73 ISSN 1225-6552, eISSN 2287-7630
https://doi.org/10.7853/kjvs.2021.44.2.73
< Original Article >
Veterinary Service
Available online at http://kjves.org
*Corresponding author: Mi-Yeon Park, E-mail. [email protected] ORCID https://orcid.org/0000-0002-0215-1836
전북지역 돼지유행성설사 바이러스 Spike 유전자 염기서열 및 계통분석
박미연
*
ㆍ문보미ㆍ강수진ㆍ이종하ㆍ박진우ㆍ조성우ㆍ허철호 전라북도 동물위생시험소 북부지소Genetic sequence and phylogenetic analysis of spike genes of porcine epidemic diarrhea virus (PEDV) in Jeonbuk province
Mi-Yeon Park*, Bo-Mi Moon, Su-Jin Gang, Jong-Ha Lee, Jin-Woo Park, Sung-Woo Cho, Cheol-Ho Her
North-Branch, Jeonbuk Veterinary Service Laboratory, Iksan 54531, Korea (Received 9 February 2021; revised 18 June 2021; accepted 19 June 2021)
Abstract
Although many swine farms continuously vaccinated to sow to prevent Porcine epidemic diarrhea(PED), PED has occurred annually in swine herds in Jeonbuk province, Korea. In the present study, the small intestine and feces samples from 17 farms where severe watery diarrhea and death of newborn piglets occurred in 2019 were collected, amplified by RT-PCR and determined the complete nucleotide se- quences of the spike (S) glycoprotein genes of nine Jeonbuk PEDV isolates. The spike (S) glycoprotein is an important determinant for molecular characterization and genetic relationship of PEDV. These nine complete S gene isolates were compared with other PEDV reference strains to identify the molecular diversity, phylogenetic relationships and antigenicity analysis. 9 field strains share 98.5∼100% ho- mologies with each other at the nucleotide sequence level and 97.3∼100% homologies with each other at the amino acid level. The nine Jeonbuk PEDV isolates were classified into G2b group including a genetic specific signal, S-indels (insertion and deletion of S gene). In addition, comparisons the neutral- izing epitopes of S gene between 9 field strains and domestic vaccine strains of Korea mutated 12-15 amino acids with SM-98-1 (G1a group) and mutated 0-3 amino acids with QIAP1401 (G2b group).
Therefore, the development of G2b-based live vaccines will have to be expedited to ensure effective prevention of endemic PED in Korea. In addition, we will need to be prepared with periodic updates of preventive vaccines based on the PEDV variants for the re-emergence of a virulent strain.
Key words : PEDV, Spike gene, RT-PCR, Phylogenetic analysis
서 론
돼지 유행성설사는 모든 연령대의 돼지에 감염되 어, 식욕부진과 침울함을 동반한 수양성 설사와 구토 증상을 일으킨다(Debouck와 Pensaert, 1980). 생후 1주 일 내 신생 자돈의 경우 3∼4일간 중증의 수양성 설
사와 구토에 이어, 광범위한 탈수 및 전해질 불균형으 로 인해 폐사하게 된다. 폐사율은 평균 50%이나 1∼3 일령 신생 자돈의 경우에는 100%에 가까운 폐사율을 보이는 이 파괴적인 바이러스는 국내 양돈 농가에 지 속적인 재정적 손실을 끼치고 있다(Pijpers 등, 1993).
1971년 영국에서 최초 보고된 PEDV는 국내에서는 1992년에 처음 PED가 보고되었고, 이후 매년 발생하 여 지난 20년 동안 상당한 경제적 손실 야기로 국내
Table 1. General information on PED outbreaks farms in Jeonbuk province
Area Isolate strain Breeding scale Vaccination Age (days) Clinical signs (Heads)*
Gimje JBGJ-1 2,400 Killed-killed-live 7 386
Gimje JBGJ-2 2,700 Live-live-live 2∼5 160
Imsil IBIMS 2,100 Killed-killed-live 3∼14 281
Iksan JBIS-1 1,300 Live-live 5∼7 297
Iksan JBIS-2 1,100 Killed-killed-killed 5∼7 307
Iksan JBIS-3 1,000 Live-killed-killed-killed 3 145
Iksan JBIS-4 1,600 Killed-killed-killed-killed 6∼7 70
Jinan JBJA 1,700 Live-live 5∼7 320
Wanju JBWJ-1 1,100 Live-live 5∼7 230
*Clinical signs: Vomiting, Watery diarrhea and death.
양돈산업을 위협해 왔다(Kweon 등, 1993; Pijpers 등, 1993). 특히, 국내에서는 2013년 11월경 재출현한 PED 의 유행으로 우리나라 돼지 농장의 40% 이상이 피해 를 입었고, 2013년 이전까지 PED의 발생이 전혀 없었 던 미국에서도 2013년 5월 PED의 갑작스러운 출현으 로 1년간 800만 마리 이상의 신생 자돈들이 목숨을 잃었다. 현재, PEDV는 세계에서 가장 치명적인 돼지 바이러스 질병 중의 하나로, 우리나라는 물론 세계 돼 지고기 산업에서 심각한 경제적인 타격을 입히고 있 다(Lee와 Lee, 2014; Lee, 2015).
PED의 원인체는 Coronaviridae과 Alphacoronavirus 속 large enveloped RNA 바이러스로 PEDV 게놈의 길 이는 약 28 Kb이며, 7개의 ORFs (open reading frames, ORF1a, ORF1b, ORF2∼6)로 구성되어 있다. 게놈의 5’의 2/3를 차지하는 ORF1a, ORF1b는 ORF3와 함께 비구조 단백질을 암호화하고 있으며, 3’영역의 나머지 ORFs는 4개의 주요 구조 단백질인 glycosylated spike [S], Membrane [M], Envelope [E], Nucleocapsid [N]를 암호화하고 있다(Duarte 등, 1993). 이 중 PEDV의 S protein은 바이러스의 주요 envelope I glycoprotein으로 서, 세포 수용체와 상호작용하여 바이러스의 유입을 매개하고 자연 숙주의 항체 중화 유도를 자극하는 역 할을 한다(Chang 등, 2002). 따라서, 가장 많은 아미노 산의 변형이 축적되어 있는 N-terminal region을 포함 한 PEDV S glycoprotein은 PEDV 분리주들 사이의 유 전적 연관성과 특성을 판단하고, 유효한 백신 개발에 활용하기 적합한 바이러스 유전자로 알려져 있다(Lee 등, 2010; Oh 등, 2014).
현재 우리나라에서 PED의 예방은 모돈에게 백신을 접종하여 모체항체가 자돈으로 이행되는 방법을 선택 하고 있으며 대부분 농가에서 백신을 접종함에도 불 구하고, 매년 전북 도내 많은 농가에서 PED가 지속적
으로 발생하고 있는 실정이다. 따라서, 본 연구에서는 2019년 전북지역 PED 발생 농가에서 분리한 PEDV S gene의 유전자 염기서열 및 아미노산을 분석하고, 기 존 국내외 분리주 및 백신주와의 유전적 특성을 비교 분석 하고자 한다.
재료 및 방법
시료채취 및 RNA 추출
2019년 전북지역에서 PED로 진단된 17 농가의 분 변 및 소장 조직 유제시료를 채취하여 PBS에 10%
W/V로 만들어 3,000 rpm에서 10분간 원심분리 한 후 상층액을 확보하였다(Table 1). 이 상층액을 Maxwell RSC Viral Total Nucleic acid Purification Kit (Promega, USA)를 이용하여 제조사가 제시한 실험 방법에 따라 RNA를 추출하여 실험에 사용하였다.
RT-PCR 검사
PEDV S gene의 증폭에 사용한 primer는 Park 등 (2016)과 동일하게 합성하였으며, 합성한 primer로 증 폭이 안되는 부위는 Genbank의 data (MH891590)를 기 초로 primer를 제작하여 사용하였다(Cosmo-genetech, Korea) (Table 2). cDNA의 합성은 iScript cDNA Synthesis kit (Biorad Laboratories, USA)를 이용하였으며, −20°C 에서 보관 후 PCR에 사용하였다. PCR은 cDNA tem- plate 2 μL (5 μg/μL), 10×SP Buffer 2.5 μL, 2.5mM dNTP 2 μL, primer (forward and reverse mixture) 1 μL, SP-taq 0.25 μL에 총 반응용액 25 μL가 되도록 증류수 를 첨가한 후 PCR을 실시하였다. PCR 반응은 94°C 5
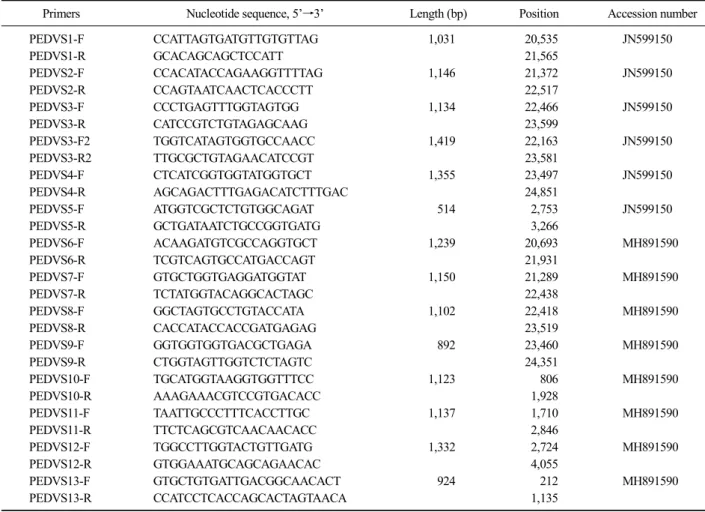
Table 2. List of primers utilized in this study
Primers Nucleotide sequence, 5’→3’ Length (bp) Position Accession number
PEDVS1-F CCATTAGTGATGTTGTGTTAG 1,031 20,535 JN599150
PEDVS1-R GCACAGCAGCTCCATT 21,565
PEDVS2-F CCACATACCAGAAGGTTTTAG 1,146 21,372 JN599150
PEDVS2-R CCAGTAATCAACTCACCCTT 22,517
PEDVS3-F CCCTGAGTTTGGTAGTGG 1,134 22,466 JN599150
PEDVS3-R CATCCGTCTGTAGAGCAAG 23,599
PEDVS3-F2 TGGTCATAGTGGTGCCAACC 1,419 22,163 JN599150
PEDVS3-R2 TTGCGCTGTAGAACATCCGT 23,581
PEDVS4-F CTCATCGGTGGTATGGTGCT 1,355 23,497 JN599150
PEDVS4-R AGCAGACTTTGAGACATCTTTGAC 24,851
PEDVS5-F ATGGTCGCTCTGTGGCAGAT 514 2,753 JN599150
PEDVS5-R GCTGATAATCTGCCGGTGATG 3,266
PEDVS6-F ACAAGATGTCGCCAGGTGCT 1,239 20,693 MH891590
PEDVS6-R TCGTCAGTGCCATGACCAGT 21,931
PEDVS7-F GTGCTGGTGAGGATGGTAT 1,150 21,289 MH891590
PEDVS7-R TCTATGGTACAGGCACTAGC 22,438
PEDVS8-F GGCTAGTGCCTGTACCATA 1,102 22,418 MH891590
PEDVS8-R CACCATACCACCGATGAGAG 23,519
PEDVS9-F GGTGGTGGTGACGCTGAGA 892 23,460 MH891590
PEDVS9-R CTGGTAGTTGGTCTCTAGTC 24,351
PEDVS10-F TGCATGGTAAGGTGGTTTCC 1,123 806 MH891590
PEDVS10-R AAAGAAACGTCCGTGACACC 1,928
PEDVS11-F TAATTGCCCTTTCACCTTGC 1,137 1,710 MH891590
PEDVS11-R TTCTCAGCGTCAACAACACC 2,846
PEDVS12-F TGGCCTTGGTACTGTTGATG 1,332 2,724 MH891590
PEDVS12-R GTGGAAATGCAGCAGAACAC 4,055
PEDVS13-F GTGCTGTGATTGACGGCAACACT 924 212 MH891590
PEDVS13-R CCATCCTCACCAGCACTAGTAACA 1,135
분간 1회, 94°C 30초, 52°C 30초, 72°C 1분 30초간 40 회, 72°C 5분간 1회 조건으로 실시하였다. 증폭산물은 1.5% agarose gel 상에서 전기영동을 한 뒤 ethidium bromide로 염색하여 band를 확인하였으며 증폭된 유 전자의 크기는 100bp DNA ladder (Bioneer, Korea)를 같이 전기영동 하여 확인하였다. 이후, PEDV 염기서 열을 분석하기 위하여, PCR 반응산물을 Gel extraction kit를 사용하여 정제하였다(Park 등, 2016).
다중 정렬 및 계통학적 분석
최근 국내에서 유행하는 PEDV와 기존에 보고된 PEDV의 S 유전자를 분석하기 위하여 BioEdit 7.2.5.0 와 Clustal W (Larkin 등, 2007)를 사용하였으며, 염기서 열과 아미노산 서열 다중 정렬을 통해 상동성을 비교하 여 백분위로 나타냈다. 계통학적 분석을 위하여 MEGA software version 7 (Kumar 등, 2016)을 이용하여 neigh- bor-joining (NJ) method로 1,000회의 bootstrap을 실시
하여 계통수를 생성하였으며, 분석된 유전자 서열은 GenBank에 공개된 자료와 비교하였다(Table 3).
결 과
PEDV S 유전자의 염기서열 및 아미노산 분석
본 연구에서 분리된 9개의 PEDV S 유전자 전체 염 기서열을 분석한 결과, 4,161 bp 길이의 nucleotide로 구성되어 있었으며 1,386개의 아미노산을 암호화하고 있는 것으로 밝혀졌다. 분리된 PEDV S 유전자를 CV777 strain의 염기서열과 비교시 9개의 분리주 모두 9 nu- cleotide (3 아미노산)가 더 길었는데, 이러한 결과는 S1 도메인의 N-terminal 영역에 아미노산의 삽입 또는 삭제가 존재하기 때문이다. 또한, 분리된 PEDV S 유 전자 모두에서 57∼58번 위치에 4개의 아미노산 삽 입, 135∼136번 위치에 1개의 아미노산 삽입, 158번과
Table 3. The recent porcine epidemic diarrhea virus (PEDV) isolates in Jeonbuk and references strains used in this study
Strain Country and year Accession number
CV777 Belgium, 1978 AF353511
SM98-1* Korea, 1998 GU937797
Virulent_DR-13 Korea, 1999 JQ023161 Attenuated_DR-13* Korea, 1999 JQ023162
Spk1 Korea, 2002 AF500215
TGEV USA, 2006 DQ811788
KNU-0801 Korea, 2008 GU180142
KNU-0901 Korea, 2009 GU180144
CH/ZMDZY/11 China, 2011 KC196276
GD-1 China, 2011 JX647847
LC China, 2011 JX489155
AH2012 China, 2012 KC210145
CH/FJZZ-9 China, 2012 KC140102
Colorado USA, 2013 KF272920
IA1 USA, 2013 KF468753
KNU-1409-1 Korea, 2014 KJ741221
QIAP1401* Korea, 2014 KX793713
KNU-1601 Korea, 2016 KY963963
PC273/O USA, 2017 MG837058
KNU-1703 Korea, 2017 MH052682
KNU-1705 Korea, 2017 MH052684
KNU-1706 Korea, 2017 MH052685
KNU-1708 Korea, 2017 MH052687
KNU-1802 Korea, 2018 MH243314
S10 Korea, 2018 MH891590
JBGJ-1† Korea, 2019 This study
JBGJ-2† Korea, 2019 This study
JBIMS† Korea, 2019 This study
JBIS-1† Korea, 2019 This study
JBIS-2† Korea, 2019 This study
JBIS-3† Korea, 2019 This study
JBIS-4† Korea, 2019 This study
JBJA† Korea, 2019 This study
JBWJ-1† Korea, 2019 This study
*Korean PEDV vaccine strains. †Jeonbuk PEDV isolates sequenced in this study.
159번 위치의 2개의 아미노산 결손을 포함하는 S in- dels (S insertion-deletions)라고 하는 G2b 그룹의 뚜렷 한 유전적 변이가 존재하였다(Fig. 1).
염기서열과 아미노산 상동성을 분석한 결과, 분리 주간 염기서열 상동성은 98.5∼100%, 아미노산 상동 성은 97.3∼100%의 상동성을 보였으며, 그 중 JBJA과 JBWJ-1주가 가장 높은 100%의 상동성을 보였다.
GenBank의 data (Table 3)와 분석 결과, 가장 높은 상 동성을 보이는 주는 PC273/O (미국, 2017)로 염기서열 상동성은 98.7∼99.9%, 아미노산 상동성은 97.6∼99.7%
로 나타났으며, 국내 분리주 중에서는 KNU-1802 (한 국, 2017)가 염기서열 상동성 98.9∼99.7%, 아미노산
상동성 98.1∼99.3%를 보였다. 반면, 국내 시판 백신 주(SM-98-1, DR-13, QIAP1401)와의 염기서열 및 아미 노산 상동성은 QIAP1401주가 각각 98.8∼99.5%, 97.8∼
98.8%로 가장 높은 상동성을 보였고, SM-98-1과 DR- 13주는 각각 93.4∼93.9%, 93.5∼94.0%의 염기서열 상 동성과 88.9∼89.5%, 88.6∼89.1%의 아미노산 상동성 을 보였다(Table 4).
Phylogenetic tree 분석
PEDV S 유전자의 전체 염기서열을 이용하여 국내 외 분리주와 phylogenetic tree를 분석한 결과, 본 연구 에서 분리된 9개 바이러스주 모두 genogroup 2b (G2b) 계열에 속하였다. G2는 2개의 subgroup (G2a, G2b)으 로 나뉘는데, G2b 그룹은 국내 분리주(KNU-1706/2017, S10/2018, KNU-1409-1/2014 등)와 미국 분리주(PC273/
O/2017, IA1/2013 등) 및 중국 분리주(AH2012/2012, GD-1/2011 등)를 포함하여 2014년 이후 국내에서 유 행하는 PEDV 야외주들이 모두 이 그룹에 속해 있다.
한편, 국내 백신주로 많이 사용되고 있는 SM-98-1과 DR-13주는 G1 계열로 현재 유행하고 있는 PEDV와는 계통발생학적으로 차이가 있었으나, QIAP1401주는 국내 발생 야외주와 동일한 G2b 계열로 분류할 수 있 었다(Fig. 2).
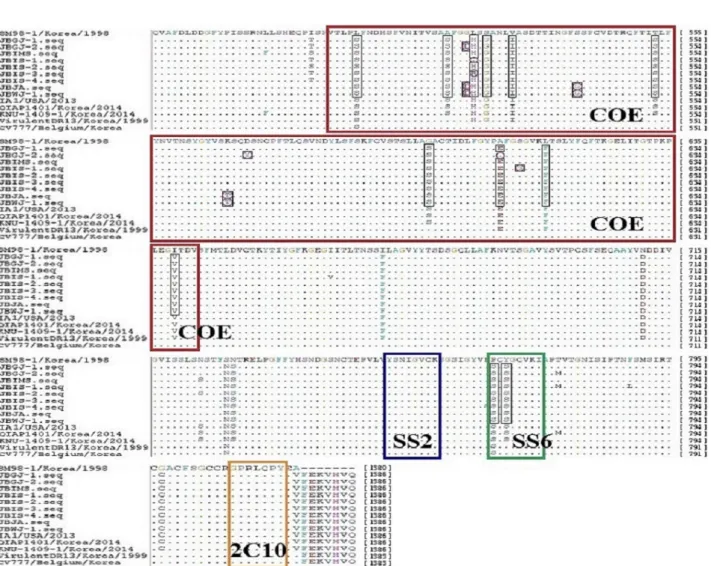
S 유전자의 항원성 분석
S 유전자의 항원성에 대한 변이를 조사하기 위해 neutralizing epitope으로 밝혀진 COE (503∼642 aa), SS2 (752∼759 aa), SS6 (768∼775 aa), 2C10 (1,372∼1,378 aa) 부위를 초기 백신주 SM-98-1주와 비교 분석하였 다. COE domain에서, 분리된 9주 모두 변이된 부위는 8개(L→S at 507, A→S at 521, S→G at 527, V→I at 531, T→S at 553, G→S at 598, L→F at 616, I→V at 639)였으며, 8주가 변이된 부위는 2개(L→H, R at 525, A→E at 609), 3주가 변이된 부위는 1개(G→D at 524), 2주가 변이된 부위는 2개(S→G at 541, K→T at 567), 1주만 변이된 부위는 2개(D→Y at 570, S→G at 612) 였다. SS6 region에서는 분리된 9주 모두 2개(P→S at 768, Y→S at 770) 부위에서 아미노산 변이가 관찰되 었으며, SS2와 2C10에서는 어떠한 변이도 관찰되지 않았다. 한편, 국내 백신주 QIAP1401와의 neutralizing epitope 부위에 대한 비교 결과, COE 부위에서 3주가 변이된 부위는 1개(G→D at 523), 2주가 변이된 부위
Fig. 1. Alignment of amino N-terminal 1-234 amino acid of S proteins between the reference strains and 9 Jeonbuk strains. The dashes (−) in- dicate the deleted sequences. Insertions and deletions within PEDV isolates are indicated by boxes (Red box indicate the inserted amino acids com- pared with CV777. Blue box indicate the deleted amino acids compared with CV777).
는 3개(H→L, R at 524, S→G at 540, K→T at 566), 1 주만 변이된 부위는 3개(D→Y at 569, E→A at 608, S→
G at 611)가 관찰되었고, SS2, SS6, 2C10 부위에서는 변이가 관찰되지 않았다(Fig. 3).
고 찰
PED는 우리나라에서 2014년 이후 매년 지속적으로 발생하고 있으며, 대부분은 낮은 폐사율을 동반한 상 재성 재발 사례를 보이나, 일부에서는 높은 폐사율을 동반한 급성 유행성 신규 발생이 보고되고 있다. 2013∼
2014년 PED의 국내 유행은 국가 재난 수준으로 약 50%의 양돈농장이 피해를 입었고, 지난 10년간 국내 양돈산업에 심각한 경제적 피해를 입혔다(Lee, 2015).
PEDV는 단일 혈청형이지만, 유전학적으로 크게 고 전형(classical strain) 바이러스들이 포함된 genogroup 1 (G1)과 야외유행주(epidemic strain) 및 대유행주(pan- demic strain)가 포함된 genogroup 2 (G2)로 나눌 수 있 다. G1 그룹은 저병원성으로 고전주 아형그룹인 G1a 와 G1a/G2b의 재조합 형태인 G1b로, G2 그룹은 고병
원성으로 2013년 대유행 이전 야외주 G2a 및 대유행 주 유래 최근 야외주 G2b로 구성되어 있다(Lee와 Lee, 2014).
본 연구에서 분리된 9주의 전체 S 유전자 염기서열 및 아미노산 분석 결과, 최근 국내 야외주들과 같은 G2b 그룹을 형성하고 있었으며, 분리주간 염기서열 상동성은 98.5∼100%, 아미노산 상동성은 97.3∼100%
로 전북 분리주들은 서로 높은 상동성을 가진 것으로 나타났다. 특히, JBJA과 JBWJ-1 주는 유전자 분석 결 과 100%의 상동성을 보였는데, 발생 시기가 유사하 고, 지리적으로 인접한 지역임을 고려할 때 역학적인 연관성에 대한 추가 조사가 요구된다. 또한, GenBank data와 비교시, 국내 분리주에서는 KNU-1802 (한국, 2017), 국내 분리주 외에서는 PC273/O (미국, 2017)주 가 가장 높은 상동성을 보여, 앞서 보고된 바와 같이 최근 국내 유행주는 미국 유래주와 유사성이 높았으 며 이는 유입된 미국 유래주가 2014년 이후 국내에서 계속 변이하고 있음을 추정할 수 있다. 또한, CV777 주와 달리 본 연구에서 분리된 9개 주는 모두 4,161 bp 길이의 염기서열로 구성된 1,386개의 아미노산을 암호화하고 있었으며, G2b 그룹의 특이적인 유전적
Table4.Comparison of the nucleotide and deduced amino acid sequences of S genes of PEDV references strains and PEDV isolates in Jeonbuk Virus strain123456789101112131415161718192021222324252627282930313233 1. JBGJ-199.597.799.397.799.799.798.598.598.599.398.899.399.297.997.898.198.298.297.997.497.296.895.996.396.189.689.990.889.391.289.688.8 2. JBGJ-299.797.599.197.599.599.798.698.698.399.198.699.099.197.897.698.198.298.297.997.697.196.995.996.295.989.890.191.089.191.089.688.8 3. JBIMS98.798.597.399.797.797.797.497.497.498.197.897.898.099.799.698.898.998.998.898.197.997.596.596.996.489.890.390.989.591.489.889.0 4. JBIS-199.799.698.597.399.499.398.198.198.199.098.598.898.997.697.497.897.897.897.697.197.096.495.596.095.789.389.690.588.990.889.488.6 5. JBIS-298.898.699.798.697.797.797.497.497.498.197.897.898.099.799.698.898.998.998.898.197.997.596.596.996.490.090.391.189.491.589.989.1 6. JBIS-399.899.898.699.798.799.798.598.598.599.398.899.199.297.997.898.198.298.297.997.597.496.895.996.396.189.790.090.989.391.289.788.9 7. JBIS-499.999.898.799.798.899.898.698.698.599.398.899.199.297.997.898.398.498.498.197.697.297.096.196.396.189.790.090.989.491.289.688.8 8. JBJA99.199.198.799.098.899.199.2100.099.498.899.398.598.797.797.598.298.398.398.097.597.297.095.996.095.889.689.990.789.191.089.488.6 9. JBWJ-199.199.198.799.098.899.199.2100.099.498.899.398.598.797.797.598.298.398.398.097.597.297.095.996.095.889.689.990.789.191.089.488.6 10. S10/Korea/2018/ MH89159099.199.098.699.098.799.099.199.699.698.899.498.598.797.797.597.998.098.097.897.297.196.795.695.995.789.389.690.589.090.889.288.4 11. KNU-1802/Korea/ 2018/MH24331499.699.598.999.599.099.699.799.399.399.299.199.799.998.498.298.698.798.798.597.897.897.196.396.696.490.290.591.489.991.890.189.3 12. KNU-1703/KOREA/ 2017/MH05268299.399.298.899.298.999.299.399.699.699.799.498.899.198.097.898.398.498.498.197.697.497.196.096.396.189.689.990.889.391.289.688.8 13. KNU-1705/KOREA/ 2017/MH05268499.599.398.999.399.099.499.599.299.299.299.799.499.698.097.898.398.498.498.197.497.496.895.996.396.190.090.391.289.791.489.788.9 14. KNU-1706/KOREA/ 2017/MH05268599.699.598.999.599.099.699.699.299.299.299.999.499.798.398.198.598.698.698.497.797.797.196.296.596.390.190.491.389.891.790.089.2 15. PC273/O/USA/ 2017/MG83705898.998.799.898.799.998.898.998.998.998.899.199.099.199.199.899.199.199.199.198.498.297.896.897.196.790.190.491.289.691.790.189.3 16. KNU-1708/KOREA/ 2017/MH05268798.898.799.798.799.898.798.898.898.898.799.199.099.099.099.998.999.099.098.998.298.097.696.697.096.590.190.491.289.491.589.989.1 17. QIAP1401/Korea/ 2014/KX79371398.998.999.498.899.598.899.099.099.098.999.299.199.199.199.599.599.999.999.799.098.798.497.597.797.290.590.791.690.092.190.589.7 18. Colorado/USA/2013/ KF27292098.998.999.498.899.598.999.099.199.198.999.299.199.299.299.699.599.9100.099.799.198.898.597.697.897.390.590.891.790.192.290.589.8 19. IA1/USA/2013/ KF46875398.998.899.498.899.598.999.099.099.098.999.299.199.199.199.699.599.9100.099.799.198.898.597.697.897.390.590.891.790.192.290.589.8
Table4.Continued Virus strain123456789101112131415161718192021222324252627282930313233 20. KNU-1409-1/Korea/ 2014/KJ74122198.798.699.398.699.398.698.898.898.898.799.098.999.099.099.499.499.799.899.799.398.598.497.397.597.190.390.591.489.891.990.389.5 21. KNU-1601/Korea/ 2016/KY96396398.598.599.098.499.198.498.598.698.698.498.698.698.698.699.199.199.499.499.499.697.997.896.696.896.390.090.391.189.691.790.289.4 22. CH/ZMDZY/11/ China/2011/KC19627698.498.398.998.399.098.498.498.598.598.498.798.698.698.699.199.099.399.399.399.298.997.797.097.196.790.490.591.289.591.790.289.5 23. AH2012/China/2012/ KC21014598.198.098.697.998.798.098.298.398.398.198.398.398.398.398.798.799.099.199.199.198.898.897.297.296.890.590.791.690.192.290.689.9 24. CH/FJZZ-9/China/ 2012/KC14010297.397.397.797.297.897.397.497.497.497.297.597.597.497.597.997.898.298.398.298.197.998.098.197.997.590.090.391.089.291.389.789.0 25. LC/China/2011/ JX48915597.197.097.597.097.797.097.197.297.297.197.397.397.297.297.797.697.998.098.097.997.697.897.998.498.590.490.691.589.791.890.589.8 26. GD-1/China/2011/ JX64784797.096.997.396.997.597.097.097.097.096.997.297.197.197.197.597.497.797.897.897.897.597.697.798.299.190.590.791.689.892.090.589.8 27. Spk1/Korea/2002/ AF50021594.094.094.293.994.494.094.194.194.194.094.294.394.394.294.494.494.694.794.694.594.494.594.694.494.494.497.595.991.591.890.489.7 28. KNU-0801/Korea/ 2008/GU18014294.394.394.694.294.794.394.494.494.494.394.594.594.594.594.794.794.895.094.994.894.794.794.894.794.794.698.997.591.891.690.589.9 29. KNU-0901/Korea/ 2009/GU18014494.894.894.994.795.194.894.994.994.994.895.095.095.095.095.195.195.295.495.395.295.095.095.395.095.094.997.397.991.892.291.090.3 30. AttenuatedDR-13/ Korea/1999/JQ02316293.693.593.893.594.093.593.693.693.693.693.993.793.993.894.094.094.294.294.294.194.194.194.394.094.093.994.794.994.797.094.693.8 31. VirulentDR-13/ Korea/1999/JQ02316194.894.795.094.795.294.894.894.894.894.795.194.995.095.095.295.295.395.495.495.395.295.395.495.295.295.195.395.395.398.397.696.9 32. CV777/Belgium/ 1978/AF35351193.893.794.093.794.293.793.893.893.893.794.093.993.993.994.294.294.394.494.494.394.294.394.494.294.494.194.594.694.496.898.599.3 33. SM98-1/Korea/1998/ GU93779793.593.493.693.493.993.493.593.493.493.493.793.693.693.693.993.994.094.194.193.993.993.994.193.994.293.994.294.394.196.598.299.7 The percent identify was shown in the lower left (nucleotide) and the upper right (deduced amino acid).
Fig. 2. Phylogenetic analysis based on the nucleotide sequence corresponding to the full-length S gene of PEDV strains. Sequencing alignments were performed using the Clustal W program, and the phylogenetic tree was constructed from the aligned nucleotide sequences by using the neigh- bor-joining method. The bootstrap values were determined from 1,000 replicates of original data. the scale bars indicate the number of 0.1 inferred substitutions per site (Putative similar regions of S gene of other distantly related coronavirus, transmissible gastroenteritis virus (TGEV), was also included in this study).
특징인 S-indels가 모두에서 관찰되었다(Lee와 Lee, 2014; Park 등, 2016).
전북지역에서 분리된 PEDV와 초기 국내 백신주 SM-98-1 및 DR-13주와의 염기서열 및 아미노산 서열 비교 결과, 염기서열에서는 각각 6.1∼6.6%, 10.5∼
11.1%, 아미노산에서는 각각 10.5∼11.1%, 10.9∼11.4%
의 차이를 보였다. 한편, 국내 백신주 QIAP1401와의 비교에서는 염기서열은 0.5∼1.2%, 아미노산 서열은 1.2∼2.2%의 차이로 나타나, G1a 그룹의 기존 백신주 들과 달리 높은 일치율을 보였다.
또한, 중화항체 생성에 관여하는 것으로 알려진 COE, SS2, SS6, 2C10 부위를 초기 국내 백신주 SM-
98-1과 비교 결과, COE 부위에서는 9개의 분리주 모 두 변이된 8개 부위를 포함하여 총 15개의 변이가 관 찰되었고 SS6에서는 9개 분리주가 2개 부위에서 모두 변이되었으나, SS2와 SS10에서는 변이가 관찰되지 않 았다(Sun 등, 2012; Park 등, 2016). 반면, 국내 백신주 QIAP1401와의 neutralizing epitope 부위에 대한 비교 결과, COE domain에서 총 7개 부위가 변이되었으나, 3주가 변이된 부위 1개, 2주가 변이된 부위 3개, 1주 만 변이된 부위 3개로 변이 정도가 크지 않았고, SS2, SS6, 2C10 부위에서는 변이가 관찰되지 않았다. 이러 한 S 유전자에서의 염기서열의 삽입, 결손 및 항원 부 위의 변이는 바이러스의 항원성 및 중화 작용에 변화