─
─ 222 ──
하계 제주해협 주변 멸치 난∙자치어의 분포와 해양환경과의 관계
1.
멸치 난∙자치어의 분포현황이 승 종∙고 유 봉 제주대학교 해양학과
Distribution of Eggs and Larvae of the Anchovy,
Engraulis japonica, and Its Relationships to Environmental Factors Around Jeju Strait in Summer
1. Distribution of anchovy eggs and larvae
Seung Jong Lee and You Bong Go
Department of Oceanography, Cheju National University, Jeju 690-756, Korea
The distribution of eggs and larvae of the anchovy, Engraulis japonica was examined in the western part of the South Sea of Korea, including Jeju Strait, during July 16~21, 2001.
A comparative capture experiment of anchovy eggs and larvae was carried out using Bongo nets with two different mesh sizes (333µm and 505µm). The catches of larvae did not sig- nificantly differ between nets of either mesh size, but 99.2% of the total egg catch was obta- ined by the net with 333µm mesh.
High densities of eggs in early developmental stage occurred in the vicinity of Chongsan Is- land (Area J), the southwestern part of Jeju Strait adjacent to the northwest coast of Jeju Is- land (Area C), and the northwestern part of Jeju Strait in the vicinity of Chuja Island (Area A).
This suggests that these areas are major spawning grounds of the anchovy in the summer.
During the survey period, the proportion of late- developmental- stage eggs was higher in the eastern part of study area than in the western part.
Anchovy larvae were concentrated at 10 m and 20 m depths. Densities of larvae were markedly higher in the northern part of Jeju Strait (Area A, B) than the other sea areas.
The proportion of larvae whose total length is longer than 10.0 mm and 20.0 mm was higher in the eastern part (Area B, D) than in the western part of Jeju Strait (Area A, C). This suggests that anchovy larvae may be transported to the eastern part of Jeju Strait by a predominant northeastward flow pattern in summer.
Key words : Anchovy, Engraulis japonica, distribution, Jeju Strait, Bongo net
서 론
멸치(Engraulis japonica)는 전형적인 표층성 부어류 로서 국내에서는 주로 남해안에 주산란장을 갖고 있으 며, 연중 남해안에서 높은 비율로 어획되는 중요한 수산 종에 속한다(장 등, 1980; 국립수산진흥원, 1998). 이러한 수산업상의 중요성으로 인해 멸치에 대해서는 과거부터 많은 연구가 행해져 왔었다.
멸치는 서식해역내의 서로 다른 환경변화에 대응하면 서 재생산전략을 취하는 어종으로 알려져 있다(Bakun and Parrish, 1991; , 1992; ∙ , 1999). 즉 멸 치는 서식 해역내에 존재하는 다양한 해양환경 조건에 적응하면서 자신들의 재생산전략을 바꾸어 가며 종족을 유지하는 생태적 특성을 지닌 종이라고 말할 수 있다. 하지만 이러한 멸치의 생태적 특성에도 불구하고 지금 까지 국내에서 멸치의 산란 및 분포에 영향을 끼치는 해양환경요인을 논의하는데 있어서 수온, 염분으로 대표 되는 물리적 환경요인들과의 관계에 대해 주로 다뤄져 왔었고 그 이외의 환경변화와의 대응관계에 대한 논의 는 소수에 지나지 않았다(Kim, 1992; 김과 김, 1993; 추
와 김, 1998).
따라서 앞으로 멸치의 산란 및 난∙자치어의 분포를 논할 때에는 수온, 염분 이외에도 해역내에 존재하는 각 종 해양환경요인들을 동시에 조사하고, 이를 바탕으로 더 나아가 국내에서 멸치들이 어떠한 재생산전략을 취 하며 서식하고 있는지를 구체적으로 파악하는 것이 시 급히 수행되어야 할 과제라고 사료된다.
본 연구에서는 남해에서도 지금까지 멸치 난∙자치어 의 분포상황이 조사되지 않았었던 제주해협을 중심으로 이들의 분포양상을 파악하고, 이와 더불어 각종 해양환 경요인을 동시에 조사하여 이들 환경요인들이 멸치의 산란 및 난∙자치어의 분포에 어떠한 영향을 미치는지 에 대해 기초적인 정보를 알아보고자 하였다. 이번 논문 에서는 그 일차적 단계로 제주해협을 중심으로 한 멸치 난∙자치어의 분포현황을 살펴보는 것을 목적으로 하였 다.
재료 및 방법
조사는 2001년 7월 16일부터 21일까지 6일 동안 제
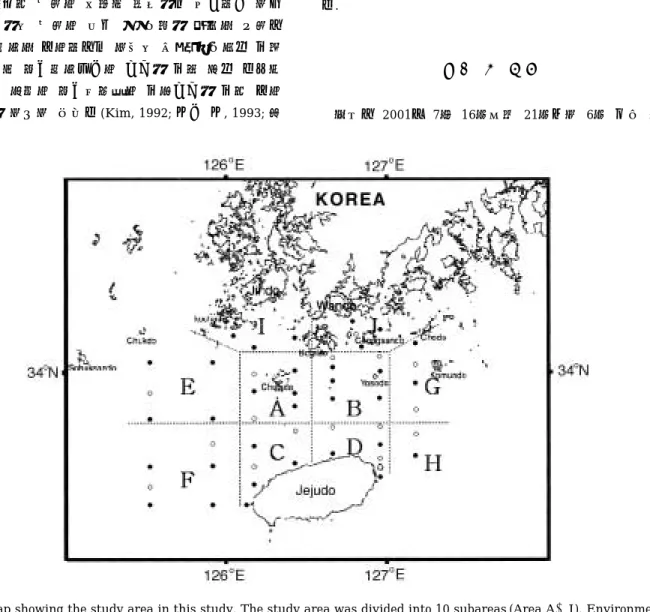
Fig. 1. Map showing the study area in this study. The study area was divided into 10 subareas (Area A~J). Environmen- tal observations were performed at all stations in July 16~21, 2001. Open circles (○) show biological sampling sta- tions.
주대학교 조사실습선 아라호를 이용하여 실시하였다. 이 번 연구에서는 조사해역을 총 10개의 해역으로 구분하 여 분석하였다(Fig. 1). 우선 제주해협(A~D해역)이라 함은 Fig. 1에서처럼 제주도를 기준으로 하여 동서방향 으로는 제주도 동쪽 우도 부근해역(127�00′E)부터 제주 도 서쪽 차귀도 부근(126�07′E)까지를, 북쪽으로는 보길 도 이남 해역(34�06′N)까지로 정의하였다. 제주해협은 다시 북서부역(A해역), 북동부역(B해역), 남서부역(C해 역) 그리고 남동부역(D해역) 등 각각 4개의 해역으로 구 분했다. 또 제주해협을 기준으로 서부 외양역을 북서부 외양역(E해역)과 남서부 외양역(F해역)의 2개 해역으 로, 또한 동부 외양역을 북동부 외양역(G해역)과 남동부 외양역(H해역)의 2개 해역으로 각각 구분하였다. 이밖 에 남해안에 해당되는 해역은 보길도를 중심으로 서쪽 을 거차군도 주변역(I해역), 동쪽은 청산도 주변역(J해 역)으로 각각 구분하였다. 전체 조사해역내에는 총 49개 의 정점을 설정하였고 모든 정점에 대해 CTD를 이용하 여 수온, 염분을 측정하였다. 멸치 난∙자치어 채집은 H 해역(1개 정점)을 제외하고 각각 구분된 해역내에 2~5 개의 정점을 선택하여 10 m, 20 m, 30 m 수층에 대해서 실시하였다.
이번 조사에서는 멸치 난∙자치어의 채집에 있어서 망목크기간 난∙자치어 출현량의 차이를 알아보고자 망 목 333µm과 505µm 두 개의 채집망이 부착된 봉고네 트(망구직경 60 cm)를 이용하였다. 채집은 각 수층을 2~3 knot의 속도로 약 10분간 수평예망하여 실시하였 다. 채집된 시료는 선상에서 5% 해수포르말린으로 고정 시킨 후 실험실에서 멸치 어란 및 자치어만을 분리, 계 수하였다. 또 멸치 난∙자치어의 분포밀도는 채집시에 네트입구에 부착시킨 유량계의 회전수에 의해 여과수량 을 계산한 후 해수 1,000 m3당 개체수로 환산하였다. 망 목의 크기에 따른 멸치 난∙자치어 출현량에 대한 유의 성 관계는 T-test (SPSS 10.0)를 사용하여 검증하였다. 또 각 수층에 대한 멸치 난∙자치어의 전체적인 분포양 상은 각 해역별 평균밀도로 표시하였다.
멸치 어란은 난내 발생진행 상황에 따라 산란된 후 얼마 되지 않아 배체가 형성되지 않은 어란을 I단계, 배 체가 형성되어 있으나 미부가 난황에서 떨어져 있지 않 은 어란을 II단계, 미부가 난황과 떨어져 있는 부화 직 전의 어란을 III단계로 하는등 총 3가지 단계로 구분하 였다. 멸치 자치어인 경우에는 채집된 모든 개체에 대해 0.1 mm 단위로 전장을 측정하였다.
본 연구에서 이용한 수온, 염분 CTD 자료는 제주대학 교 해양산업공학부 어장환경학실험실의 자료임을 밝힌 다.
결 과
1. 수온∙염분의 분포 변화
조사기간내 연구해역의 각 수층별 수온∙염분의 수평 분포 상황을 살펴보면(Fig. 2), 우선10 m 수층인 경우는 전체적으로 12.9~26.3�C의 수온범위를 보이고 있었다. 제주도 주변해역은 대마난류수의 영향으로 인해 22.6~ 26.3�C의 고수온의 해수가 존재하고 있었고 남해안으로 접근할수록 점차 수온의 낮아지는 경향을 보였다. 특히 제주해협 북서부 외양역(E해역)부터 제주해협 북부역 (A~B해역) 그리고 제주해협 북동부 외양역(G해역)에 걸쳐 강한 수온 전선대가 동서방향으로 길게 형성되어 있는 것이 특징적이었다. 염분은 28.8~32.7‰의 범위를 보이고 있었고, 제주해협 남서부 외양역(F해역)은 중국 대륙연안수의 영향으로 28.8‰ 정도의 저염분대 해수가 제주해협내로 유입되고 있었다. 또 청산도 주변역(J해 역)을 중심으로 남해안 연안을 따라 염분전선대가 형성 되고 있었으며 남해안 연안쪽이 상대적으로 고염의 특 성을 보이고 있었다.
20 m 수층인 경우에는 수온범위가 12.1~23.1�C로 10 m 수층보다 평균 3~4�C 정도 낮게 나타나고 있었다. 제 주도 주변에는 20�C 이상의 고온수가 분포하고 있었고 제주해협의 남서부 외양역(F해역)을 시작으로 북동방향 으로 수온전선대가 비스듬하게 형성되고 있었다. 염분은 31.4~33.4‰의 범위로 10 m 수층에 비해 평균 1‰정도 높게 나타났으며 10 m 수층에서와 같은 저염분대의 해 수는 출현하지 않았다. 그리고 제주해협 남서부역(C해 역)과 제주해협 동부 외양역(G, H해역)에서 비교적 고 염의 해수들이 분포하고 있었다.
30 m 수층은 수온범위가 10.2~20.7�C로 20 m 수층에 비해 평균 2~3�C 낮은 분포범위를 보이고 있었으며 등 온선의 분포는 20 m 수층과 비슷한 양상을 나타내고 있 었다. 염분은 32.1~34.0‰의 범위로 20 m 수층에 비해 평균 0.5‰ 정도가 더 높았고, 등온선의 분포와 유사한 양상으로 제주해협 남서부 외양역(F해역)에서부터 북동 방향으로 비스듬하게 염분전선대가 형성되고 있었다.
2. 망목에 따른 멸치 난∙자치어의 출현량 비교 이번 연구에서는 망목 333µm와 505µm의 두 가지 채집망이 장착된 봉고네트로 동시에 채집을 행한 후 각 네트에 채집된 멸치 난∙자치어의 출현량을 서로 비교 해 보았다(Table 1).
우선 멸치 어란인 경우에는 전 수층을 통틀어 망목
333µm네트에서는 평균밀도 2,125 No./1,000 m3, 망목 505µm네트에서는 평균밀도 17 No./1,000 m3가 출현하
였는데 이는333µm네트에서가 전체 출현량의99.2%를 차지하고 있는 것으로서505µm네트(0.8%)보다 월등히 Fig. 2. Horizontal distribution of temperature (�C) and salinity (‰ ) at the depths of 10 m, 20 m and 30 m.
Table 1. Comparisons of mean abundances of anchovy eggs and larvae collected by a Bongo net with two different mesh
sizes (333µm and 505µm) (Unit: No./1,000 m3and ind./1,000 m3)
Depth Mean eggs abundance Mean larvae abundance
(m) 333µm % 505µm % df t P 333µm % 505µm % df t P
10 m 1,102 99.6 4 0.4 6 4.192 * 49 50.5 48 49.5 21 1.098 n.s.
20 m 593 98.8 7 1.2 7 6.040 * 40 53.3 35 46.7 19 -1.151 n.s.
30 m 430 98.6 6 1.4 6 5.238 * 21 51.2 20 48.8 17 0.792 n.s.
Total 2,125 99.2 17 0.8 110 51.6 103 48.4
*P⁄0.05; n.s., not significant.
많은 어란 출현량을 나타내고 있었다. 또한 망목별 멸치 어란 출현량에 대한 T-test 결과에서도 모든 수층에서 출현량에 유의한 차이(유의수준 5%)를 보여주었다.
멸치 자치어의 출현량에 있어서는 전 수층을 통틀어 망목 333µm네트는 평균밀도 110 ind./1,000 m3(51.6%), 망목 505µm네트는 평균밀도 103 ind./1,000 m3 (48.4%) 가 출현하였으며 수층별로 살펴보면 333µm네트쪽이 조금 많았을 뿐 전체적인 출현량에는 큰 차이가 없었다. 또 망목별 자치어 출현량에 대한 T-test 결과에서도 전 수층에서 유의한 차이는 나타나지 않았다.
이상의 결과로부터, 멸치 어란의 채집에는 망목 333 µm네트가 더 유효하였고 멸치 자치어에 있어서는 망목 에 따른 출현량의 차이가 크지 않음을 알 수 있었다. 따 라서 이후부터 서술하는 멸치 난∙자치어에 관한 모든 사항은 망목 333µm네트에서 채집된 것들을 대상으로 하였다.
3. 멸치 난∙자치어의 분포현황
조사기간 동안 연구해역내에서 채집된 멸치 난∙자치 어의 해역별 평균 분포밀도(Table 2)와 수층별 분포상 황(Fig.3)은 다음과 같다.
1) 멸치 어란
이번 조사에서의 멸치 어란은 전 수층을 통틀어 평균 2,125 No./1,000 m3정도로 출현하였다. 10 m 수층에서는 전체 평균 1,102 No./1,000 m3정도의 멸치 어란이 출현 하고 있었는데 해역별로 살펴보면, 청산도 주변역(J해 역)에서 평균 9,351 No./1,000 m3로 가장 많았고 제주해 협 북동부 외양역(G해역)에서도 289 No./1,000 m3로 비 교적 많은 출현을 보였다. 제주해협인 경우에는 특히 서 부에 해당하는 북서부역(A해역)과 남서부역(C해역)에서 각각 평균 1,172 No./1,000 m3와 172 No./1,000 m3로 동 부(B, D해역 평균 10 No./1,000 m3에 비해 상대적으로
높았으며 반면에 제주해협 서부 외양역(E, F해역)과 제 주해협 남동부 외양역(H해역)에서는 평균 3 No./1,000 m3정도로 아주 낮았다.
20 m 수층에서는 전체 평균 593 No./1,000 m3정도가 출현하였는데 10 m 수층과 유사한 양상으로 청산도 주 변역(J해역)에서 평균 3,423 No./1,000 m3로 가장 많이 출현하고 있었다. 제주해협 북동부 외양역(G해역)에서 는 평균 1,294 No./1,000 m3로 10 m 수층보다 더 높은 값을 나타냈다. 제주해협인 경우도 10 m 수층과 마찬가 지로 북서부역(A해역)과 남서부역(C해역)이 각각 평균 591 No./1,000 m3와 573 No./1,000 m3로 북동부역(B해역) 과 남동부역(D해역)의 각각 22 No./1,000 m3, 9 No./1,000 m3에 비해 분포밀도가 높았다. 한편 외양역에 해당하는 제주해협 서부 외양역(E해역 20 No./1,000 m3, F해역 2 No./1,000 m3)에서는 낮은 값을 보이고 있었다.
30 m 수층인 경우는 전체 평균 430 No./1,000 m3로 채 집수층 중 출현량이 가장 낮았는데 그 분포양상을 살펴 보면 역시 청산도 주변역(J해역)이 평균 2,047 No./
1,000 m3로 가장 많았고 제주해협 북동부 외양역(G해 역)에서 평균1,258 No./1,000 m3로 그 다음으로 많이 출 현하고 있었다. 제주해협내에서는 남부에 해당하는 남서 부역(C해역)과 남동부역(D해역)이 각각 750 No./1,000 m3와 119 No./1,000 m3로 북부인 북서부역(A해역) 98 No./1,000 m3, 북동부역(B해역) 10 No./1,000 m3보다 상대 적으로 높은 값을 보이고 있었다.
이상의 결과를 종합해 보면, 연구해역내에서의 멸치 어란은 전 수층을 통틀어 청산도 주변역(J해역)과 거문 도 부근의 제주해협 북동부 외양역(G해역), 추자도 중심 의 제주해협 북서부역(A해역) 그리고 제주도 북서부연 안에 인접한 제주해협 남서부역(C해역)등 전반적으로 외양역보다는 일부 섬을 중심으로 한 집중분포 현상이 뚜렷하였다. 한편 보길도 서쪽에 해당하는 거차군도 주 변역(I해역)에서는 전 수층을 통해 멸치 어란들이 출현
Table 2. Mean abundances of anchovy eggs and larvae collected in the study area on July, 2001
(Unit : No./1,000 m3and ind./1,000 m3)
Depth Mean eggs and larvae abundance
Average
(m) A B C D E F G H I J
10 m egg 1,172 10 172 11 6 3 289 1 - 9,351 1,102
larvae 188 113 38 12 33 31 12 - 11 49 49
20 m egg 591 22 573 9 20 2 1,294 - - 3,423 593
larvae 72 103 43 23 32 41 24 - 13 44 40
30 m egg 98 10 750 119 20 2 1,258 - - 2,047 430
larvae 11 42 20 21 33 20 38 - - 22 21
Total egg 1,861 42 1,495 139 46 7 2,841 1 - 14,821
larvae 271 258 101 56 98 92 74 - 24 115
하지 않고 있는 것이 특징적이었다. 2) 멸치 자치어
멸치 자치어는 전 수층을 통틀어 평균 110 ind./1,000 m3정도로 출현하고 있었다. 수층별로 살펴보면, 우선10 m 수층에서는 전체 평균 49 ind./1,000 m3정도의 멸치
자치어가 출현하였으며 특히 제주해협 북부에 해당하는 북서부역(A해역)과 북동부역(B해역)에서 각각 평균188 ind./1,000 m3와 113 ind./1,000 m3로 출현량이 가장 많았 다. 반면에 그 외의 해역들에서는 모두 평균 50 ind./
1,000 m3이하의 낮은 값을 보였다.
20 m 수층에서는 전체 평균 40 ind./1,000 m3정도의 Fig. 3. Distribution of mean abundances of anchovy eggs (No./1,000 m3) and larvae (ind./1,000 m3) at the depths of 10 m, 20
m and 30 m.
출현을 보였다. 해역별 분포양상을 살펴보면 10 m 수층 보다는 분포밀도가 낮았지만 역시 제주해협 북부역인 A, B해역에서 각각 평균 72 ind./1,000 m3와 103 ind./
1,000 m3로 높은 값을 나타내고 있었다. 그 외 나머지 해역에서도 모두 평균50 ind./1,000 m3이하로 낮았다.
30 m 수층인 경우는 전체 평균 21 ind./1,000 m3로 채 집수층 중 멸치 자치어가 가장 적게 출현하고 있었다. 해역별로 살펴보았을 때에도 모든 해역에서의 평균분포 밀도가 10~50 ind./1,000 m3의 범위로 나타나 다른 수 층에 비해 아주 낮은 분포밀도를 보여주고 있었다.
따라서 조사기간 동안 연구해역내에서의 멸치 자치어 는 어란과는 달리 10 m와 20 m 수층에서 주로 출현하였 는데 대부분 제주해협 북부역(A, B해역)내에서 고밀도 로 분포하고 있었고, 수심이 깊어질수록 멸치 자치어의 분포밀도가 점차 낮아지는 경향을 나타내고 있음을 알 수 있었다.
4. 멸치 어란의 발생단계별 분포양상
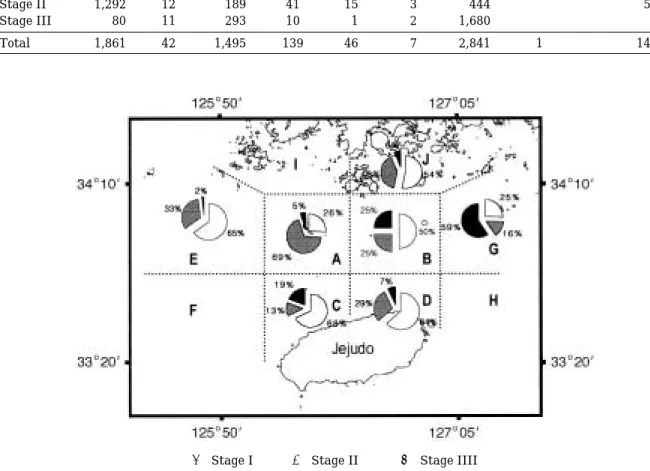
멸치 어란의 발생단계별 분포밀도와 분포비율상황을 해역별로 구분하여 살펴보았다(Table 3, Fig.4). 여기서 발생단계별 분포비율을 논의할 때에 전체 평균분포밀도 가 10 No./1,000 m3이하로 출현한 제주해협 남서부 외 양역(F해역)과 제주해협 남동부 외양역(H해역)은 논의 대상에서 제외시켰다. 우선 전 수층 평균분포밀도에서는 청산도 주변역(J해역), 제주해협 북서부역(A해역), 제주 해협 남서부역(C해역), 제주해협 북동부 외양역(G해역) 이 모두 1,400 No./1,000 m3이상으로 높은 값을 보이고 있었다. 발생단계별로 살펴보면, 산란된 후 얼마 지나지 않은 I단계 어란의 경우는 제주해협 남부역(C, D해역) 과 북동부역(B해역), 제주해협 북서부 외양역(E해역) 그 리고 청산도 주변역(J해역)에서 50% 이상의 높은 비율 로 출현하고 있었다. 평균분포밀도는 J해역이 7,927 No./1,000 m3로 가장 많았고 그 다음으로 C해역 1,013
Table 3. Mean abundances of anchovy eggs by the developmental stages (Unit: No./1,000 m3)
Developmental Mean egg abundance
stage A B C D E F G H I J
Stage I 489 19 1,013 88 30 2 717 1 - 7,927
Stage II 1,292 12 189 41 15 3 444 - - 5,953
Stage III 80 11 293 10 1 2 1,680 - - 941
Total 1,861 42 1,495 139 46 7 2,841 1 - 14,821
Fig. 4. Spatial distribution of anchovy eggs by developmental stages in the study area.
� Stage I � Stage II � Stage IIII
No./1,000 m3, G해역 717 No./1,000 m3, A해역 489 No./1,000 m3의 순으로 출현하고 있었다.
난내에 배체가 형성된 II단계 어란은 제주해협 북서 부역(A해역)이 69%로 분포비율이 가장 높았고 그 다음 으로 청산도 주변역(J해역) 40%, 제주해협 북서부 외양 역(E해역) 33%, 제주해협 남동부역(D해역) 29%의 순으 로 나타나고 있었다. 평균분포밀도는 J해역이 5,953 No./1,000 m3로 가장 높고 A해역 1,292 No./1,000 m3, G 해역 444 No./1,000 m3, C해역 189 No./1,000 m3순으로 출현하고 있었다.
부화하기 직전의 III단계 어란은 제주해협 북동부 외 양역(G해역)이 59%로 가장 높았으며 다음으로 제주해 협 북동부역(B해역) 25%, 제주해협 남서부역(C해역) 19% 순으로 나타나고 있었다. 평균분포밀도 역시G해역 이 1,680 No./1,000 m3로 가장 많았고 J해역 941 No./
1,000 m3, C해역 293 No./1,000 m3순으로 나타나고 있었 다.
이상의 결과들을 종합해 보면, 조사기간 동안 연구해 역내에서 I단계 멸치 어란들은 외양역(E, F, H해역)에 비해 주로 섬이나 연안역 부근(A, C, J해역)에서 대부분 높은 비율(50% 이상)을 차지하며 고밀도로 출현하고 있 었고, II단계 어란인 경우는 모든 해역에서 출현하였으 나 뚜렷한 경향은 나타나지 않았지만 부화하기 직전의 III단계 어란은 전반적으로 연구해역의 동쪽 방향으로 갈수록 그 출현율이 점차 증가하고 있는 경향임을 알
수 있었다.
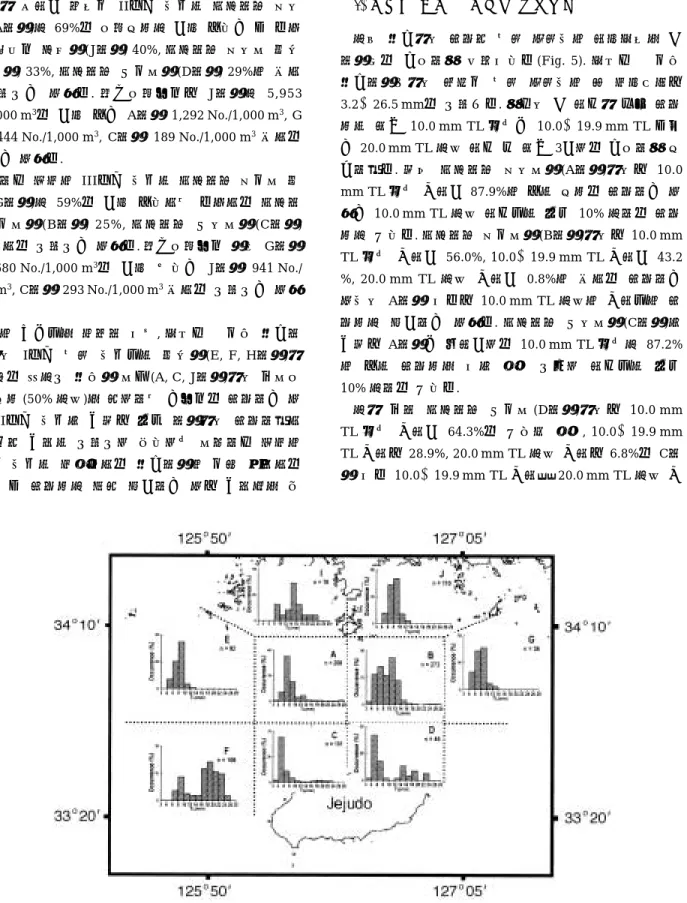
5. 해역별 멸치 자치어의 체장조성
이번 연구에서 출현한 멸치 자치어의 체장조성을 각 해역별로 구분하여 살펴보았다(Fig. 5). 조사기간 동안 연구해역내에서 채집된 멸치 자치어의 총 전장범위는 3.2~26.5 mm로 나타났다. 여기서 각 체급에 따른 출현 율은 크게 10.0 mm TL 미만과 10.0~19.9 mm TL 그리 고20.0 mm TL 이상 체급 등 크게 3가지로 구분하여 비 교하였다. 우선 제주해협 북서부역(A해역)에서는 10.0 mm TL 미만 개체가87.9%의 높은 비율로 출현하고 있 었고10.0 mm TL 이상 체급들은 모두 10% 이하로 출현 율이 낮았다. 제주해협 북동부역(B해역)에서는 10.0 mm TL 미만 개체가 56.0%, 10.0~19.9 mm TL 개체가 43.2
%, 20.0 mm TL 이상 개체가 0.8%의 순으로 출현하고 있어서 A해역 보다는 10.0 mm TL 이상의 개체들의 출 현율이 증가하고 있었다. 제주해협 남서부역(C해역)인 경우는 A해역과 마찬가지로 10.0 mm TL 미만이 87.2%
의 높은 출현율을 보인 반면 나머지 체급들은 모두 10% 이하로 낮았다.
이에 대해 제주해협 남동부(D해역)에서는 10.0 mm TL 미만 개체가 64.3%로 낮아진 반면, 10.0~19.9 mm TL 개체는 28.9%, 20.0 mm TL 이상 개체는 6.8%로 C해 역 보다10.0~19.9 mm TL 개체와 20.0 mm TL 이상 개
Fig. 5. Total length frequency distributions of anchovy larvae collected in the study area.
체의 출현율이 증가하고 있었다.
한편, 제주해협 북서부 외양역(E해역)과 북동부 외양 역(G) 그리고 청산도 주변역(J해역)에서는 10.0 mm TL 미만 개체들이 모두 90% 이상을 차지하였으며 상대적으 로 10.0 mm TL 이상 개체들의 출현율은 극히 낮았다. 이에 대해 제주해협 남서부 외양역(F해역)인 경우에는 10.0 mm TL 미만 개체가 18.5%인데 반해 10.0~19.9 mm TL 개체는 48.1%, 20.0 mm TL 이상 개체는 33.4%
로 대형개체들의 출현율이 높게 나타나고 있었다. 또 거 차군도 주변역(I해역)인 경우도 비록 개체수는 적었지 만 10.0 mm TL 이상 개체가 84.2%로 높은 출현율을 보 이고 있었다.
이상의 결과로부터, 제주해협인 경우 서부역에 해당하 는 A, C해역에서는 주로 10.0 mm TL 미만 개체들이 많 은(87% 이상) 반면에 동부역인B, D해역에서는10.0 mm TL 보다 큰 개체들의 출현율이 높았고 이 밖에 제주해 협 이외의 E, G, J해역인 경우는 모두 10.0 mm TL 미만 의 개체가 주(90% 이상)를 이루고 있었으며 F, I해역에 서는 10.0~19.9 mm TL과 20.0 mm TL 이상의 비교적 큰 개체들이 많이 출현하고 있었음을 알 수 있다.
고 찰
일반적으로 해양에서 중∙대형 동물플랑크톤을 채집 할 때에는 망목330µm의 네트가 그리고 중∙소형 플랑 크톤은 망목 100~200µm의 네트가 널리 이용되고 있 으며( ∙ , 1976), 어류의 난∙자치어 채집시에는 전형적으로 망목 333µm과 505µm의 네트를 이용한 채 집이 주를 이루고 있다( , 1981). 특히 난∙자치어의 경 우에는 어떠한 망목의 네트를 선택하느냐에 따라서 어 종별로 그 출현량이 달라지는 것으로 알려지고 있다 (Lenarz, 1972; Weinstein et al., 1980; 谷, 1987). 이에 대한 원인으로는 어란의 형태나 크기, 난막이 지니는 장 력 그리고 자치어의 체형이나 체고 또는 체장등을 들 수 있다. 기존 연구결과에 의하면 망목 333µm와 505 µm 두 개의 네트를 가지고 난∙자치어를 동시에 채집 했을 경우에는 333µm쪽에서가 출현량이 유의하게 많 다는 것이 보고되고 있는데(Barnett et al., 1984; 차, 2002), 이번 조사에서도 333µm과 505µm 두 가지 망목 의 봉고네트를 가지고 동시에 채집을 행한 결과 멸치 자치어의 출현량에 있어서는 전 수층을 통틀어 유의한 차이가 없는 반면 멸치 어란인 경우에는 333µm네트에 서가 전체 출현량의 99.2%를 차지하고 있어서 505µm 네트에서는 멸치 어란의 채집이 거의 이루지지 않고 있
음을 알 수 있었다. 이와 같은 양상은 천수만에서 금강 하구에 이르는 해역에서 실시된 연구에서 채집된 멸치 어란의 약 96.6%가 333µm네트에서 출현하였다는 차 (2002)의 보고와도 잘 일치하고 있음을 알 수 있다. 이 에 대한 가장 주요한 원인으로는 난의 크기를 들 수 있 는데 일반적으로 난경이0.7 mm 이하의 어란(멸치 어란 은 타원형으로 단경이 0.60~0.75 mm임)은 네트의 예망 시 수압에 의해 505µm 망목을 쉽게 빠져나간다고 추정 되고 있다(차, 2002). 이상의 내용들을 종합해 보았을 때 앞으로 멸치 난∙자치어의 채집을 행할 경우 망목크기 의 선택에 있어서는 505µm네트보다 333µm네트를 사 용하는 것이 바람직하다고 판단된다. 이와 더불어 금후 에 망목이333µm보다 더 작은 중∙소형 플랑크톤 채집 용 네트를 이용한 멸치 난∙자치어 채집도 실시해 봄으 로써 망목에 따른 출현량에 어떠한 변화가 있는지 또한 그 원인은 무엇인지에 대해서도 자세히 논의할 가치가 있다고 생각된다.
이번 조사기간 동안 망목 333µm네트에 의해 출현한 멸치 어란은 전체 평균 2,125 No./1,000 m3로 조사기간 인 7월을 기준으로 살펴보았을 때 서해안 일대(허와 유, 1984)에서의 118.7 No./1,000 m3, 남해 중부 고흥반도 부 근해역(한 등, 2002)에서의 1,798 No./1,000 m3와 비교하 여 비슷하거나 높은 수준으로 출현하고 있음을 알 수 있었다. 멸치 자치어인 경우에는 이번 조사에서 전체 평 균 110 ind./1,000 m3가 출현하고 있어서 서해안 일대(허 와 유, 1984)에서의 약 250 ind./1,000 m3, 고흥반도 부근 해역(한 등, 2002)에서의 7,232 ind./1,000 m3, 대한해 협(이 등, 1996)에서의 27,617 ind./1,000 m3와 비교하여 출현량이 훨씬 적었다. 이러한 출현량의 차이가 환경과 생물량 분포, 정점수 또는 채집방법의 차이에 의한 것인 지 등에 대해서는 금후 논의의 여지가 있다고 생각된다. 우리나라 남해안에서 멸치의 산란은 3~11월에 걸쳐 일어나며, 집중적으로 산란을 행하는 시기는 4~8월에 해당한다(최와 김, 1988; 김과 강, 1992). 따라서 이번 조 사기간인 7월도 연구해역내에서 멸치들이 활발히 산란 을 행하는 시기라 판단된다. 또한 멸치는 산란시기나 서 식해역내의 해양환경에 따라서도 산란장소가 달라지는 어종으로 잘 알려져 있다( ∙ , 1999; Aoki and Miyashita, 2000). 본 조사기간 동안 멸치 어란들의 고밀 도 분포는 청산도를 중심으로 하는 청산도 주변역(J해 역)과 제주해협내 추자도 주변해역(A해역), 제주도 북부 연안과 인접한 제주해협 남부역(C, D해역) 그리고 초도 와 거문도 주변역(G해역) 등 외양역 보다는 주로 연안 역에서 더 뚜렷하게 나타나고 있었다. 이와 같은 현상은 남해안 일대에서 멸치 어란들이 외양보다 연안측에서
대부분 높은 밀도로 분포하고 있다는 기존의 보고(김, 1983; 김과 김, 1991)와도 유사한 결과였다. 따라서 이번 연구결과를 통해 본 조사해역내에서의 멸치의 산란도 일부 남해안 연안역과 제주해협내 섬과 인접한 연안역 에서 주로 이루어지고 있다고 사료된다. 한편 이번 조사 에서는 특이하게 거차군도 주변역(I해역)에서 전 수층 을 통틀어 멸치 어란이 한 개도 발견되지 않았다. 일반 적으로 어류의 산란에는 여러 외부 환경요인들 중 수온 이 가장 중요한 요인으로 작용하고 있는데(Asahina and Hanyu, 1983; Razani and Hanyu, 1986) 멸치의 경우에 는 산란에 필요한 임계수온이 14~15�C 정도이며, 15�C 이하의 수온환경에서는 멸치 자치어의 성육이 부적합하 다고 알려져 있다(Kawaguchi et al., 1990; , 1996).
그런데 이번 조사기간 동안 거차군도 주변역(I해역)의 전 수층 평균수온은 13.1�C로 다른 해역에 비해 상당히 낮은 편이었다. 따라서 이 해역에서의 멸치의 산란제어 에는 우선 저수온이 크게 작용했으리라 추측된다. 그밖 에 다른 환경요인들도 멸치의 산란에 영향을 끼쳤겠지 만 조사해역내 멸치 난∙자치어 분포와 여러 환경요인 들과의 관계에 대한 면밀한 검토는 다음 논문에서 자세 히 다루기로 한다.
이번 연구에서 멸치 어란의 발생단계별 분포현황에 있어서는 산란된 후 얼마 지나지 않은 I단계 어란들이 청산도 주변역(J해역), 제주도 북서부 연안역과 인접한 제주해협 남서부역(C해역) 그리고 추자도 주변인 제주 해협 북서부역(A해역)에서 집중적으로 분포하고 있음을 확인할 수 있었다. 이러한 결과는 특히 이 해역들이 멸 치들의 주산란장이 되고 있음을 나타내는 것이라 할 수 있다. 또한 부화직전의 III단계 어란인 경우는 연구해역 내에서도 가장 동쪽인 초도와 거문도 주변해역(G해역) 에서 가장 많이 분포하는 등 전반적으로 연구해역의 동 쪽방향으로 갈수록 발생단계가 진행된 어란의 출현율이 증가하는 경향을 보이고 있었다. 기존의 표층부표나 인 공위성 추적부표등을 통한 해류패턴에 관한 연구결과에 의하면 일반적으로 여름철 제주해협을 포함한 남해 서 부해역에서의 해류패턴은 주로 동쪽으로 진행하거나 제 주해협을 통과하여 남해안으로 이동하는등 전형적으로 북동방향의 해류패턴이 우세하다고 보고되고 있다(김, 1982; 김, 1987; 김과 노, 1997). 따라서 이번 연구결과에 서 나타난 발생단계가 진행된 멸치 어란들의 출현율이 동쪽 해역으로 갈수록 증가하는 경향과 이러한 여름철 해류패턴을 함께 고려하여 생각해 본다면 이는 곧 연구 해역내에서 산란된 멸치 어란들이 난내 발생이 진행되 면서 시간이 경과함에 따라 해류의 흐름에 의해 점차 동쪽방향으로 수송되고 있음을 시사하는 것이라 할 수
있다.
한편, 멸치 자치어는 10 m, 20 m 수층에서 고밀도로 출 현하고 있었으며 특히 제주해협 북부역(A, B해역)에서 높은 출현밀도를 보이고 있었다. 여기서 제주해협 북서 부역(A해역)은 멸치의 주산란장 중 하나로 추정되는 해 역인데 이곳에서는 이번 조사기간 동안10 m, 20 m 수층 에서 비교적 강한 수온전선대(Fig. 2)가 형성되고 있었 다. 기존의 보고에 따르면 남해안 일대에서 멸치 난∙자 치어의 분포양상은 해역내 형성된 전선을 중심으로 양 분되어 분포하거나 전선상에 집중적으로 분포하는 등 해역내 전선의 형성 유무와 강약에 따라 그 분포가 크 게 달라지고 있음을 알 수 있다(김, 1983; 김과 최, 1988;
추와 김, 1998). 따라서 이번 연구결과에서도 산란장 근 처에서 부화한 멸치 자치어들이 부근 해역에 형성된 전 선등에 의한 해수장벽 효과나 수렴현상(Iles and Sinclair, 1982; , 1992)등과 같은 물리적 해수유동에 의해 이 해역에 집중적으로 분포하고 있는 것으로 추정 된다.
멸치 자치어들의 해역별 체급조성을 살펴본 결과, 제 주해협내인 경우에는 서부역(A, C해역)에 비해 동부 역(B, D해역)에서 10.0 mm TL 이상 개체들의 출현율이 높게 나타나고 있어 동쪽으로 갈수록 크기가 큰 개체의 출현비율이 증가하고 있음을 확인할 수 있었다. 이러한 현상은 앞서 언급한 어란의 경우와 마찬가지로 여름철 제주해협내에서의 탁월한 북동방향의 해류패턴에 의해 멸치 자치어들이 성장함에 따라 점차 북동쪽으로 수송 되고 있음을 시사하고 있는 것이라 생각된다. 이밖에, 제 주해협 북동부 외양역(G해역)과 청산도 주변역(J해역) 에서는 10.0 mm TL 미만의 개체들이 높은 비율(90% 이 상)로 출현하고 있었고 10.0 mm TL 이상 개체의 출현율 이 매우 낮았다. 조사 당시 이들 해역은 부화 직전의 III 단계 어란들이 대량으로 출현한 해역이었다. 따라서 이 들 해역내에는 갓 부화해서 유영력이 미약할 것이라 판 단되는 10.0m m TL 이하의 자치어들이 집중분포하고 있었던 것으로 추정되는 반면에 10.0 mm TL 이상의 개 체들은 조사 당시 이미 해류 등에 의해 이동되었기 때 문에 해역내 출현율이 매우 낮았던 것과 관계가 있다고 판단된다. 또한 제주해협 남서부 외양역(F해역)인 경우 는 10.0 mm TL 이상(81.5%)의 개체들이 상대적으로 높 은 출현율을 보였는데 이 해역에서 이러한 대형개체들 의 출현이 제주해협 북서부 외양역(E해역)에서 남하한 것인지 아니면 제주도 남서부 외양역으로부터 대마난류 에 의해 북상한 것인지에 대해서는 아직 확실치 않다. 앞으로 본 연구해역내에서 멸치 난∙자치어들의 분포 및 수송상황등을 더욱 더 자세하게 논의하기 위해서는
해역내 해수 유동상황과 같은 물리적 특성등의 조사가 동시에 이루어져야 할 것으로 사료된다.
요 약
2001년 7월 16부터 21일까지 제주해협을 포함한 남 해 서부 일대에서 멸치 난∙자치어의 분포현황에 대해 조사하였다. 망목 333µm와 505µm의 두 가지 네트를 통한 멸치 난∙자치어 출현량 비교에서는 멸치 어란인 경우 333µm네트에서가 출현량이 월등히 많았고 망목 간에도 유의한 차이를 보였으나 자치어에서는 유의차가 없었다.
멸치 어란 중 I단계 어란들은 청산도 주변역(J해역), 제주해협 남서부역(C해역) 그리고 추자도 부근의 제주 해협 북서부역(A해역)에서 고밀도로 출현하고 있어서 이 해역들이 멸치들의 주산란장임을 알 수 있었다. 부화 직전의 III단계 어란인 경우는 전반적으로 연구해역의 동쪽방향으로 갈수록 출현율이 증가하는 경향을 나타냈 다.
멸치 자치어들은 10 m, 20 m 수층에서 고밀도로 출현 하고 있었으며 특히 제주해협 북부역(A, B해역)에서 높 은 출현밀도를 보이고 있었다. 멸치 자치어들의 해역별 체급조성을 살펴본 결과 제주해협내인 경우에는 서부 역(A, C해역)에 비해 동부역(B, D해역)에서10.0 mm TL 이상 개체들의 출현율이 높게 나타나고 있었다.
사 사
이 논문은 2002년도 제주대학교 두뇌한국21사업에 의하여 지원되었습니다. 또한 본 연구는 2002년도 해양 수산부 수산특정과제 “제주도 주변해역의 난치자 수송 기구에 관한 연구(과제번호: 20020002)”의 일환으로 수 행되었음을 밝힙니다. 이번 연구에 적극적으로 협조해 주신 제주대학교 실습선 아라호 최찬문 선장님과 선원 여러분들께 진심으로 감사드립니다. 더불어 시료채집에 도움을 준 제주대학교 해양생태학연구실 김봉수, 조준, 김상현군을 비롯한 여러 연구실원들에게도 고마움을 전 합니다.
인 용 문 헌
Aoki, I. and K. Miyashita. 2000. Dispersal of larvae and
juveniles of Japanese anchovy Engraulis japonicus in the Kuroshio Extension and Kuroshio-Oyashio transi- tion regions, western North Pacific Ocean. Fisheries Research, 49 : 155~164.
Asahina, K. and I. Hanyu. 1983. Role of Temperature and photoperiod in annual reproductive cycle of the rose bitterling Rhodeus ocellatus ocellatus. Bull. Jap. Soc.
Sci. Fish., 52(6) : 965~969.
Bakun, A. and R. Parrish. 1991. Comparative studies of coastal pelagic fish reproductive habitats: the anchovy (Engraulis japonica) of the southeastern Atlantic. ICES J. Mar. Sci., 48 : 343~361.
Barnett, A., A.E. Jahn, P.D. Sertic and W. Watson. 1984.
Distribution of ichthyoplankton off San Onofre, Cali- fornia and methods for sampling very shallow coastal waters. Fish. Bull., 82(1) : 97~111.
Iles, T.D. and M. Sinclair. 1982. Atlantic herring: stock discreteness and abundance. Science, 215 : 627~633.
Kawaguchi, K., Y. Yamashita and A. Hayashi. 1990. Some aspects of spawning of the reared Japanese anchovy (Engraulis japonica) in relation to the photoperiod, water temperature and starvation. Bull. Japan. Soc.
Fish. Oceanogr., 54 : 364~372.
Kim, J.Y. 1992. Relationship between anchovy, Engraulis japonica, egg and larval density and environmental factors in the eastern waters of Korea. Bull. Korean Fish. Soc., 25(6) : 495~500.
Lenarz, W.H. 1972. Mesh retention of larvae of Sardinops caerulea and Engraulis mordax by plankton nets. Fish.
Bull., 70(3) : 839~848.
Razani, H. and I. Hanyu. 1986. Annual reproductive cycle of 2~3 years old female goldfish and its artifical modification by manipulations of water temperature and photoperiod. Bull. Jap. Soc. Sci. Fish., 52(6) : 965~969.
Weinstein, M.P., S.L. Weiss, R.G. Hodson and L.R. Gerry.
1980. Retention of three taxa of postlarval fishes in an intensively flushed tidal estuary, Cape Fear River, North Carolina. Fish. Bull., 78(2) : 419~436.
∙ . 1976. プランクトン 究 .
究 講 5, 共 , pp. 229
. 1996. 11 究 , 3. に
� するカタクチシラスの . 究,
60(4) : 418~419.
慶 . 1981. プランクトンの .
究 , 22 : 29~52.
∙ . 1999. におけるカ
タクチイワシの . 刊 , 31(4) : 222~226.
康 . 1992. 南 のカタクチイワシ及びマイワシ
の と 過 . 究, 56(4) :
526~536.
. 1992. フロント近 の 構 . 究
ノ-ト, 29 : 215~228.
谷 . 1987. ∙ および の .
と 49, 9(2) : 108~110.
국립수산진흥원. 1998. 연근해 주요 어종의 생태와 어장, pp.
96~102.
김 구. 1982. 한국 남서해의 해류분포와 그 영향에 관한 연 구. 서울대학교 연구보고서, pp. 89.
김 구. 1987. 제주도 주변 해류에 관한 연구의 현재와 미래. 제주도 연구, 4 : 83~97.
김상현∙노홍길. 1997. 제주해협의 해수유동특성에 관한 연 구. 한수지, 30(5) : 759~770.
김진영. 1983. 한국남해 및 서해 연안해역에서의 멸치 난∙치 어의 분포. 한수지, 16(4) : 401~409.
김진영∙강용주. 1992. 한국남해 멸치의 산란생태. 한수지, 25(5) : 331~340.
김진영∙김주일. 1991. 한국 남해안 멸치의 난∙자어 밀도와 가입. 수진연구보고, 45 : 101~107.
김진영∙김주일. 1993. 한국 남해 연안의 멸치 난∙자어 분포
와 사망률의 변동. 한어지, 5(2) : 217~225.
김진영∙최영민. 1988. 멸치, Engraulis japonicus난∙치어의 연직분포. 한수지, 21(3) : 139~144.
이은경∙유재명∙김 성∙이영철. 1996. 대한해협 멸치 치자 어의 수직분포. 한어지, 8(2) : 47~56.
장선덕∙홍성윤∙박청길∙진 평∙이병기∙이택열∙강용주
∙공 영. 1980. 멸치자원의 회유에 관한 연구. 부산수대 해연보, 12 : 1~38.
차성식. 2002. 우리 나라의 어류 초기 생태 연구 현황과 향후 과제. 한어지, 14(1) : 76~82.
최영민∙김진영. 1988. 한국남해산 멸치, Engraulis japonica 의 재생산력. 수진연구보고, 41 : 27~34.
추효상∙김동수. 1998. 한국 남해의 대마난류 변동이 멸치 난∙자어의 연안역 수송에 미치는 영향. 한수지, 31(2) : 226~244.
한경호∙신영호∙황동식. 2002. 고흥반도 연안에 분포하는 부 유성 난∙자치어 종조성의 계절 변동. 한어지, 14(1) : 45~52.
허성범∙유재명. 1984. 한국 서해안의 어류 난치어 분포. 한수 지, 17(6) : 536~542.
Received : June 2, 2002 Accepted : September 18, 2002