1)
†To whom correspondence should be addressed.
National Institute of Forest Science E-mail: [email protected]
멸종위기 식물인 갯봄맞이 최남단 군락의 식생구조
임정철・이철호
*・ 김의주
**・ 최병기
***,†국립습지센터
*동의대학교 분자생물학과
**공주대학교 생명과학과
***국립산림과학원
Vegetation Structure and Management Strategies of Glaux maritima var.
obtusifolia Community on the Southernmost Distribution Area in Korea
Jeong Cheol Lim・Cheol Ho Lee
*・Eui Ju Kim
**・Byoung Ki Choi
***,†National Wetlands Center, NIER
*
Department of Molecular Biology, Dongeui University
**
Department of Life Science, Kongju National University
***
National Institute of Forest Science
(Received : 08 September 2017, Revised: 13 October 2017, Accepted: 18 October 2017)
요 약
갯봄맞이는 한국에서 매우 제한적인 분포를 보여 멸종위기종으로 지정・보호되고 있다. 본 연구는 갯봄맞이의 남방한 계지역에 해당하는 당사해안습지의 식생 다양성을 확인하고 식생결정 요인과 서식처 중요성을 평가하는데 목적을 두 고 있다. 식생분류를 위해 Z.-M. 학파의 식물사회학적 방법을 활용하였으며, 환경요인과 식생과의 관계 분석을 위해 수리통계분석을 수행하였다. 연구결과 당사해안습지는 갯봄맞이-지채군락 (3개 하위군락), 갯꾸러미풀s.l.군락, 기장대 풀군락, 갈대-꼬마부들군락의 4개 단위식생으로 구성되어 있었다. 이들 식생의 주요 결정요인은 수분환경과 토심, 수 위교란정도, 식생고, 군락구조 등인 것으로 확인되었다. 갯봄맞이는 갯봄맞이-지채군락의 전형아군집에서 가장 우세하 게 생육하는 것으로 확인되었으며, 인접한 일본의 남방한계지역과도 유사한 종조성 및 우점상황을 보이는 것으로 나타 났다. 당사해안습지는 식물지리 및 식생지리학적 측면에서 큰 의미를 지니는 입지이며, 다양한 종의 피난처로서 기여 하고 있는 것으로 평가되었다. 당사해안습지의 가치를 다양한 관점에서 정밀하게 파악하고 보호 및 관리하기 위한 보 전전략의 수립이 요구된다.
핵심용어 : 군락지리, 기후변화, 식물지리, 최남단분포, 피난처
Abstract
Glaux maritima var. obtusifolia is distributed in very limited areas in South Korea and thus designated and protected as an endangered species. This study aimed to identify the diversity of vegetation in the Dangsa coast wetlands, the southern limit of G. maritima var. obtusifolia and to assess determinants of the vegetation and the importance of habitats. The phytosociological method of the Zürich-Montpellier School was used for vegetation classification and mathematical and statistical analyses were also conducted to analyze environmental factors and their relationship with the vegetation. The results of this study showed that there were 4 vegetation units in the Dangsa coast wetlands:
Glaux maritima var. obtusifolia-Triglochin maritimum community (included three subcommunities), Puccinellia nipponica s.l. community, Beckmannia syzigachne-Isachne globosa community and Typha laxmannii-Phragmites communis community. It was also found that major determinants of the vegetation include moisture environment, soil depth, water level disturbance, vegetation height, community structure, etc. Glaux maritima was identified to grow most dominantly in the typicum subassociation of Glaux. maritima var. obtusifolia-Triglochin maritimum community, and the species compositions and dominant situations were observed to be similar to those in the southern limit in Japan, adjacent to South Korea. The assessment results indicated that the Dangsa coastal wetlands have a significant meaning from phytogeographical and syngeographical aspects, and contribute as a shelter for
1. 서 론
유라시아 대륙과 북미의 한대 및 온대 해안지역에 넓게 분포 하는 갯봄맞이(
Glaux maritima sensu lato
)는 2012년부터 국내에서 제한된 분포와 서식처 독특성으로 인해 멸종위기종 으로 지정하여 보호되고 있다(Hu and Kelso, 1996; NIBR, 2012). 국내에 분포하는 종은 본종인G. maritima
의 변종으로 상대적으로 ‘잎의 끝이 둔한(obtusifolia
)’ 종이다. 그러나 분류 학적 논의와 별개로 갯봄맞이(G. maritima
var.obtusifolia
)는 본종과 유사한 서식처 환경에 분포하고 있으며, 식생을 형성함 에 있어 동질한 종조성으로 집단을 구성하고 있으므로 식물사 회학적(phytosociological)으로 동일 또는 대응단위식생에 위 치시키고 있다(Miyawaki and Ohba, 1965). 연구지역 내 갯 봄맞이의 서식은 2012년 울산지역 희귀동식물 자생지 및 서식 지 조사과정에서 발견되었으며, 현재까지의 국내 식물분포 연구결과를 고려할 때 한반도의 최남단 분포지로 고려되고 있다(Shin et al., 2012). 보전생태학적으로 후방가장자리개체 군(rear edge population)은 대상종의 분포중심지에 비해 개체 군의 훼손 및 서식처의 질적 쇠퇴에 대해 보다 민감하게 반응하 며, 소멸 가능성이 높은 것으로 고려된다(Hampe and Petit, 2005; O'Grady et al., 2004). 또한 이들 개체군은 지구기후변 화의 살아있는 흔적으로 고려되며, 고기후와 연계된 지역생태 계를 이해하고 해석하는데 중요한 생물증거로 활용하고 있다.나아가 개체군의 유전자급원(gene pool)이 상대적으로 극단적 기후에 적응된 메타개체군으로, 전체 집단 가운데 싱크개체군 (sink population)에 비해 독립된 유전자 선택으로 인해 이탈 된 적응유전자의 대립인자 획득으로 종유전적 가치에서 상향 평가되기도 한다(Kim et al., 2012).
최근 Kim et al.(2016)에 의해 한반도 동해안 일대에 분포하 는 갯봄맞이군락지에 관한 연구가 이루어졌으며, 4개 지역의 개체군 특성과 유기물에 의한 개체군 활력 비교, 월동아, 열매 및 종자생산 능력과 발아특성 등에 대한 종합적인 고찰이 의미 있게 수행되었다. Kim et al.(2016)은 개체군을 구성하게 하는 주요 환경요인으로 염분과 주기적인 침수 그리고 낮은 토양층 으로 기술하였으며, 경쟁관계 식물의 억제를 갯봄맞이 분포에 중요요인으로 제시하였다. 이러한 특성은 유럽(Adam, 1981;
Burnside et al., 2007; Ellenberg, 2009; Kaplan, 2012)과 일본(Takagi and Shirai, 2013), 북미(Hitchcock, 1973;
Brotherson, 1984; Heidel et al., 2002) 등에서 공통적으로 인식되고 있으며, 해안지역 뿐 아니라 내륙지역인 중앙아시아 (Hilbig, 2003; Markovskaya, 2014; Sergienko, 2012), 몽골 (Miehe, 2009), 중국(Minggagud and Yang, 2013) 등에서도 유사한 경향을 보이는 것으로 알려져 있다.
Bae et al.(2014)은 갯봄맞이 종자의 휴면타파와 기내 증
식에 관한 연구에서 6주 이상의 저온처리가 개체에 있어 높은 발아효율을 보이는 것으로 확인하였으며, 개체증식을 위한 기내배양 조건들에 대해 고찰하였다. 기내배양 조건 중 마디배양의 경우 특정절간의 배양이 상대적으로 높은 효율을 보임을 확인했으며, 담수환경에서도 염수환경과 동 일한 생활환을 나타내는 것으로 기재한 바 있다(Bae, 2016). 그 외 갯봄맞이에 관한 국내연구들은 종의 분류학 적 비교, 종 지리 분포 및 개체군의 크기 등에 대한 언급에 그치고 있다(Oh, 1980; Baek, 2015; Son et al., 2011;
Shin et al., 2012). 그러나 가장 시급하게 보호가 필요한 자생서식지에 대해서는 멸종위기종인 갯봄맞이의 종보호 측면에서의 가치 외에 이들 종으로 구성된 식생의 분류 및 서식처 독특성에 대한 접근이 상대적으로 미흡한 실정이다.
본 연구는 국내뿐 아니라 전세계적인 최남단 분포지역 중 하나인 당사해안 일대의 갯봄맞이군락지에 대해 식물사회 학적 군락분류 및 군락생태, 군락동태, 환경결정요인 등을 밝혀, 본 입지의 서식처 특성과 중요성을 밝히는데 목적을 두고 있다. 또한 연구지역 일대의 정밀현존식생도를 통해 멸종위기종을 포함한 지역 식생의 분포현황을 기록하고, 식 생 구성종과 함께 주변 지역의 출현종을 파악하여 지역 종 급원(local species pool)을 확보하고자 하였다. 이들 정보 는 연구지역 일대의 서식처 특성에 대한 이해와 함께 직간 접적인 위협요인을 도출하여, 보전생태학적 관리방안을 제 시하는데 의미 있는 기초정보를 제공할 수 있을 것으로 기 대된다.
2. 연구 지역 및 방법
2.1 연구 지역
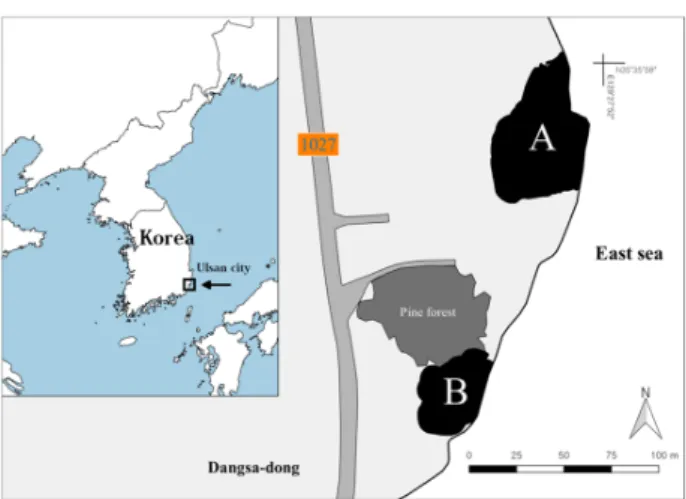
본 연구는 갯봄맞이군락이 분포하는 당사해안습지와 그 일대에 대하여 조사를 실시하였다. 연구지역은 울산광역시 북구 당사동의 해안에 위치한 습지(이하 당사해안습지 통 일)이며, 35︒35’56”N, 129︒27’49”E ~ 35︒35’58”N, 129︒27’51”E 일대에 위치한다. 당사해안습지는 두 개 습 지로 구분되며, 각 습지의 면적은 2,650 ㎡와 1,326 ㎡으로 수심 0.5 m 미만의 소형 습지이다(Fig. 1). 습지의 집수역도 습지 주변공간의 좁은 면적에 제한되는데, 습지 서측에 위치 한 용출소의 존재로부터 연중 적정 수심이 유지되는 습지를 형성하고 있다(KNA, 2012). 습지에서 바다까지의 거리는 20 m이하로서, 일반적 수위 시에는 습지로의 해수 유입이 발생되지 않으나, 태풍을 비롯한 높은 파도가 발생할 시에는 습지로 직접적인 해수의 유입이 가능한 지역이다. 습지는 동측으로 완만한 평탄지형을 형성하고 있으며, 동북측으로 흐르는 유출수는 무식생대를 거쳐 바다로 이어진다.
diverse species. It is required to establish conservation strategies to accurately determine the value of the wetlands of the Dangsa coast from various perspectives, and to protect and manage them.
Key words : Climatic Change, Phytogeography, Refugium, Southern Limited Distribution, Syngeography
Fig. 1. Study area (area, A: 2,650m2
and B: 1,326m
2)
연구지역의 연평균기온은 14.1 ℃, 최저기온 9.8 ℃, 최고 기온 19.2 ℃이며, 강수량은 1,277.1 mm이다(KMA, 2016).식생지리적으로 남부해안요소와 동해안요소가 만나는 지역 이며, 냉온대와 난온대의 전이대로서 쿠로시오난류의 영향 을 받는 동해안남부형의 생물기후구계에 위치한다(Moon, 2006; Kim and Lee, 2007; Choi, 2013)
2.2 연구 방법
식생연구는 Z-M학파의 식물사회학적 방법으로 현장 식생 조사가 이루어졌으며(Braun-Blanquet, 1965), 피도는 9등급 의 순차척도인 변환통합우점도를 활용하여 산출하였다 (Westhoff and van der Maarel, 1978). 단위식생(syntax)추출 은 전통적인 표조작 분류법(Becking, 1957)과 수리통계분석을 동시에 고려한 통합분류방법으로 수행하였다(Kim and Lee, 2007). 식생 구성종의 군락 내 기여도를 파악하기 위해 출현빈 도와 피도를 바탕으로 각 종의 상대기여도(relative Net Contribution Degree)를 도출하였으며, 단위식생명은「국제 식생명명규약」(Weber et al., 2000)에 따랐다. 식물사회학적 분류 체계를 확립하고 단위식생들 간의 질적, 양적 상관관계와 유의성 분석을 위해 [SYNTAX 2000](Podani, 2001)의 PCoA(Principal coordinates analysis) 좌표결정과 SPSS를 통 한 상관관계분석이 이루어졌다. 일본 최남단 분포지와의 식생 비교를 위해 대상지역의 식생자료를 확보하고 두 지역 간 군락 종조성을 바탕으로 수리분석을 수행하였다.
∑
×
min≤
≤
max∑
: 식생단위(군락/군집) 내의 종의 피도 적산값
: 전체 조사구 수: 종이 출현한 조사구수
max
×
: 대상 식생단위에 대한 종의 기여도
: 대상 식생단위 내의 기여도 최대값식물종명은 「국가표준식물목록」 (KBIS, 2016)과 Lee(1996) 를 따랐다. 멸종위기 야생식물과 식물구계학적 특정식물종, 생 태계 교란 야생식물을 특기하였다(Ministry of Environment, 2016). 귀화식물은 Park(2009)을 기준으로 구분하였다. 식생 공간분포 현황과 특성을 밝히기 위하여 국토정보플랫폼 (NGII, 2016)에서 제공하는 항공사진을 참조하여 QGIS 2.14.3으로 현존식생도를 제작, 분석하였다.
연구지역의 수질환경을 확인하기 위해 수온, 용존산소량, 염도, 전기염도도, pH 등을 1차 측정하였다. 당사해안습지 는 수리수문환경을 고려할 때, 정수환경, 유수환경, 우수환 경, 해수환경 등의 4가지 유형이 구분되며, 23개 지점에 대 해 DM측정기(YSI QUATRO-10102030)를 이용하여 수 질환경을 조사하였다.
3. 연구결과
3.1 군락분류
현장 식생조사 결과 당사해안 습지식생에 대해 28분류군 으로 이루어진 총 16개의 식생조사표를 획득하였으며, 종 조성과 서식처 환경에 의해 갯봄맞이-지채군락의 전형군락 과 올방개아재비하위군락, 물골풀하위군락, 갯꾸러미풀
s.l.
군락, 기장대풀군락, 갈대-꼬마부들군락 등의 4개 단위식 생으로 분류하였다(Table 1).
갯봄맞이-지채군락(
Glaux. maritima
var.obtusifolia-
Triglochin maritimum
community)은 갯봄맞이, 지채를 진 단종으로 구분하였다. 본 군락은 단층의 식생구조를 형성하 며, 최고 식생고는 15 cm 내외의 수변단경초본식물군락이 다. 군락 출현 종수는 17종이며, 구성종 가운데 갯봄맞이가 가장 높은 상대기여도를 나타낸다. 당사해안습지의 수변부 저수위대에 대상분포(zonal distribution)를 나타낸다. 전형 하위군락(typicum)은 수변부의 토심이 얕은 토양층에 분포 하고 있으며, 연중 얕은 수심이 유지되는 과습한 입지로 산 발적인 범람을 경험하는 서식처이다. 군락 구성종 가운데 지채, 천일사초, 갯까치수염, 갯길경, 갯사상자 등 염생식물 의 다양성이 높게 확인되는 군락이다(Table 1). 종조성 및 서식처 특성에서 일본 홋카이도 일대에서 확인된 지채군락 (Triglochin maritimum
community)과 유사하며, 일본의 최남단 입지에서는 지채의 기여도가 급격히 감소하는 경향 이 확인된다(Miyawaki and Ohba, 1965; Takagi and Shirai 2013). 올방개아재비하위군락(Eleocharis kamtschatica
subcommunity)의 구분종은 올방개아재비이며, 최고 식생 고는 20 cm 내외이다. 군락 출현 종수는 10종으로 전형군 락에 비해 상대적으로 적으며, 군락 내에 올방개아재비가 가장 높은 상대 기여도로 출현하였다. 전형하위군락에 비해 상대적으로 토심이 발달된 수변부이며, 군락 내 상대적으로 건조한 일부 입지에서는 갈대 유묘가 낮은 피도로 산발적 인 분포를 보였다. 일본의 경우 올방개아재비는 절멸위기종 I급종으로 고려되며, 최남단 분포지에서도 분포가 확인되기는 하나 낮은 빈도로 출현하고 있으며, 홋카이도 일대에서는Puccinellio kurilensis-Caricetum subspathaceae의 주요구 성종으로 아군집의 진단종으로 기재하고 있다(Miyawaki and Ohba, 1965). 종조성 특성을 고려할 때 갯개미취군강 (Asteretea tripolium)에 귀속될 것으로 판단된다. 물골풀하 위군락(
Juncus gracililimus
subcommunity)은 물골풀과 갯 하늘지기를 진단종으로 구분되었으며, 전형하위군락에 비 해 토양 내 자갈의 분포가 우세하게 확인되었다. 군락출현 종수는 11종이었으며, 고경초본식물군락과 인접한 입지에 서도 분포가 확인된다. 종조성 특성에 의해 갯개미취군강 (Asteretea tripolium)의 Puccinellion kurilensis에 귀속될 것으로 판단된다(Miyawaki and Ohba, 1965).갯꾸러미풀
s.l
.군락(Puccinellia nipponica s.l.
community) 의 진단종은 갯꾸러미풀s.l.
이다. 단층 식생구조를 형성하며, 최고 식생고는 30 cm 내외인 것으로 나타났다. 갯꾸러미풀s.l.
이 가장 높은 상대기여도로 출현하였다(Table 1). 해안 암반에 접한 서식처이며, 토심의 발달이 빈약한 입지이다. 상대적으로 수위변화에 따른 침수 영향을 적게 받는 입지로 갈대의 출현이 배제된 단층식물군락이며, 낮은 빈도지만 갯봄맞이가 혼생하고 있다. 갯꾸러미풀군락은 갯봄맞이를 진단종 가운데 하나로 하여 일본의 비교적 고위도해안지역에서 염습지수변 단경초본식물 군락인 Puccinellietum kurilensis의 갯봄맞이아군집 (Puccinellietum Glaucetosum maritimae)인 형태로 보고된 바 있다(Miyawaki and Ohba, 1965). 한편, 본 군락의 진단종인 갯꾸러미풀
s.l.
은 갯꾸러미풀속(Puccinella
)의 종으로 판단되 지만, 본종과 엽설 및 과피의 형질에서 일부 차이가 존재하며, 국내 종에서 형질이 일치하는 종이 존재하지 않아 종의 동정확 정을 보류하고 미결정 광의(s.l.: sensu lato
) 단위식생명을 제시 하였다. 선행연구에서 포항지역에 특이적 분포를 나타낸 갯겨 이삭(P. coreensis
)을 동일속 내 종으로 제시하고 있으나, 여전 히 형태적 차이가 존재하여 추후, 식물분류학적 재확인을 통해 단위식생명의 개선이 요구된다. 갯봄맞이-지채군락과 마찬가 지로 본 단위식생도 군집으로 명명하지는 않았으나, 종조성적 특징을 고려할 때 갯개미취군강(Asteretea tripolium)에 귀속 될 것으로 판단된다(Miyawaki et al., 1994).기장대풀-개피군락(
Beckmannia syzigachne-Isachne globosa
community)의 진단종은 기장대풀, 개피이다. 최고 식생고 가 1 m 내외로 2층의 식생구조를 나타낸다. 군락 출현 종 수는 5종이었으며 기장대풀이 가장 높은 상대기여도로 초 본1층을 우점하고 있다. 습지의 정수환경에서 바다로 이어 지는 유출구 수로에 소규모 군락을 형성하고 있으며, 하천 수변식생의 진단종 가운데 하나인 개피가 아우점종으로 출 현한다(Lee and Kim, 2005). 기장대풀과 개피가 혼생하는 본 군락은 국내에서 보고된 바 없는 독특한 군락이다. 일반 적으로 기장대풀군락은 빈영양 또는 중영양수질환경의 정 수서식처인 산지습지에서 분포가 보고되고 있다(Choi et al., 2012; Ryu, 2004; Miyawaki et al., 1994). 한국 저지 대의 습지는 일반적으로 집수역으로부터 반복적인 유기물 유입에 의해 부영양 환경을 형성하며, 지하 및 지상부로의 유출환경이 우세하게 형성된다. 따라서 저지대 습지에서 기장대풀우점군락이 출현하는 것은 드문 상황이다. 연구지역 은 습지대 내 용출소를 수원으로 하며, 좁은 집수역을 형성 하고 있어 외부로부터의 유기물 유입이 제한되기 때문에 기장대풀이 출현하고 있는 것으로 판단되며, 약한 수류환경 을 따라 주변 경작지 및 습지대에서 유입된 개피가 혼생하 는 양상을 보이는 것으로 판단된다. 종조성 및 식생특성을 고려할 때 산지습지에 분포하는 저층습원군락으로 보고된 기장대풀군락(
Isachne globosa
community)과 가장 유사하 여, 갈대군강(Phragmitetea communis)에 귀속시키는 것이 적절할 것으로 판단된다(Ryu, 2004; Choi et al., 2012).갈대-꼬마부들군락(
Thypa laxmannii-Phragmithes communis
community)의 진단종은 갈대, 털부처꽃, 꼬마부들 등이다.식생고는 2 m 내외로서 연구지역 습지식생 중 최고식생고를 나타내며, 2층의 층구조를 형성한다. 초본1층은 갈대가 우점하 며, 꼬마부들이 낮은 피도로 혼생한다. 초본2층은 천일사초가 가장 높은 상대기여도(40.7)를 보이며, 그 외 황새냉이, 소리쟁 이, 미국가막사리 등이 습윤하고 부영양화된 토양환경에 대응 하여 낮은 피도로 혼생하고 있다(Table 1). 습지 내 토심이 가장 깊은 입지를 중심으로 분포가 확인되며, 주변부와 습지 내 퇴적물의 발달을 견인하는 것으로 판단된다. 또한 지하경을 통한 영양번식에 의해 주변 단경초본식생 영역으로 지속적으 로 면적을 넓혀가고 있는 것이 확인되었다. 갈대-천일사초군 집과 주요 종에서 중첩을 확인하였으나, 연구지역의 토양환경 이 염습지환경과 서식처 차이를 보이며, 군락 내 다양한 구성종 출현양상에서 구별되었다(Lee and Kim, 2005). 국내 염습지 를 중심으로 보고된 갈대-천일사초군집과 유사한 식생형으로 고려되며, 갈대군강(Phragmitetea communis)이 상급단위로 판단된다(Miyawaki et al., 1994).
국내 갯봄맞이군락 및 분포서식지의 식생에 대한 현재까 지의 연구는 서식처에 대응된 진단종군 발굴에 관한 연구 가 아닌 분포집단 간 출현종 연구로 제한되었다(Kim et al.
2016). 그로인해 본 연구에서 추출된 단위식생과의 식물사 회학적 비교는 수행되지 못하였다. 추후 분포중심지에 보다 인접한 북부지역 식생형과의 종조성 및 분포결정인자 발굴, 군락동태, 군락구조, 군락생태 등에 대한 추가적 비교가 요 구된다.
3.2 갯봄맞이의 서식환경 분석
당사해안습지의 식물군락은 종조성 특성과 환경요인에 대한 좌표결정 분석으로부터 수분환경, 토심, 수위교란정 도, 식생고, 군락구조 등이 식생형성에 높은 연관관계를 가 지는 것으로 확인되었다(Fig. 2). 제1축은 수분환경 및 토심 으로부터 군락의 출현 및 종조성을 결정짓는 것으로 나타 났으며, 갯봄맞이-지채군락, 기장대풀-갈풀군락과 갯꾸러 미풀
s.l.
군락, 갈대-꼬마부들군락을 대별하는 것으로 고려 된다. 제2축은 기장대풀-갈풀군락, 갈대-꼬마부들군락과 갯꾸러미풀s.l.
군락을 대별하고 있으나, 갯봄맞이-지채군락 내에서는 전형군락과 올방개아재비하위군락, 물골풀하위군 락 간에 미지형을 바탕으로 한 차이가 군락 간 발생되고 있는것으로 확인되었다. 갯봄맞이가 우점 또는 아우점하는 단위 식생인 갯봄맞이-지채군락은 토심이 얕지만 수분환경이 습 윤하고, 수위변화가 잦으며, 식생고가 낮은 단층의 식생구 조에서 출현하는 것으로 분석되었다. 갯봄맞이가 낮은 기여 도로 출현하는 갯꾸러미풀
s.l.
군락은 갯봄맞이-지채군락에 비해 상대적으로 토심이 발달하며, 건조한 입지에서 분포를 나타내는 것으로 확인되었다. 1 m 이상의 식생고와 다층의 식생구조를 나타내는 기장대풀-개피군락과 갈대-꼬마부들 군락은 상대적으로 수위변화가 적은 안정적인 입지에서 분 포를 나타내는 것으로 확인되었으나, 갈대-꼬마부들군락이 보다 토심이 깊고 건조한 입지에서 서식하는 것으로 확인 되었다.집괴 분석 결과, 갯봄맞이가 출현하는 군락과 비출현군락 간의 명확한 집단차이가 확인되었으며, 공표형 상관계수 (cophenetic correlation)는 0.939로 높은 수치를 보였다.
11.01의 이질도(dissimilarity)에서 단위식생의 종조성에 따 라 식생형이 구분되며, 좌표분석 결과의 axis 2와 동일한 환경 경향에 의해 구분되었다. 14.05 이질도에서 axis 1의 환경요소에 따른 식생형의 구분이 확인되었으며, 갯봄맞이
-지채군락 내에서도 하위군락 간 종조성의 차이에 따른 집 괴형성이 명확히 구분됨을 확인하였다(Fig. 2). 집괴분석 결 과는 좌표결정 분석과 전반적으로 유사한 결과를 보이며, 각 단위식생별 고유의 서식처와 그에 대응하는 특유의 종 조성이 발달해 있는 것으로 나타났다.
일본의 갯봄맞이 최남단 단위식생과 국내 단위식생의 좌표 결정 및 집괴분석 결과 두 집단은 유사한 종조성 및 서식처 특성을 보였으며, 분포극지로서의 군락 종조성 변화에 대한 일반적 양상을 보여주고 있는 것으로 판단된다(Fig. 3). 국내와 일본 모두 갯봄맞이 분포중심지에 비해 상대적으로 종조성의 이탈(
Carex subspathacea, C. lyngbyei, C. ramenskii
andC. pseudocuraica
)과 제한적인 구성종의 출현이 나타내고 있 으며, 복합적인 종조성 및 우점정도의 변화가 발생되고 있는 것으로 판단된다(Miyawaki and Ohba, 1965). 현재 일본도 갯봄맞이를 멸종위기종으로 지정하여 보호하고 있으며, 최남 단 분포지에 대해서는 보다 적극적인 보호와 교육에 많은 노력 을 기울이고 있다(Takagi and Shirai, 2013). 두 지역 간 종 관리를 위한 보전방안 및 보호전략의 수립을 위한 상호협력체 계 구축이 요구된다.Fig. 2. Ordination and clustering analysis of wetland vegetation in Dangsa coastal wetland (relevé numbers (1~16) show such as Table 1).
Fig. 3. Comparison of Glaux maritima
community of southern distribution limit between Korea (no. 1~16) and Japan (no. 17~22).
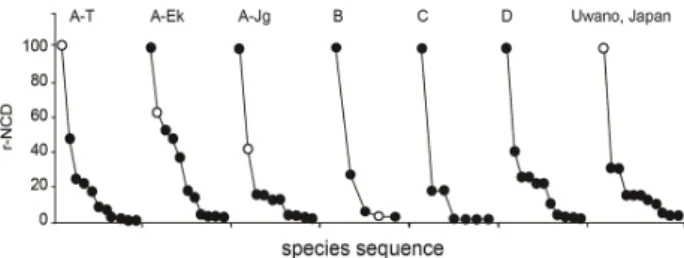
Fig. 4. Ranking according to r-NCD values of species
composition (acronyms are same such as Table 1) Open circles
are for
Glaux maritimavar. obtusifolia
연구지역 습지식생에 대한 상대기여도 분석으로부터 단 위식생 내 갯봄맞이 출현군락은 갯봄맞이-지채군락의 3개 하위군락과 갯꾸러미풀
s.l.
군락으로 확인되었다(Fig. 4). 갯 봄맞이 출현군락 가운데 전형하위군락에서 갯봄맞이는 가 장 높은 상대기여도(100)를 나타냈으며, 이는 일본의 최남 단 분포지역과 유사한 경향으로 확인되었다(Takagi and Shirai, 2013). 그러나 일본은 두 번째로 높은 상대기여도를 보이는 종이 상대적으로 낮은 상대기여도(30.8)를 보여 한 국과 차이를 보였으며, 최우점종인 갯봄맞이가 보다 우세하 게 군락을 형성하는 것으로 확인되었다(Fig. 4). 그 외 올방 개아재비하위군락과 물골풀하위군락, 갯꾸러미풀s.l.
군락에 서는 갯봄맞이 상대기여도가 63.2, 40.5, 5.9 등으로 상대 적으로 낮은 수치를 보였다. 이러한 수치의 경향은 연구지 역 식생의 토심깊이와 식생고의 증가경향과 유사하며, 식생고 1m 이상의 2층 구조 식생에서는 갯봄맞이의 출현이 제 한됨을 확인할 수 있었다(Fig. 4). 결국 갯봄맞이의 서식특 성이 높은 광환경을 선호하며, 주기적 교란이 발생되는 불 안정한 토양환경에서 선구적 식생을 형성한다는 선행연구 결과들(Brotherson, et al., 1984; Kim et al., 2016)과 일치 하는 결과를 확인하였다. 현재 연구지역 수변부의 얕은 토 심과 개방된 광조건 입지에서 갯봄맞이-지채군락이 여전히 우세한 식생을 형성하고 있으나, 토심이 발달할수록 올방개 아재비, 물골풀, 갯하늘지기, 갈대 등이 동일 입지 내에서 경쟁관계를 형성하는 것으로 고려된다. 반면 개미자리, 갯 까치수염, 갯길경, 갯사상자 등은 낮은 상대기여도이기는 하나 연구지역 습지식생에서 갯봄맞이-지채전형군락에서 만 분포를 보여, 연구지역 식생천이에서 갯봄맞이와 유사한 천이계열을 구성하는 수반종으로 판단된다.
3.3 현존식생도
연구지역의 군락단위 단위식생을 포함한 정밀 현존식생 도를 작성하였다(Fig. 5). 식생도는 11개 범례로 구성되며, 갯봄맞이-지채군락의 전형군락과 올방개아재비하위군락, 물골풀하위군락, 갯꾸러미풀
s.l.
군락, 기장대풀-개피군락, 갈대-꼬마부들군락 등의 습지식생과 이차초지식생, 암극식 생, 해안자갈식생 및 개방수면 등이었다(Fig. 5).A 습지는 습지 중앙부에 위치한 용출소를 중심으로 습지
Fig. 5. Actual vegetation map of Dangsa coastal wetland (Aug. 2016).
가장자리와 북측으로 흐르는 유출수입지를 따라 대상분포 경향을 나타내고 있다. 해안측으로 암극식생이 위치해 있으 며, 내륙측에 이차초지식생과 해안자갈식생 등이 위치해 있 다. B 습지는 습지 외부에서 발원한 지표수 영향으로 식생 이 분포하고 있으며, 내륙측에서 해안측으로 대상분포 경향 을 보였다(Fig. 5).
당사해안습지 식생 가운데 가장 넓은 면적(291.23 m2)은 갈대-꼬마부들군락이며, 전체 면적의 약 11.6%로 출현하 였다. 멸종위기종인 갯봄맞이가 출현하는 군락의 총 면적은 173.07 ㎡로 전체의 7.1%이며, 습지식생의 개방수면 인접 부와 자갈해안에 인접한 단경초본식물군락들에서 분포를 확인하였다(Fig. 5). 갯봄맞이가 우점종으로 출현하는 갯봄 맞이-지채군락의 면적은 26.6 m2이며, 전체 면적의 1.1%
에 불과한 것으로 확인되었다. 갯봄맞이가 습지 가장자리의 주기적 간섭이 발생하는 입지에 선구적으로 식생을 유지・
지속하고 있는 것으로 확인되었으나, 서식처 안정화와 함께 지속적으로 세력이 약해지는 상황으로 판단된다. 특히 식생 의 가장자리는 상대적으로 식생고가 높은 식생들이 위치하 고 있으며, 이들 군락의 근계 발달은 갯봄맞이의 주요 번식 전략 중 하나인 단위생식 번식을 저해하고 토양 퇴적을 견 인하여 갯봄맞이군락의 유지에 또 다른 위협요인인 것으로 판단된다(Grime, 1973; Jerling, 1988a; Jerling, 1988b).
그 외 습지 주변부에는 습지와 수문환경이 독립적인 건생 초지식생, 해안자갈식생, 해안암반식생 등이 위치해 있으 며, 해안암반 일대에는 식생이 형성되지 않은 물웅덩이가 산재해있다(Fig. 5). 이들은 현재 갯봄맞이 분포에 특별한 교란요인이나 기여요인으로 작용하고 있지 않으나, 인위적 교란 발생지를 중심으로 잠재적 종급원이 되고 있으며, 습 지유지를 위한 관리대상이 된다.
습지 동측에는 건생초지식생이 위치해 있으며, 억새, 참 새귀리, 큰김의털 등이 주요 우점종으로 출현한다. 일부 잔 자갈이 노출된 입지 및 애추사면에서는 포복형 소관목식물 인 돌가시나무와 인동덩굴이 높은 피도를 형성하였다. 해안 암반식생 범례가 넓은 면적을 차지하고 있으나, 이들 지역 은 피복된 식생의 면적이 매우 낮아 대부분이 무식생대를 이루며, 좁은 면적의 암상 또는 암극에 땅채송화, 큰개미자 리, 갯까치수염, 갯질경, 갯강아지풀 등의 소형식물들이 낮 은 피도로 출현하고 있었다. 해안자갈식생의 경우에도 입지 내 식생의 피도는 높지 않았으며, 갯메꽃, 수송나물, 번행초 등이 주요종으로 출현하였다.
3.4 당사해안습지의 식생지리적 의미
당사해안습지는 작은 규모의 습지임에도 불구하고 분포 식생 및 식물상을 고려할 때 식생지리 및 식물지리학적으 로 큰 가치를 지니는 습지로 판단된다. 멸종위기식물인 갯 봄맞이의 동아시아 최남단분포지로 판단되며, 입지 내에서 여전히 우점 또는 아우점 식생을 형성하여 양호한 생육상 태를 나타내고 있다(Fig. 5). 가까운 일본에 비해 위도 1°25′, 분포중심을 나타내는 유럽에 비해서도 1°00′가량
남측에 위치해 있는 것으로 확인되었다(Takagi and Shirai, 2013). 또한 연구지역은 인접한 국내 자생분포지와도 직선 거리 약 52 km 이격되어 분포하며, 주변지역에서 잠재분 포가능 대체서식지가 확인되지 않고 있다(Kim et al., 2016; Han and Park, 2014). 최남단 군락지의 특성과 대 체서식처의 이격분포 상황을 고려할 때 빙하기 유존식물인 갯봄맞이는 당사해안습지를 피난처(refugium)로 삼아 극히 의존되어 생육하고 있는 것으로 판단된다(Pickett and Thompson, 1978). 멸종위기종 및 종자원의 측면에서도 분 포극지는 종의 유전자 집단, 특히 분포제한환경에 대한 유 전적 적응과 발달을 반영한 메타개체군으로서의 의미 있는 연구재료로 고려되며, 추후 국내 갯봄맞이 집단의 유전자 다양성 확보를 위한 필수적 개체군으로 판단된다(NIBR, 2013; Hunter and Gibbs, 2011). 식물지리 및 식생지리학 적 특성에도 당사해안습지는 검바늘골, 물골풀, 갯꾸러미풀
s.l.
등의 북부요소 식물종과 지채, 갯하늘지기, 갯까치수염, 벋음씀바귀 등 남부요소 식물종이 기후대의 전이대적 식생 을 구성하고 있다(Lee, 1996; Miyawaki et al., 1994; Kim et al., 2012). 일본의 경우 현재 국내에서는 현장정보의 부 족으로 법정보호를 받지 않는 올방개아재비와 갯하늘지기 까지 절멸위기 I급과 II급으로 지정하여 갯봄맞이가 출현하 는 습지의 특성을 이해하고 보호하기 위한 노력을 하고 있 다. 국내에서도 당사해안습지의 생태학적 가치를 고려한 지 속적인 서식처 모니터링과 보호를 위한 방안의 마련이 필 요한 시기라고 판단된다.3.5 수환경 특성 및 관리를 위한 제언
당사해안습지의 수환경 특성을 확인하기 위해 습지 내 정 수입지, 유수입지, 우수입지, 해수입지로 구분하여 총 23개 입지에 대해 수온, 전기전도도, 염도, pH, 용존산소량 등을 측정되었다(Fig. 6). 본 연구에서는 측정이 1회만 이루어져 습지의 수환경을 온전히 반영하기는 어려우나, 추후 연구의 참고와 습지식생 유지에 대한 의미 있는 단초를 제공한다 고 판단된다.
입지별 평균수온은 우수수면이 25.7 ℃로 가장 높은 수온을 보였으며, 유수수면은 23.98 ℃, 정수수면 22.45 ℃, 해수수면 19,83 ℃으로 확인되었다. 우수입지는 암반 위에 형성된 서식 처로 지하수와 독립되어 있으며, 암반의 온도상승과 함께 수온 이 높게 나타난 것으로 판단된다. 유수입지와 정수입지는 일반 적인 경향과 달리 정수입지가 낮은 수온으로 확인되는데, 이는 연구지역 정수입지가 지하수 공급이 이루어지는 소형의 샘이 존재하기 때문으로 판단된다(KNA, 2012). 특히 갯봄맞이가 서식하는 습지대가 모두 이들 정수입지에 위치하고 있다는 점은 본 연구지역이 후빙기 이후 기온상승과 함께 동위도의 주변지역에서 분포가 사라진 것과 달리 연구지역에서만 갯봄 맞이의 잔존을 가능하게 했던 요인 중 하나로 판단된다. 전기전 도도와 염도는 정수입지가 0.73 ms/cm와 0.35 ppt로 상대적으 로 낮은 수치를 보여 담수습지의 특성을 나타냈으며, 유수입지 (5.32 ms/cm, 3.25 ppt), 우수입지(11.19 ms/cm, 7.00 ppt),
해수입지(47.95 ms/cm, 31.3 ppt)는 상대적으로 높은 수치를 보여, 해수의 영향을 직간접적으로 받고 있는 것으로 확인되었 다. 그러나 정수입지의 경우에도 일부 지점에서는 상대적으로 높은 수치(SW3 site: 0.93 ms/cm)를 보여, 수문환경 및 해안파 도의 영향정도에 따라 소형의 기수환경이 형성될 수 있을 것으 로 판단된다. 이러한 경향은 연구지역 주변의 유수수면과 우수 수면의 전기전도도와 염도가 해수와 인접한 정도, 정수습지로 부터 담수유입량, 수공간의 크기 등에 따라 다양한 수치차이를 보이는 것으로부터 추론된다(Fig. 6). pH에서도 우수입지 9.01, 유수입지 8.54, 해수입지 8.34에 비해 정수입지는 7.44로 상대적으로 낮은 수치를 나타냈으며, 기수 및 담수습지의 수질 환경과 유사한 pH범위를 보였다. 입지별 용존산소량(DO)은 유수입지가 11.19 mg/ℓ로 가장 높은 값을 보였으며, 우수입 지 8.09 mg/ℓ, 정수입지 7.15 mg/ℓ, 해수입지 6.23 mg/ℓ의 순으로 확인되었다(Fig. 6).
당사해안습지는 특히 해안에 인접해 있으면서 지하수 용출 이 일어나는 독특한 수리수문 특성에 의해 갯봄맞이의 대표적 인 서식처인 석호 환경과 유사한 미소환경(microhabitat)을 형성함으로써 분포가 가능했던 것으로 판단된다(Kim et al., 2016). 특히 인접한 해안의 해수유입과 해무에 의한 염분의 공급 및 교란은 선구자적 성격을 보이는 갯봄맞이에게 다른 종과의 경쟁을 극복하게 하는 환경요인으로 작용했을 것으로 사료된다(Jerling, 1998b; Grime, 2002).
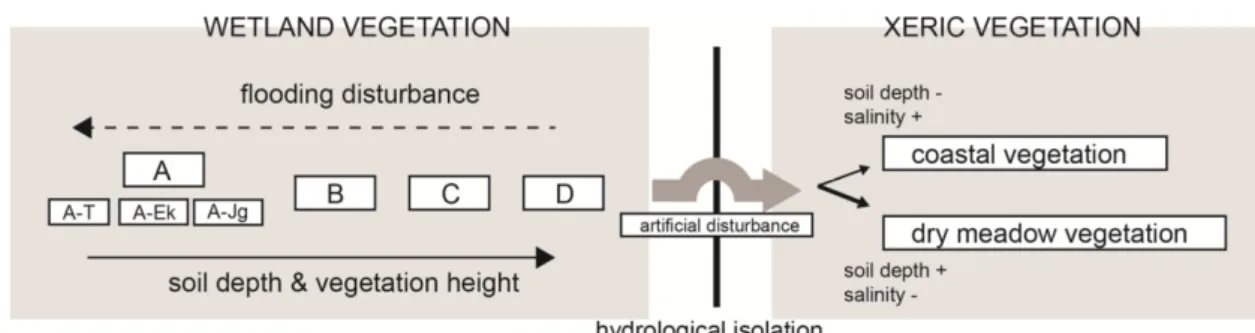
연구지역 식생의 특성을 고려할 때 현재 갯봄맞이 분포의 부정적 영향은 역설적이게도 서식처 안정화로부터 발생되 고 있는 것으로 판단된다(Figure 7). 습지의 많은 면적이 반복적 퇴적에 의한 토심 발달이 확인되며, 그와 동시에 갯 봄맞이군락의 분포가 다른 식생에 의해 교체되는 현상이 확인된다. 현재 수변부에 갯봄맞이에 의해 우점된 식생이 산발적으로 분포하고 있기는 하나, 갈대, 기장대풀, 개피 등 을 비롯한 상대적으로 높은 식생고를 형성하며, 경쟁우위에 있는 종들이 갯봄맞이 출현군락으로 분포를 확장해 갈 것 으로 판단된다. 따라서 갯봄맞이군락의 유지를 위해 이들 고경초본식생에 대한 생태학적 관리에 대한 신중한 접근이 요구된다(Kim et al., 2016). 또한 습지지역 일부 구간에서 소로길이 확인되며, 답압과 함께 수문교란이 확인되었다.
당사해안습지는 집수환경이 주변지역과 독립되어 있어 고 유한 수문체계를 구성하고 있으나, 인위적 교란에 의해 질 경이, 가시상추, 한삼덩굴, 좀명아주, 참새귀리 등의 외지종 유입이 확인되어 관리가 요구된다. 특히 이러한 교란은 입 지의 수문환경의 변화로부터 습지의 질적 변화를 가져올 수 있으며, 일부입지에서는 건생식생으로의 종 변화가 확인 되었다(Fig. 7).
연구지역 주변이 경작지로 활용되는 점도 잠재적 위협요 소로 판단된다. 이들 경작지의 토지용도 변경을 위한 습지 수문환경의 변화가 발생되거나, 경작지 식물의 유입 또는 Fig. 6. Water quality at Dangsa coastal wetland and around.
(SW: stable water sites, FW: flowering sites, OW: ombrophilic sites, SEA: sea water)
농약사용 등의 부정적 가능성이 당사해안습지의 유지에 위 협이 될 수 있을 것으로 판단된다. Kim et al.(2016), Son et al.(2011), Shin et al.(2012) 을 비롯한 일부 연구자에 의해 멸종위기종의 자생지 위치정보 노출이 서식지 파괴로 이어질 것을 우려한 바 있으나, 연구지역은 적절한 관리전 략만 갖춰진다면 종을 인식한 관찰자에 의한 위협은 상대 적으로 낮을 것으로 판단된다. 그러나 서식처 가치에 대한 미(未)인식으로 인한 돌발적 행위에 따른 위협요소가 보다 크다고 판단되므로 연구지역의 가치를 제시하고 지점에 대 한 지자체와 유관기관의 즉각적인 보전방안 마련을 요청한 다. 일본의 경우 이미 남한계 분포지역에 대해 수문환경을 비롯한 서식처 온전성을 유지하기 위한 시설물과 탐방로의 설치와 서식처가 가지는 가치에 대한 교육 및 안내가 이루 어지고 있으며, 현지 내(in situ) 보전을 위한 모니터링과 현지 외(ex situ) 보전을 위한 노력들이 병행되고 있다(Li and Pritchard, 2009; Mayberry and Elle, 2010). 국내에 서도 국제적 보호의 가치가 인정되는 입지에 대해서는 보 다 적극적이고 구체적인 보호계획의 수렵이 요구된다.
사 사
논문의 해석과 진행을 위해 도움을 주신 계명대학교 김종 원 교수님과 한강물환경연구소 정현기 박사님, 현장조사를 위해 노력해준 박지원, 신동주 학생에게 고마움을 전합니다.
References
Adam, P (1981). The Vegetation of British Saltmarshes,
The New Phytologist
, 88(1), pp. 143-196.Bae, SJ, Kang, BC and Han, JS (2014). Dormancy-break of Seeds by Low Temperature Treatment and Investigation of Factors affecting in Vitro Propagation in
Glaux maritima
L.Proceedings of the Conference on Korean Society For Horticultural Science
, Korea, May, 2014, p. 218. [Korean Literature]Baek, SH (2015).
Classification of Primulaceae species in Korea using DNA barcoding technique,
Master's Thesis, Daegu University. [Korean Literature]Bae, SJ, Kang, BC, Jeong, MY, Kim, SC, Kim, CK and Han, JS (2016). In vitro Multiplication through Single-Node Culture of Sea-Milkwort (
Glaux maritima L.
),Korea J. of Horticultural Science & Technology
, 6, pp. 461-471.[Korean Literature]
Becking, RW (1957). The Zürich-Montpellier school of phytosociology,
Botanical Review
, 23, pp. 411-488.Braun-Blanquet, J (1965).
Plant Sociology
, The study of Plant Communities. New York.Brotherson, JD and Barnes, SJ (1984). Habitat relationships of Glaux maritima in central Utah.
Great Basin Naturalist
, 44(2), pp. 298-309.Burnside, NG, Joyce, CB, Puurmann, E and Scott, DM (2007).
Use of vegetation classification and plant indicators to assess grazing abandoncent in Estonian coastal wetlands,
J. of Vegetation Science
, 18, pp. 645-654.Choi, BK (2013).
Syntaxonomy and Syngeography of Warm-Temperate Evergreen Broad-leaved Forests in Korea
. Doctor's Thesis, Keimyung University. [Korean Literature]Choi, BK, Kim, JW, Kim, SY and Lim, JC (2012). Vegetation of Jangdo island.
Korean Society of Environment and Ecology
. 26(4), pp. 512-527. [Korean Literature]Ellenberg, HH (2009).
Vegetation Ecology of Central Europe
, 4th Edition, Cambridge University Press, New York.Grime, JP (1973). Competitive exclusion in herbaceous vegetation.
Nature
242, pp. 344-347Grime, JP (2002).
Plant strategies, vegetation processes, and ecosystem properties
. John wily & sons, Ltd., New York.p. 471.
Hampe, A and Petit, RJ (2005). Conserving biodiversity under climate change: the rear edge matters.
Ecology letters
, 8(5), pp. 461-467.Han, GS and Park JH (2014). Water quality, flora and fauna of 7 wetlands in Donghae city.
J. of Wetland Research
, 16(4), pp. 335-352. [Korean Literature]Heidel, B, Jean, C and Crispin, S (2002).
Plant Species of Concern and Plant Associations of Powder River County
, Montana.Montana Natural Heritage Program. Montana. p. 137.
Fig. 7. Relation diagram of syntaxa with successional patterns and environmental factors in Dangsa coastal wetland
(acronyms are same such as Table 1).
Hitchcock, CL and Cronpuisy, A (1973).
Flora of the Pacific Northwest
, Univ. of Washington Press, seattle, p. 730.Hilbig, W (2003). The distribution of the vegetation in the Uvs‐
nuur basin and its surrounding mountain ranges.
Feddes Repertorium
, 114(7‐8), pp. 540-558Hu, CM and Kelso, S (1996). Primulaceae. In: Wu, Z.Y. &
Raven, P.H. (eds.), F
lora of China Vol. 15. Science Press
, Beijing & Missouri Botanical Garden Press, St. Louis, pp.99−185.
Hunter, Jr., ML and Gibbs, J (2011).
Fundamentals of conservation biology.
Blackwell Publishing, Oxford, pp.497.
Jerling, L (1988a). Clone Dynamics, Population Dynamics and Vegetation Pattern of Glaux maritima on a Baltic Sea Shore Meadow,
Vegetatio
, 74(2), pp. 171-185.Jerling, L (1988b). Population dynamics of
Glaux maritiama
(L.) along a distributional cline,Vegetatio
, 74(2), pp.161-170.
Kaplan, Z (2012). Flora and phytogeography of the Czech Republic,
Preslia
, 84, pp. 505-573.Kim, JW and Lee, YK (2007).
Classification and Assessment of Plant Communities
. Worldscience Press, Seoul. [Korean Literature]Kim, SM, Cho, YH, Kim, JH, Shim, SD and Kim, ST (2012).
Juncus fauriei
Lév. & Vaniot (Juncaceae): a new record in Korea.Korean J. of Plant Taxonomy
. 42(4): pp. 330-334.[Korean Literature]
Kim, YC, Chae, HH, Oh, HK and Lee, KS (2016). Distributional Characteristics and Factors Related to the Population Persistence, an Endangered Plant
Glaux maritima
var.obtusifolia
Fernald1a.Korean Society of Environment and Ecology
. 30(6), pp. 939-961. [Korean Literature]Korea National Aboretum(KNA) (2012).
Rare plants in Korea.
Korea National Aboretum, Gyeonggi-do.
Korea National Aboretum. p. 412 [Korean Literature]KMA (2016). http://kma.go.kr/.
Lee, WC (1996).
Coloured standard illustrations of Korean
, p.624, Academy, Seoul. [Korean Literature]
Lee, YK and Kim, JW (2005).
Riparian Vegetation of South Korea
. p. 293, Keimyung univ. press, Daegu. [Korean Literature]Li, DZ and Pritchard, HW (2009). The science and economics of ex situ plant conservation.
Trends in plant science
, 14(11), pp. 614-621.Markovskaya, E, Sonina, A, Sergienko, L, Morosova, K and Elkina, N (2014). Morphological and functional peculiarities of saltmarsh plants and epilithic lichens in tidal conditions of Russian Arctic Seas,
Czech Polar Reporys
, 4(2), pp. 168-177.Mayberry, RJ and Elle, E (2010). Conservation of a rare plant requires different methods in different habitats:
demographic lessons from Actaea elata.
Oecologia
164, pp.1121-1130.
Miehe, G, Miehe, S and Schlütz, F (2009). Early human impact in the forest ecotone of southern High Asia (Hindu Kush, Himalaya),
Quaternary Research
, 71(3), pp. 255-265.Minggagud, H and Yang, J (2013). Wetland plant species diversity in sandy land of a semi-arid inland region of China.
Plant Biosystems-An International J. Dealing with all Aspects of Plant Biology
, 147(1), pp. 25-32.Ministry of Environment (2016). http://me.go.kr/.
Miyawaki, A and Ohba, T (1965). Studien uber Strand-Salzwiesengesellschaften anf Ost-Hokkaido (Japan),
横浜国立大学理科紀要. 第二類, 生物学・地学
, 12, pp. 1-25. [German Literature]Miyawaki, A, Okuda, S and Fujiwara, K (1994).
Handbook of Japanese Vegetation
. 910pp, Shibundo, Tokyo. [Japanese Literature]Moon, KH (2005).
Syntaxonomy and synecology of the actual vegetation of Ulsan Metropolitan City
. Doctor's Thesis, Keimyung University. [Korean Literature]KBIS (2016). http://nature.go.kr/.
National Institute of Biological Resources(NIBR) (2012). Red data book of endangered vascular plants in Korea. National Institute of Biological Resources(NIBR). Incheon, 390pp.
[Korean Literature]
National Institute of Biological Resources(NIBR) (2013).
Genetic evaluation of important biological resources:
genetic diversity analysis for endangered species(2013)
. National Institute of Biological Resources, Incheon, 113pp.[Korean Literature]
NGII (2016). http://map.ngii.go.kr/.
O'Grady, JJ, Reed, DH, Brook, BW and Frankham, R (2004).
What are the best correlates of predicted extinction risk?.
Biological Conservation
, 118(4), pp. 513-520.eOH, SY (1980). A Review of the Family Primulaceae of Korea,
Kyungpook National University
, 29, pp. 379-417. [Korean Literature]Park, SH (2009).
New Illustrations and Photographs of Naturalized Plants of Korea
. Ilchokak, Seoul. [Korean Literature]Pickett, STA and Thompson JN (1978). Patch dynamics and the design of nature reserves.
Biological Conservation
, 13, pp. 27-36.Podani, J (2001). SYNTAX 2000:
Computer Program for Data Analysis in Ecology and Systematics
. Budapest: Scientia Publishing.Ryu, SH (2004).
Studies on vegetation and successive dynamics of moors in Motane zone, Korea
. 204 pp. Doctor's Thesis, Chungnam University. [Korean Literature]Sergienko, L, Markovskaya, E and Starodubtceva, A (2012).
Distribution of Vascular Plants in The Coastal Ecosystems
of The White Sea,
Water resources and wetlands
, Pertre, G, William, LJ, Petre, B (eds.), Editura Transversal, Tulcea, Romania, pp. 14-16.Shin, HT, Yi, MH, Shin, JS, Lee BC and Yoon, JW (2012).
Distribution of Rare Plants - Ulsan, Busan, Yangsan,
J. of Korean Nature
, 5(2), pp. 145-153. [Korean Literature]Son, SW, Lee, BC, Yang, HH and Seol, YJ (2011). Distribution of five rare plants in Korea.
Korean J. of Plant Taxonomy
, 41(3), pp. 280-286.Takagi, M and Shirai, N (2013). Growth environment of
Lysimachia maritima
community in the Uwano seashore,Noto Peninsula; vegetation research and soil salinity,
Bulletin of the Ishikawa Museum of Natural History
, 3, pp.1-9.
Westhoff, V and van der Maarel, E (1978). The Braun-Blanquet Approach.
In: Ordination and Classification of Communities
. R.H. Whittacker (ed.), Dr. W Junk by Publisher. Hague, Boston, London. pp. 287-399.Weber, HE, Moravec, J and Theurillat, JP (2000). International code of phytosociological nomenclature. 3rd edition.