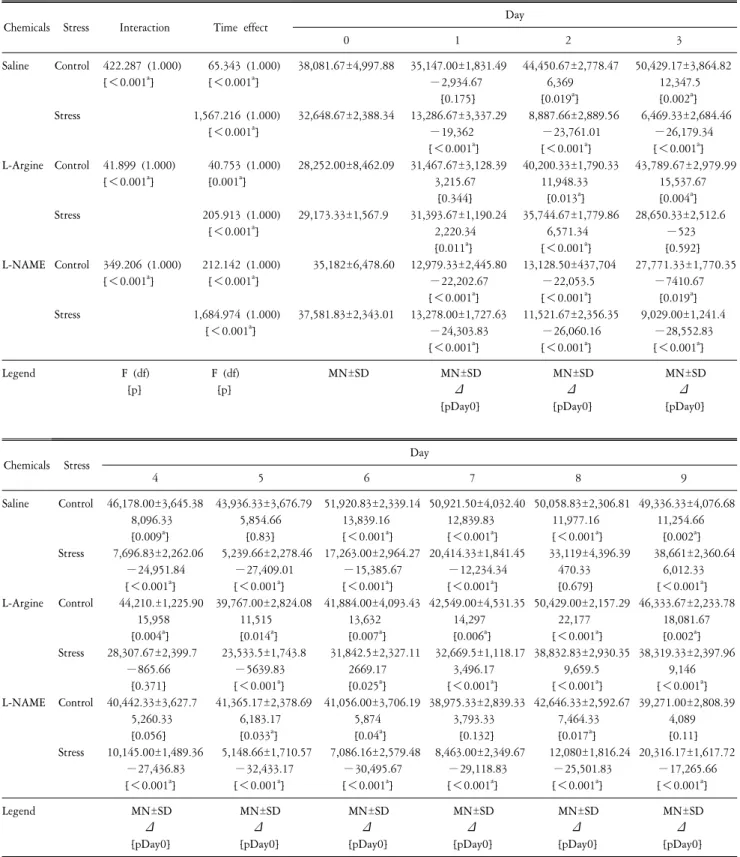
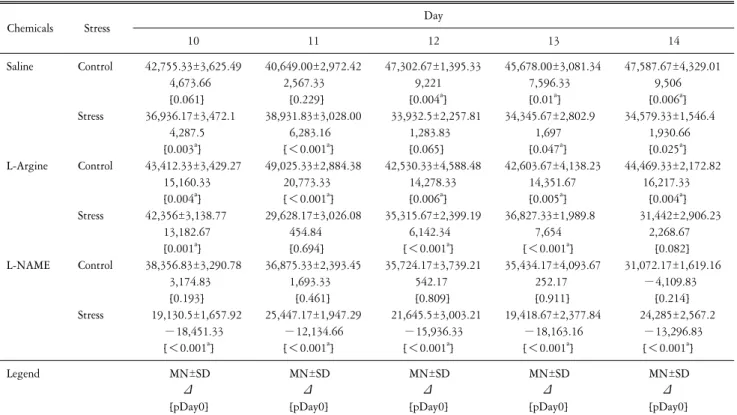
Time Course of the Effects of Nitric Oxide on Voluntary Wheel Running Behavior following Restraint Stress
8
0
0
전체 글
(2)
(3)
(4)
(5)
(6)
(7)
(8)
수치

관련 문서
■ 행렬 A을 가역행렬(invertible matrix) 또는 정칙행렬(nonsingular matrix)이라 한다.. 프로그램의 running time 계산하기. 34.. 프로그램의
1 John Owen, Justification by Faith Alone, in The Works of John Owen, ed. John Bolt, trans. Scott Clark, "Do This and Live: Christ's Active Obedience as the
Also by analysing changes of educational process, the seventh education course, and the direction of educational revision, I closely examined music
The objective of this study is to understand the effects of walking activity using fairy tales on the peer competence and emotional intelligence of
The Study on the Changes of Bacterial Inorganic Phosphate Metabolism in Interactions between Nitric Oxide and Salmonlla enterica serovar Typhimurium..
The purpose of this research is to confirm whether voluntary early morning physical exercise effects on the emotional and behavioral development of
Findings from a six-year longitudinal study which examined the effects over time of using the William and Mary language arts for gifted learners in a
The purpose of this study was to examine the effects of football on PAPS in sports club activity for 12 weeks in middle school boys.. The following
