® Kit (BioMerieux, France)를 이용하여 균의 탄소 원 이용능을 분석하였고, gas chromatography (HP 6890, U.S.A.)로 세포내 지방산을 분석하였고 그리고 HPLC를 이 용하여 quinone을 분석하였다[13]. 분자생물학적 동정으로는 균의 16S rDNA의 염기서열을 비교하였으며 주사전자현미 경(JSM-5600, Japan)에 의해 분리균주의 형태 및 크기를 관 찰하였다.
결과 및 고찰
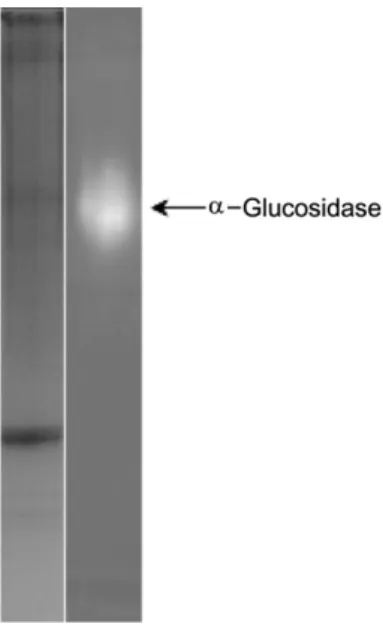
돼지소장유래 α-glucosidase 효소원의 효소 활성 분석 돼지소장으로부터 분리하여 동결 건조한 α-glucosidase 효 소원의 단백질 함량은 약 30%(w/w)로써 일반적인 효소의 구성성분인 단백질의 존재가 확인되었으며, Native-PAGE 분 석결과 Fig. 1의 왼쪽 편 gel에서는 단백질 함량이 적어서 α-glucosidase가 잘 보이지 않았으나, 오른쪽 편 gel에서는 α-glucosidase에 특이적으로 작용하는 기질인 4-methylum- berlliferly-α-D-glucoside을 사용하여 형광으로 검출하였을 때 돼지소장의 효소추출액내에 α-glucosidase가 존재함을 뚜 렷하게 확인할 수 있었다. 또한, 효소추출액의 이당류들에 대한 α-glucosidase 효소활성을 조사한 결과, maltose와의 반 응에서만 glucose 생성이 확인되었다(Fig. 2). 이상의 결과, 돼지소장 추출액 내에는 α-glucosidase가 함유되어 있고, 이 α-glucosidase는 maltose에 대해 기질특이성을 보이고 있음 을 확인하였다. 돼지소장유래 α-glucosidase 효소원의 효소 역가를 측정한 결과 효소단백질 1 mg 당 7.7 unit의 α- glucosidase 활성을 보유하고 있는 것으로 나타났다.
α-Glucosidase inhibitor 및 DNJ 생산균주의 선발 DNJ 생산균주 선발은 해당 균주 배양액이 첨가된 α-
glucosidase 효소반응액을 TLC로 분석한 결과 우선 α- glucosidase 저해활성에 의해 glucose의 생성량이 대조군에 비하여 적거나 혹은 생산이 되지 않는 균주들을 1차적으로 Fig. 1. α-Glucosidase zymogram of porcine intestinal extract.
The extract was separated by 7.5% native PAGE and stained with Coomassie brilliant blue R-250 (left) or 4-methylumbelliferyl α-D- glucoside (right).
Fig. 2. TLC chromatogram following the incubation of por- cine intestinal extract and disaccharides. The 500µl reaction mixture contained 0.2 mg/ml of porcine intestinal extract and 8 mM of substrate [maltose (lane 1), sucrose (lane 2), cellobiose (lane 3) or lactose (lane 4)] in 0.1 M potassium phosphate buffer, pH 6.8. The reaction mixture was incubated at 37oC for 45 min.
Ten microliters of the reaction mixture was separated by TLC using butanol-propanol-acetic acid-water (5 : 3 : 3 : 1.5, v/v/v/v) as a solvent. The separation of 8 mM of D-glucose is shown in lane C.
선발하였다. 이들 선발된 균주들 가운데 그 배양액을 HPLC 로 분석하여 DNJ 생산능이 상대적으로 우수한 균주를 2차 로 선발하였다. 그 결과 α-glucosidase 활성 억제에 의해 glucose의 생성이 상대적으로 약하며 동시에 DNJ의 생산이
확인된 Streptomyces sp. 4 균주와 Bacillus sp. 10 균주를 후보 균주로 선발하였다. 이렇게 선발된 균주들 가운데 한 국전통식품인 청국장으로부터 분리되었으며 α-glucosidase 활성 저해능이 가장 우수하고(Fig. 3) DNJ 생산성이 높은
Fig. 3. TLC chromatograph showing α-glucosidase (maltase)- inhibiting effects of the culture broth from several microorgan- isms. The 700 µl reaction mixture contained 0.14 mg/ml of porcine intestinal extract, 5.7 mM of maltose, and 200 µl of culture broth [lane 1; control (non-treated culture broth), lane 2; Streptomyces lavendulae subsp. lavendulae KCCM 12293, lane 3; Bacillus amy- loliquefaciens KCCM 40764, lane 4; Streptomyces sp. SS-0247, lane 5; Streptomyces sp. SS-4299, lane 6; B. subtilis MORI]. The reaction mixture was incubated at 37oC for 45 min and 10 µl was separated by TLC as described in Fig. 2.
Fig. 4. HPLC separation of the culture broth of B. subtilis MORI. B. subtilis MORI was cultured at 37oC for 5 days with shaking at 150 rpm in culture medium containing 1%(w/v) glu- cose, 0.5%(w/v) soybean meal, and 0.3%(w/v) yeast extract. The dotted line represents the separation of several standard alkaloid compounds: peak A; 2-O-α-D-galactopyranosyl-1-deoxynojirimy- cin, peak B; 1,4-dideoxy-1,4-imino-(2-O-β-D-glucopyranosyl)-D- arabinitol, peak C; 1-deoxynojirimycin, peak D; 3-epi-fagomine, peak E; fagomine/1,4-dideoxy-1,4-imino-D-arabinitol, and peak F;
calystegin B2.
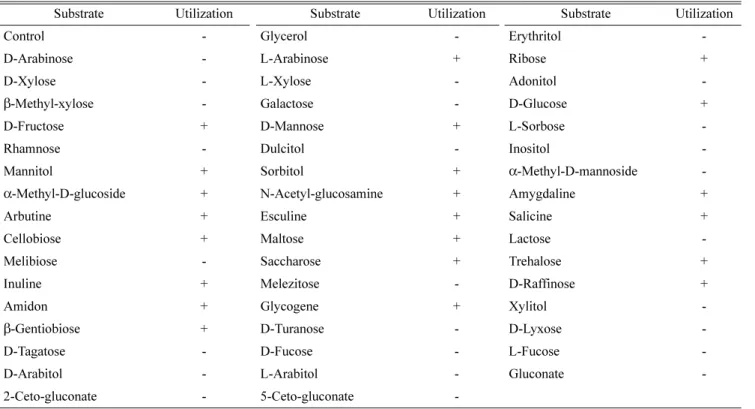
Table 1. Sugar utilization of B. subtilis MORI.
Substrate Utilization Substrate Utilization Substrate Utilization
Control - Glycerol - Erythritol -
D-Arabinose - L-Arabinose + Ribose +
D-Xylose - L-Xylose - Adonitol -
β-Methyl-xylose - Galactose - D-Glucose +
D-Fructose + D-Mannose + L-Sorbose -
Rhamnose - Dulcitol - Inositol -
Mannitol + Sorbitol + α-Methyl-D-mannoside -
α-Methyl-D-glucoside + N-Acetyl-glucosamine + Amygdaline +
Arbutine + Esculine + Salicine +
Cellobiose + Maltose + Lactose -
Melibiose - Saccharose + Trehalose +
Inuline + Melezitose - D-Raffinose +
Amidon + Glycogene + Xylitol -
β-Gentiobiose + D-Turanose - D-Lyxose -
D-Tagatose - D-Fucose - L-Fucose -
D-Arabitol - L-Arabitol - Gluconate -
2-Ceto-gluconate - 5-Ceto-gluconate -
+; reaction, - ; non-reaction.
Bacillus 속의 한 개 균주를 최종 목적균주로 선발하였다.
DNJ를 체액 중에 고농도로 함유하고 있는 것으로 알려진 누 에(Bombyx mori L.)의 종명에 근거하여 MORI 균주로 명명 하였으며 아래 기술한 일련의 동정을 실시하였다(Fig. 4).
선발균주의 동정
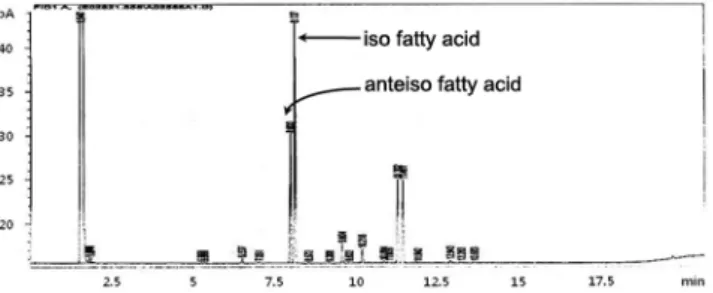
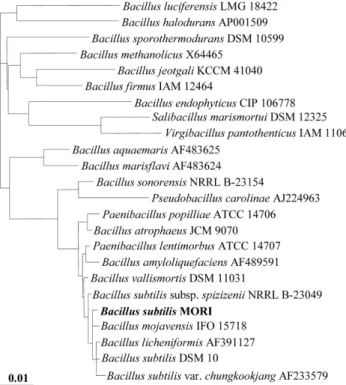
최종 선발한 MORI 균주에 대한 생화학적 및 분자생물학 적 동정을 하였다. 먼저 API kit를 이용한 49 종류의 당 기 질에 대한 이용능 시험(Table 1)과 이에 대한 API 분석프로그 램(Biomerieux, France)의 동정 결과, MORI 균주는 Bacillus 속에 속하는 균주로 확인되었고, 광학현미경상에서 Bacillus 균의 형태적 특징인 포자형성이 관찰되었다. 또한 지방산 분 석결과 MORI균은 세포내 주 지방산으로서 iso- 및 anteiso- branched 구조가 많이 함유되어 있으며, Bacillus subtilis와 75%의 높은 유사성을 보였고(Fig. 5), quinone 분석에서는 표준균주로 사용한 Bacillus subtilis (KCCM 11316) 균주의 menaquinone-7과 일치하는 것으로 확인되었다(Fig. 6). 이상 의 생화학적 동정 외에 16S rDNA의 염기서열을 비교 분석 한 결과 MORI 균주는 그람양성 세균 중 Bacillus 속에 속 하는 Bacillus subtilis와 99.0%의 유사성을 갖는 것으로 나 타났다(Fig. 7). 한편, 균주들의 16S rDNA 염기서열을 기준 으로 dendrogram을 작성하고 균주들 간의 유연관계를 조사 한 결과 MORI 균은 B. subtilis에 속하는 균주들 외에 B.
licheniformis, B. mojavensis 등의 균주들과도 매우 가까운 유연관계에 있음을 알 수 있었다(Fig. 8). 또한 주사전자현 미경 관찰로부터 MORI 균이 Bacillus 균의 형태적 특성인 간균임도 확인할 수 있었다(Fig. 9). 따라서 한국전통식품인 청국장으로부터 분리된 MORI 균주는 이상의 생화학적 및 분자생물학적 균 동정법을 통하여 GRAS(generally recognized as safe)로 분류되는 안전성이 검증된 Bacillus subtilis 균주 임이 확인되었다. 따라서 MORI 균주는 전혀 인체에 유해성 이 없는 식품으로 적용이 가능한 균주로써 건강기능식품으 로의 개발이 가능하다고 할 수 있다.
앞으로 B. subtilis MORI 균주가 DNJ와 함께 생산하는 여타 수종의 polyhydroxylated alkaloids 성분들에 대해서도 이들 물질의 분리와 정제, 생화학적 특성 및 기능성 규명에 대한 연구가 필요할 것으로 사료된다. 아울러 선발된 MORI 균주의 DNJ 생산성을 높이기 위한 균주개량 및 배양조건의 최적화 등에 관한 연구도 계속되어야 할 것이다. 나아가 DNJ 의 생화학적 합성경로 및 DNJ 생성 관련 유전자가 밝혀진 Fig. 5. Fatty acid composition of a whole cell extract of B. sub-
tilis MORI using gas chromatography.
Fig. 6. HPLC chromatogram of isoprenoid quinone extract from B. subtilis MORI.
Fig. 7. 16S rDNA sequence of B. subtilis MORI.
다면 보다 새로운 방식의 경제성 있는 DNJ 생산도 가능할 수 있을 것으로 기대된다.
요 약
토양으로부터 분리한 Streptomyces sp. 30여 균주들과 한
국전통식품에서 분리한 Bacillus sp. 200여 균주들로부터 α- glucosidase 의 활성을 저해하고 동시에 DNJ를 생산하는 유 용균주를 선발하였다. 실험결과 한국전통식품인 청국장으로 부터 α-glucosidase 저해능이 높고 DNJ 생산능이 우수한 한 개의 균주를 선발하였다. 이 균주의 동정을 위하여 API kit 에 의한 균의 당 이용능 분석, HPLC와 GC에 의한 균체의 quinone 및 지방산 분석 등과 함께 균의 16S rDNA 염기서 열을 분석하였으며 주사전자현미경에 의해 균주의 형태적 특성을 관찰하였다. 그 결과 본 연구를 통해 선발한 균주는 건강기능성식품 개발 등에 적용할 수 있는 GRAS에 속하는 균주임을 확인하여 B. subtilis MORI로 명명하였다.
Acknowledgment
This study was supported in part by the MKE (The Minis- try of Knowledge Economy), Korea, under the Leading Industry Development for Gangwon Economic Region pro- gram (70007579).
R
EFERENCES1. Asano, N., A. Kato, K. Oseki, H. Kizu, and K. Matsui. 1995.
Calystegins of Physalis alkekengi var. francheti (Solanaceae).
Structure determination and their glycosidase inhibitory activities. Eur. J. Biochem. 229: 369-376.
2. Asano, N., R. J. Nash, R. J. Molyneux, and G. W. Fleet.
2000. Sugar-mimic glycosidase inhibitors: natural occurrence, biological activity and prospects for therapeutic application.
Tetrahedron 11: 1645-1680.
3. Asano, N., T. Yamashita, K. Yasuda, K. Ikeda, H. Kizu, Y.
Kameda, A. Kato, R. J. Nash, H. S. Lee, and K. S. Ryu.
2001. Polyhydroxylated alkaloids isolated from mulberry trees(Morus alba L.) and silkworms(Bombyx mori L.). J.
Agric. Food. Chem. 49: 4208-4213.
4. Bradford, M. M. 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 72: 248- 254.
5. Davis, B. 1964. Disc electrophoresis-II. Method and application to human serum proteins. Ann. New York Acad.
Sci. 121: 404-427.
6. Inouye, S., T. Tsuruoka, T. Ito, and T. Niida. 1966. The structure of nojirimycin, a piperidinose sugar antibiotic. J.
Antibiot. Ser. A. 19: 288-292.
7. Inouye, S., T. Tsuruoka, T. Ito, and T. Niida. 1968. Structure and synthesis of nojirimycin. Tetrahedron. 24: 2125-2144.
8. Ishida, N., K. Kumagai, T. Niida, T. Tsuruoka, and H.
Yumoto. 1967. A new antibiotic, nojirimycin. II. Isolation, characterization and biological activity. J. Antibiot. Ser. A.
20: 66-71.
9. Kim, J. W., S. U. Kim, H. S. Lee, I. S. Kim, M. Y. Ahn, and K. S. Ryu. 2003. Determination of 1-deoxynojirimycin in Fig. 8. Phylogenetic analysis of the 16S rDNA sequence from
B. subtilis MORI and those of 23 other Bacillus sp. and related bacteria. The tree was constructed by using the neighbor-joining method based on partial 16S rDNA sequences. The scale bar indi- cates the 0.01% nucleotide difference.
Fig. 9. Scanning electron micrograph of B. subtilis MORI. The bar indicates 1 µm.
Morus alba L. leaves using derivatization with 9- fluorenylmethyl chloroformate followed by reversed-phase high-performance liquid chromatography. J. Chromatogr. A.
1002: 93-99.
10. Murao, S. and S. Miyata. 1980. Isolation and characterization of a new trehalase inhibitor, S-GI. Agric. Biol. Chem. 44:
219-221.
11. Olden, K., P. Breton, K. Grzegorzewski, Y. Yasuda, B. L.
Gause, J. K. Cha, V. A. Miles, S. A. Newtown, and S. L.
White. 1991. The potential importance of swainsonine in therapy for cancers and immunology. Pharmacol. Ther. 50:
285-290.
12. Schmidt, D. D., W. Frommer, L. Müller, and E. Truscheit.
1979. Glucosidase-inhibtoren aus Bazillen. Naturwis- senshaften. 66: 584-585.
13. Shin, Y. K, J. S. Lee, C. O. Chun, H. J. Kim, and Y. H. Park.
1996. Isoprenoid quinone profiles of the Leclercia adecarboxylata KCTC 1036. J. Microbiol. Biotech. 6: 68- 69.
14. Taylor, D. L., P. Sunkara, P. S. Liu, M. S. Kang, T. L.
Bowlin, and A. S. Tyms. 1991. 6-O-Butanoylcastanospermine (MDL 28,574) inhibits glycoprotein processing and the growth of HIVs. AIDS 5: 693-698.
15. Watson, A. A., G. W. Fleet, N. Asano, R. J. Molyneux, and R. J. Nash. 2001. Polyhydroxylated alkaloids-natural occurrence and therapeutic applications. Phytochemistry 56:
265-295.
16. Yagi, M., T. Kouno, Y. Aoyagi, and H. Murai. 1976. The structure of moranoline, a piperidine alkaloid from Morus species. Nippon Nogeikagaku Kaishi. 50: 571-572.