Allergy Immunity Regulation and Synergism of Bifidobacteria
Kwang Keun Cho
1and In Soon Choi
2*
1
Department of Animal Resources Technology, Gyeongnam National University of Science and Technology, Gyeongnam 660-758, Korea
2
Department of Biological Science, Silla University, Busan 617-736, Korea
Received March 15, 2017 /Revised April 26, 2017 /Accepted April 27, 2017
Allergic diseases have increased over the past several decade worldwide including developing countries.
Allergic inflammatory responses are caused by Th (T helper)2 immune responses, triggered by aller- gen ingestion by antigen presenting cells such as dendritic cells (DCs). Intestinal microorganisms con- trol the metabolism and physiological functions of the host, contribute to early immune system matu- ration during the early life, and homeostasis and epithelial integrity during life. Bifidobacteria have strain-specific immunostimulatory properties in the Th1/Th2 balance, inhibit TSLP (thymic stromal lymphopoietin) and IgE expression, and promote Flg (Filaggrin) and FoxP3 (Treg) expression to alle- viate allergies. In addition, unmethylated CpG motif ODN (oligodeoxynucleotides) is recognized by TLR (toll-like receptors)9 of B cells and plasmacytoid dendritic cells (pDCs) to induce innate and adaptive immune responses, while the butyrate produced by Clostridium butyricum activates the GPR (G-protein coupled receptors)109a signaling pathway to induce the expression of anti-inflammatory gene of pDCs, and directly stimulates the proliferation of thymically derived regulatory T (tTreg) cells through the activation of GPR43 or inhibits the activity of HADC (histone deacetylase) to differentiate naive CD4⁺ T cells into pTreg cells through the histone H3 acetylation of Foxp3 gene intronic enhancer.
Key words : Allergy, Bifidobacteria, Butyrate, Clostridium butyricum, CpG ODN
*Corresponding author
*Tel : +82-51-999-5348, Fax : +82-51-999-5644
*E-mail : [email protected]
This is an Open-Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/3.0) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
Journal of Life Science
2017 Vol. 27. No. 4. 482~499 DOI : https://doi.org/10.5352/JLS.2017.27.4.482서 론
음식 allergy 발병률은 지난 20년 동안 꾸준히 증가 하고 있으며, 현재 중요한 공중 보건의 문제(major public health issue)로 대두되고 있다[89]. 그 원인으로서 과도한 위생(exces- sive hygiene)이 allergy 발병률을 증가 시키는 것으로 설명되 어 왔으나[1], 최근에는 장내 미생물 조성(gut microflora com- position) 변화가 또 다른 원인으로 제기되고 있다[64]. 무균 (germ-free) 쥐에서 allergy 발생이 더욱 잘 일어나며, 특정 probiotics 급여 시 allergy 증상이 완화될 수 있다는 것이 이러 한 원인을 잘 설명해 준다[2].
식이 섬유소(dietary fiber)는 대장(colon)의 혐기적 미생물 (anaerobic bacteria)에 의하여 acetate와 propionate, butyrate 와 같은 단쇄지방산(short-chain fatty acids)으로 발효된다. 이 들 단쇄지방산은 GPR41과 GPR43, GPR109A와 다양한 친화 력으로 결합하며, 이러한 수용체는 면역 세포뿐만 아니라 상 피세포에서도 발현된다. 서구 식단(western diets)은 전형적으
로 지방이 높고 섬유소가 낮은데 이러한 것은 장내 미생물 환경(gut bacterial ecology)과 상피 총체성(epithelial in- tegrity), Treg 세포 발달과 연관되어 있고[23], 음식 allergy의 원인이 될 수 있다. 상피 총체성이 붕괴되어 IL (interleukin)- 33이 방출되고 과도한 TSLP가 생산되면 수지상 세포가 Th (T helper)2-like phenotype으로 바뀌어 allergy 감작(allergic sensitization)을 유발한다[29]. 또한 장염(intestinal inflamma- tion)이 발생하면 CD103
+DCs의 Treg cell 분화(differentia- tion) 촉진 능력을 무효화 시킬 수 있다[45].
Bifidobacteriales (order)는 사람이나 다른 포유류 또는 새나
벌의 장내(gastrointestinal tract) 존재하는 중요한 미생물로
Actinobacteria phylum과 Bifidobacteriaceae family에 속하며, 8개의 genera (Aeriscardovia, Alloscardovia, Bifidobacterium, Ga-
rdnerella, Pseudoscardovia, Neoscardovia, Parascardovia, Scardovi)로 나누어지고 Bifidobacterium genus는 39 species와 9 sub- species로 나누어진다[55]. Bifidobacteria는 Gram-positive anaerobic bacteria로서 모유 급여 유아(breast-fed babies)의 경유 장내 미생물(total gut microbiota)의 90%를 차지하고, 분 유 급여 유아(formula-fed babies)의 경우는 75%를 차지한다 [26]. Bifidobacteria는 로타바이러스(rotavirus)나 항생제 관련 설사, 염증성 장 질환[66], 괴사성 장염(necrotizing enter- ocolitis), allergy 등을 포함하는 다양한 질병에 대하여 효능을 나타내는 유익한 균이며[32], Th1/Th2 balance에 대한 strain- specific 효과를 나타낸다[52]. 이러한 strain-specific 효과의 차
- Review -
Fig. 1. Early life impacts of microbial colonization upon immune development in the lung and gut. Before birth there is a negligible load of bacteria present in mucosal tissues. Immune cells present at these sites can broadly be considered ‘immature’ and have a functionality that is representative of the stimuli present in the intrauterine environment. Immediately following birth, mucosal tissues are exposed to, and colonized by, bacteria. These early host–microbe interactions could be critical in shaping the trajectory of an individual’s immune development, and consequently risk of disease. Formation of a balanced and diverse microbiota is associated with ‘healthy’ immune development and low risk of allergies, whilst a lack of microbial diversity and an over-representation of bacteria, typically considered as pathogens, is associated with the development of allergy [51].
이는 TLR에 의해서 인식되는 미생물의 구조적 차이(bacterial structures)에 기인한다[52]. 면역 자극 특성(immunostimula- tory properties)은 미생물 염색체(bacterial genome)에 보존 되어 있는 높은 GC 함량 즉 unmethylated CpG motifs와 관련 되어 있으며, CpG motif가 TLR9을 활성화 시키고 Th1-type 면역 반응(immune responses)을 유도한다[90]. CpG motif는 진핵생물(CpG motifs의 10~30%가 unmethylated)보다 원핵 생물(CpG motifs의 95%가 unmethylated)에서 더 많이 존재 한다[41]. 50종의 Bifidobacteria 중에서 6종의 genome sequence 가 밝혀졌으며, 평균 GC 함량이 60.1%로 나타났다. 또한 173 종의 Lactobacillus 중에서 26종의 genome sequence가 밝혀졌 으며, 평균 GC 함량이 46.61% 나타나 Bifidobacteria가 Lactoba-
cillus 보다 GC 함량이 14.49% 높은 것으로 나타났다[59]. CpGODN은 TLR9과 결합하여 Th1 면역을 유도하고 Th2 면역을 억제하는 기능을 가지고 있으며[72], Clostridium butyricum은 butyrate를 생산하여 Treg 세포 활성화에 의한 allergy 반응을 억제한다[48].
본 논문에서는 allergy 억제에 효과적인 Bifidobacteria를 중 심으로 장내 미생물과 probiotics, CpG ODN synergism,
Clostridium butyricum synergism, butyrate synergism에 대하여 작용기전 중심으로 설명하고자 한다.
본 론
장내 미생물
사람을 포함한 모든 척추동물은 무균(germ-free) 상태로 태 어난다. 출생하는 과정이나 또는 출생 후에 일어나는 사람과 미생물 간의 상호작용은 장 항상성(intestinal homeostasis) 유 지뿐만 아니라 면역 체계의 발달과 기능에 아주 중요한 역할 을 한다(Fig. 1) [50]. 장내 유익, 유해 미생물 간의 불균형 (dysbiosis)이 형성될 경우 장 상피세포의 투과성(permeability) 이 증가하게 되어 염증성 장 질환(inflammatory bowel dis- ease)과 allergy 같은 만성 염증 질환(chronic inflammatory diseases)이 발생한다[64, 65].
장관 면역 시스템은 사람의 신체 중에서 가장 크고 복잡한
면역 기관이다. 무해한 음식 항원에 대하여 효과적으로 항상
성을 유지하지만, 음식 항원에 대한 관용이 허용되지 않을 경
우 음식 allergy를 유발한다[65]. 음식 allergy는 감작기 동안
대부분 Th2 면역반응(Th2 polarized immune response)에 의
해서 유도되어 항원 특이 IgE (allergen-specific IgE)를 생산하
고 항원 노출이 계속 될 경우 설사가 발생하며 심하면 비만세 포 탈과립(mast cell degranulation)으로 과민성 쇼크(anaphy- lactic shock)가 일어난다. 장 상피 세포는 몸으로 들어 온 음식 물과 면역세포 사이에 존재하는 첫 번째 장벽으로 존재하며 면역 반응 조절과 같은 중요한 역할을 한다[4].
유아기 때 allergy 질환이 급증하고 있는 것은 미생물 다양 성(bacterial diversity)과 Lactobacilli나 Bifidobacteria와 같은 유 익균의 감소에 기인한다[1]. Lactobacilli나 Bifidobacteria와 같은 probiotics는 비침입성(noninvasive), 비병원성(nonpathogenic) 이며 그람 양성 박테리아(Gram-positive bacteria)로서 strain- dependent 한 면역 조절 특성(immunomodulatory proper- ties)을 가진다[8]. 또한 probiotics는 병원성 미생물이나 독소 들과 장 상피세포의 부착에 대해 상호 경쟁하고, 장 상피세포 생존(intestinal epithelial cell survival)과 장벽 기능성(barrier function) 증가, 수지상 세포와 같은 면역세포와 직접 상호작 용[85], TLR이나 NOD (nucleotide-binding oligomerization domain) 또는 C-type lectin receptor와 결합 후 cytokine 분비, T-helper cell 반응을 유도한다[42, 75].
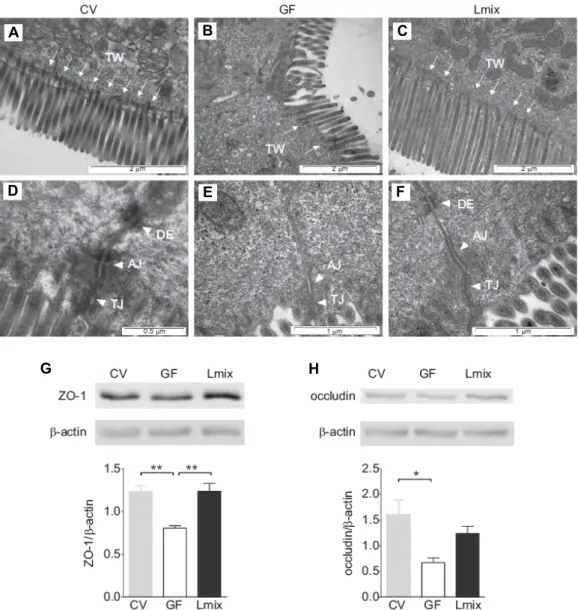
Kozakova 등은 쥐를 대상으로 장벽(intestinal barrier) 발달 에 대한 복합 유산균(2×10
8CFU/ml L. rhamnosus LOCK0 900,
L. rhamnosus LOCK0908, L. casei LOCK0919)의 효능을 조사하였다(Fig. 2) [43]. 일반 환경(conventional conditions) 쥐와 무 균 쥐, 유산균 급여 쥐(무균 + 복합 유산균)를 비교한 결과 일반 환경 쥐의 장벽은 미세융모(brush border)가 규칙적으로 잘 발달되어 있으며, 미세섬유(microfilaments)가 terminal web 방향으로 잘 뻗어 있었다(Fig. 2A). 또한 밀착연접(tight junction)과 부착연접(anchoring junction)에 속하는 adherens junction과 desmosome의 조직화가 잘 이루어져 있었다. 그러 나 무균 쥐의 미세 융모는 불규칙으로 배열되어 있고, 미세섬 유 수가 감소되어 있었으며, 일반 환경 쥐와 유산균 급여 쥐에 비하여 부착 연접 부위(AJ region) 면적이 감소하였다. 무균 쥐 상피세포의 약 30%는 desmosome이 결여된 불완전한 세포 연접 상태를 나타내었으나(Fig. 2B), 유산균 급여 쥐는 일반 환경 쥐의 미세융모 배열이나 미세섬유 상태와 같이 나타났다 (Fig. 2C). 밀착연접 단백질 ZO-1 (Fig. 2G)과 occluding (Fig.
2H)의 발현을 western blot으로 확인한 결과, 무균 쥐에 비해 서 일반 환경 쥐와 유산균 급여 쥐에서 높게 발현되는 것으로 나타났다. 밀착연접은 물질이 새어나가는 것을 막기 위해 세 포를 함께 묶어 판을 형성하는데, 이와 같은 판은 소화관을 따라 이어져 있으며, 소화관 속의 내용물이 주변 조직으로 새 어 들어가는 것을 방지하는 역할을 한다. 부착연접은 세포가 서로 붙어 있게 하거나 세포를 세포외 기질에 붙어 있게 하는 역할을 한다. Adherens junction은 인접 세포의 액틴 미세 필 라멘트(actin cytoskeleton)와 연결되어 있으며, desmosome은 세포 부착단백질(cell surface adhesion proteins)이 세포내 keratin cytoskeletal filaments와 연결되어 있고 미세섬유는 세
포의 모양 유지와 운동에 관여한다[9]. 따라서 장내 미생물이 장 상피 총체성(epithelial integrity)을 견고하게 한다는 것을 알 수 있다.
복합 유산균이 Th1과 Th2 cytokine 생산에 미치는 영향을 조사하기 위하여 자작나무 꽃가루 항원(birch pollen allergen Bet v 1)으로 감작 시킨 쥐로부터 비장(splenocytes)과 장간막 림프절(mesenteric lymph node)을 분리하여 자작나무 꽃가루 항원과 in vitro 공배양 하였다. 그 결과 무균 쥐에 비하여 복합 유산균 급여 쥐에서 비장의 Th2 cytokine IL-4와 IL-5가 감소 하고, Th1 cytokine IFN (interferon)-γ와 TGF-β 수준이 증가 하는 경향을 나타내었으며, 장간막림프절에서도 IL-5가 감소 하고 TGF-β 수준이 증가하는 경향을 나타내어 복합 유산균 급여가 allergy를 억제하는 면역조절 기전(immunoregula- tory mechanisms)을 유도 하였다[43, 73].
장내에 존재하는 박테리아는 수지상 세포나 장 상피세포의 TLR 또는 NOD를 통해 인지되어 장-연관림프 조직(gut-asso- ciated lymphoid tissue; GALT)과 상호작용 한다[10, 84]. 장 상피세포 표면에서 발현되는 TLR은 병원성 미생물을 인식하 며, 염증상태에서 발현이 증가하지만 항상성 상태에서는 TLR ligand에 반응하지 않는다[18]. TLR9이 박테리아의 unmethy- lated DNA와 결합하게 되면 TLR 자극에 대한 관용이 유도되 며, 장 항상성을 유지하는데 기여한다[46]. 설치류에서 땅콩 알러지를 유발하고 TLR9 agonist를 경구 투여 하였을 때 TLR9 활성화를 통해 분비된 장 상피세포 분비 물질들(media- tors)이 수지상 세포의 표현형을 Treg 세포로 분화 시키고 Th1 과 Th17의 발달을 억제하여 allergy 증상을 감소시켰다[31].
Probiotics
2001년 세계보건기구/식량농업기구(World Health Orga-
nization/Food and Agricultural rganization)에서는 pro-
biotics를 살아 있는 미생물로서 사람이나 동물이 충분한 양을
섭취했을 때 유익한 효과를 나타내는 것이라고 정의하였으며,
2002년 FDA는 probiotics로 사용되는 미생물의 안전성에 대
하여 항생제 저항성이나 독소 생산에 대한 가이드라인을 제시
하였다[20, 21]. 장내에서 probiotics는 정상적 미생물 균형이
나 대사 또는 장 면역을 유지하는데 중요한 역할을 한다. 특히
probiotics는 장내에서 대식세포(macrophage)나 수지상 세포
와 같은 항원제시 세포(antigen presenting cell)와 작용하여
특정 cytokine을 분비 시키고 이들 cytokine은 Treg 세포의
기능을 조절하는 immunomodulation 기전을 통하여 면역을
증진시킨다. 이러한 immunomodulation 기전은 다양한 염증
반응이나 gut-organ axises (gut-brain axis, gut-lung axis,
gut-skin axis)로 연결되어 allergy 민감성을 감소시킨다[6, 22,
70]. 장내 미생물은 습진(eczema)과 같은 피부 allergy와 천식
(asthma)과 같은 호흡기계 allergy를 감소시키고[7], gut-brain
axis를 통하여 뇌와 행동에 영향을 주며, 스트레스와 같은 뇌
A
G
B C
D E F
H
Fig. 2. The effects of Lmix colonization on the architecture of the apical junctional complex of enterocytes and the production of ZO-1 and occludin. Electron microscopy micrographs of the apical surfaces of ileal enterocytes in CV, GF and Lmix-colonized mice (Lmix). The epithelial surface is covered by microvilli. Microfilaments extend from the microvilli into the apical cytoplasm and filamentous TW, which was deficient in the GF animals and was restored in the Lmix-colonized mice (A–C). The epithelial cell junctional complex contains the TJs, AJs and DEs. DEs were absent in 30% of the junctional complexes in the GF mice (D–F). Representative micrographs were obtained from 10-15 measurements per sample; n55 samples per group.
Western blot analysis of ZO-1 (G) and occludin (H) in the ileum. A representative mouse from each group is shown (3-4 mice per group were analyzed). Quantification of the signals was performed using ImageJ. The data are expressed as the mean ± SEM of 3-4 mice per group. *p<0.05 and **p<0.01. AJ, adherens junction; DE, desmosome; GF, germ-free; Lmix, Lactobacillus mixture; TJ, tight junction; TW, TW, terminal web; ZO-1, zonulin-1 [43].
기능을 조절 할 수 있다(Fig. 3) [13, 54].
Probiotics로 이용되고 있는 미생물은 Bifidobacteria와 Lacto-
bacillus가 대표적이며, 모유 급여 유아에서 발견되는 대표적인종은 B. bifidum, B. breve, B. longum, B. animalis ssp lactis 이다 [83]. Bifidobacteria는 1899년 파스퇴르 연구소(Pasteur Insti- tute)에 근무하던 소아과 의사이며 노벨상 수상자인 Henry Tissier에 의해 분리되었으며, 두 개의 다리(bifid)를 가진 Y자 모양으로 인해 당시에는 Lactobacillus bifidum 라고 불리었을
만큼 형태나 생리학적 특성이 Lactobacillus와 유사하다[80]. 비 피도 속(genus)에 속하는 종과 아종은 현재 50개 이며, 그 수는
Lactobacillus 173 종에 비교했을 때 상대적으로 적다. 1930년경교토 의대에 근무하는 Minoru Shirota 박사가 Lactobacillus
casei strain Shirota 균주를 배양하였으며, 1935년 일본 YakultHonsha Co., Ltd. 회사가 최초로 probiotics를 상업화 하였다 [86].
탄수화물 구조를 인식하는 lectin 수용체들은 비소화성 올
Fig. 3. Role of probiotics in systemic immunity. Probiotics can improve the generalized immunity by immunomodulation. In im- munomodulation, gut probiotics interact with antigen presenting cells i.e. macrophages and dendritic cells (DC). These APC then affect the T cells population (mainly regulatory cells) to induce systemic immunomodulation. This immunomodulation through gut-brain axis, gut-lung axis and gut-brain-skin/gut-skin axis improves the function of brain, lungs and skin re- spectively, by reducing the susceptibility to various stresses and allergies [77].
리고당을 인식하며, 장 상피 세포에서 발현되는 soluble-type lectin 중의 하나인 galectin은 β-galactoside와 결합하는 특이 성을 가지고 있다[28]. 장 상피 세포에서는 galectin-2, -3, -4, -9가 발현되며 세포질 단백질이지만 분비되어 당단백질로 결 합, 세포 표면에 galectin-glycoprotein lattice를 형성하여 면역 관용을 유도하는 반응을 조절한다[67]. Galectin-9은 Th1, Th17과 수지상 세포에서 발현되는 TIM-3 (T cell im- munoglobulin mucin-3)에 결합하여 Th1과 Th17의 apoptosis 를 유도하고, 수지상 세포를 활성화 시켜 naive CD4
+T 세포에 서 IFN-γ, IL-4와 IL-5를 분비하게 한다. 또한 galectin-9은 IgE 와 복합체를 형성하여 비만세포의 탈분극을 억제함으로써 al- lergy 유발 물질 분비를 억제한다[14].
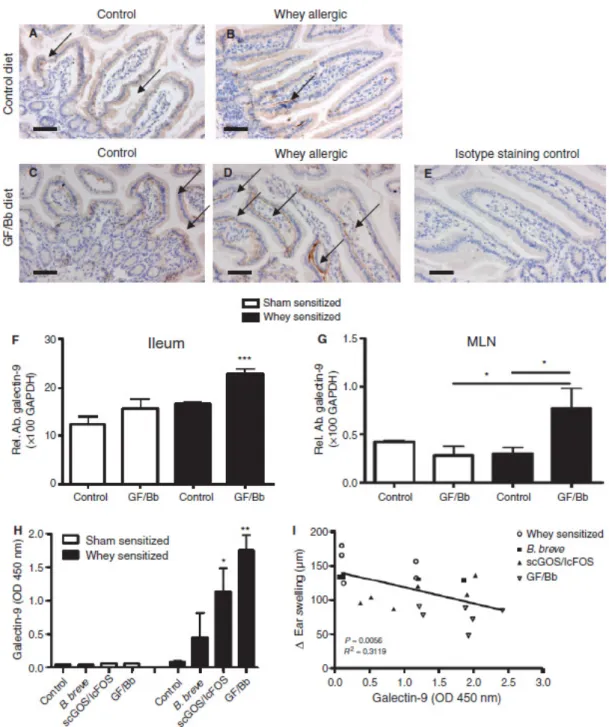
de Kivit 등은 유청(whey)-allergy 유발 생쥐에게 Bifidobac-
terium breve M-16V[2% wt:wt 2×109CFU/g; 1% wt:wt, 9 : 1 short chain galacto oligosaccharides (scGOS)/long chain fructo oligosaccharides (lcFOS)]를 각각 또는 조합한 syn- biotics로 급여하여 galectin-9의 발현을 분석 하였다[15]. 면역 조직학적 염색(immunohistochemical staining) 방법으로 ga- lectin-9 발현을 분석한 결과 galectin-9 발현은 장 상피세포 기저부(basolaterally)에서 발현되며, synbiotics 처리구에서 가장 많이 증가하였다(Fig. 4A - Fig. 4E). 회장(ileum)과 장간 막림프절에서 galectin-9 발현은 synbiotics 처리구에서 증가 하였으며, 특히 회장은 대조구에서도 발현이 증가하여 ga- lectin-9이 면역 세포보다 장 상피세포에서 주로 발현된다는 것을 알 수 있으며(Fig. 4F - Fig. 4H), 혈청 내 galectin-9의 함량은 synbiotics, prebiotics, probiotics 순서로 증가하였다 (Fig. 4H). 혈청 내 galectin-9 발현 함량 증가는 유청-allergy 생쥐에서 급성 과민 반응(acute hypersensitivity response)을 감소시키는 것과 연관된다(Fig. 4I).
Fig. 5는 생쥐에서 synbiotics (GOS, FOS/Bifidibacterium
breve) 처리가 IgE-mediated allergic reaction을 억제하는 설명으로 감작 상태(Fig. 5 중앙 좌측, 소 유청 급여)에서는 allergen 에 의해 Th2-type effector 반응이 증가하여 B 세포에서 IgE 생산이 증가하며, 반복적으로 allergen에 노출될 경우 IgE- primed mast cell degranulation이 일어나 임상적 징후가 발생 하게 된다(blue arrows). 이러한 감작 상태에서 synbiotics 급 여는 장 상피세포에서 galactin-9 분비를 증가시켜 Th1과 Treg 의 분화를 유도한다(I). 그림 중앙 우측은 감작 후 귀에 유청을 주입하는 challenge phase로 synbiotics가 galactin-9의 분비를 촉진시켜 mast cell degranulation을 억제한다(II, Green ar- rows-stimulation, red arrows-suppression). 장 고유층(intes- tinal lamina propria)에 존재하는 수지상 세포가 장 상피세포 에서 유래하는 galectin-9에 노출이 되면 장간막림프절로 향하 여 Th1과 Treg 면역 반응을 일으킨다. 따라서 synbiotics는 IgE 매개 allergy 예방을 위한 효과적이며 안전한 전략이 될 수 있다.
Bifidobacteria
Bifidobacteria는 장내 세균으로 Clostridium, Shigella, E. coil
등의 병원성 세균에 의한 부패성 발효 억제와 Bifidus 균에 의 해 생성된 acetic acid, lactic acid 및 다른 유기산 등이 장관 벽을 자극하여 연동운동과 소화흡수 촉진, 장내 pH 저하 등으 로 각종 유해 세균의 증식을 억제하며 또한 vitamin B1, B2, B12 등을 생성하여 생체에 이용되게 하는 유익한 균이다(Fig.
6) [32, 81].
포유류의 양막은 무균 환경이며, 태아가 산도를 나오기 위 해 양막이 터지는 순간 미생물과의 접촉이 시작된다[36].
Bifidobacteria는 태아의 장내에 가장 먼저 자리 잡는 박테리아
Fig. 4. Galectin-9 is basolaterally expressed by intestinal epithelial cells and increased in serum in whey allergic mice fed GF/Bb to protect against allergy. Immunohistochemical analysis of galectin-9 expression in the proximal ileum (A–E). Diffuse ga- lectin-9 staining was observed in the epithelium of sham- or whey-sensitized mice fed the control diet (A, B) as well as in sham-sensitized mice fed GF/Bb (C). In whey-sensitized mice fed GF/Bb, specific basolateral galectin-9 expression was observed (D). Isotype staining control showed no background (E). In whey-sensitized mice – but not sham-sensitized mice – fed GF/Bb, galectin-9 expression in ileum (F) and mesenteric lymph nodes (MLN) (G) was increased, which was reflected by increased serum galectin-9 levels (H). Increased serum galectin-9 levels correlated with a reduction in acute hypersensitivity response in whey-sensitized mice (I). Data represent n = 5-6 mice per group, mean ± SEM, *p<0.05, **p<0.01[15].
중 하나로서 모유 안에서도 발견되며 산모의 모유를 통해 유 아에게 직접 전달되어 급증하게 된다[12]. Bifidobacteria는 유아 기 장내 세균의 90% 정도를 차지하며, 분유를 먹는 유아에 비해 모유를 먹는 유아에서 더 많이 서식하게 되는데 초기의
높은 비율은 면역과 대사 기능을 조절하는 중요한 역할을 한
다. 이유 후 고형 식이를 시작함에 따라 다양한 종의 박테리아
가 장내에 증가함에 따라 Bifidobacteria의 장내 상대적 비율이
급격히 감소된다[26].
Fig. 5. Serum galectin-9 is enhanced in infants that were effectively treated with GF/Bb. A schematic view of the mechanisms by which GF/Bb suppresses IgE-mediated allergic reactions. During allergic sensitization, a Th2-type effector response is raised against an allergen, which results in IgE production by B cells. Upon repeated exposure to the allergen, IgE-primed mast cells degranulate, resulting in clinical symptoms (blue arrows). Supplementation of GF/Bb to the diet 2 weeks before and during sensitization to whey induces galectin-9 secretion by the intestinal epithelium driving Th1 and Treg cell develop- ment (I). During the challenge phase, galectin-9 induced by GF/Bb directly inhibits mast cell degranulation (II). Green arrows, stimulation; red arrows, suppression [15].
A B
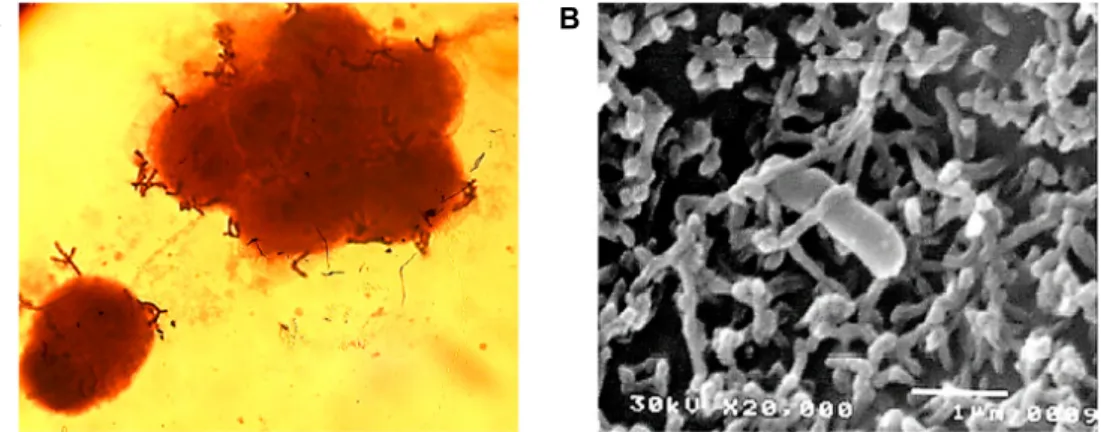
Fig. 6. Adhesion of B. bifidum BGN4 onto the epithelial Caco-2 cell observed by: (A) optical (magnification of 1,000); and (B) scanning electron microscopy (magnification of 20,000, interaction with microvilli of Caco-2 and B. bifidum BGN4). Microbial adherence in (A) was observed after simple staining with crystal violet. Panel (B) was adapted from Kim et al. [39].
장내 미생물 불균형(intestinal microbiota dysbiosis)은 여 러 가지 질병과 연관되며, Bifidobacteria의 불균형(Fig. 7)은 아 토피 질환(atopic disease) [34]과, 과민성 장 증후군(Irritable Bowel Syndrome; IBS) [37], 염증성 장 질환(Inflammatory Bowel Disease; IBD) [57], 대장암(Colorectal carcinoma; CRC) [24], 셀리악 병(celiac disease) [11] 또는 비만(obesity) [11]과 관련되어 있다. 따라서 Bifidobacteria fingerprint는 장내 미생 물 불균형과 장내 미생물 관련 질병을 판단하기 위한 bio- marker로 사용될 수 있다[81].
Bifidobacteria 복합 균주 probiotics (7 종)를 쥐(NC/Nga
mouse)에 급여 했을 때 TSLP와 IgE 발현을 억제 시키고 Flg (Filaggrin)과 FoxP3 (Treg) 발현을 촉진 시켜 아토피를 억제시
켰으며[76], 130명의 산모에게 분만 전 1개월 동안 Bifidobacte-
rium breve M-16V와 Bifidobacterium longum BB53을 급여하고분만 후 유아에게 6개월 동안 급여하여 비급여 유아(36명)와 비교하였다. Bifidobacteria 급여 유아의 아토피가 유의적으로 감소되었으며, 장내 미생물 중 Proteobacteria 또한 유의적으로 감소되었다[17].
Bifidobacteria는 Th1/Th2 balance에 strain-specific 한 면역
자극 특성을 가지며, 이러한 특성에 unmethylated CpG motif
가 일부 기여한다. CpG motif는 TLR9을 활성화 시켜 Th1-
type immune response 유도한다. CpG motif는 진핵생물
(CpG motif 95%가 unmethylated) 보다 원핵생물(CpG motif
95%가 10-30% unmethylated)에 더 많이 존재하며, Bifidobac-
Fig. 7. Bifidobacterial dysbiosis and its relationship with dis- eases: A target for probiotic intervention. IBD: Inflam- matory bowel disease; IBS: Irritable bowel syndrome
[81]. Fig. 8. Immunostimulatory effect of CpG ODNs. CpG ODNs
directly activate human B cells and pDCs, and facilitate the innate and adaptive immune responses [91].
teria가 Lactobacillus 보다 더 많은 GC 함량(60.1% vs 46.61%)와
CpG motif를 가지고 있다(Table 1) [53].
Bifidobacteria가 높은 GC 함량을 가질 경우 unmethylated
CpG motif 빈도수도 높게 나타나는데, Menrd 등이 6개의 균 주(Bifidobacterium longum NCC2705, B. longum DJO10A, B.
longum subsp. infantis ATCC 15697, B. animalis subsp. lactis
AD011, B. adolescentis ATCC 15703)를 대상으로 genome을 조사한 결과 전체 염기 수는 1,933,695 bp~2,832,748 bp 이며, coding sequence는 84~86%, CpG motif는 18,000~27,000개로 나타났다(Table 2) [53]. Bifidobacteria 유래 CpG-rich PCR 산물 을 대식세포(RAW 264.7 macrophage)에 처리했을 때 TNF-α (tumor necrosis factor-α)와 MCP-1 (monocyte chemotactic protein 1)의 발현이 증가 되었다.
CpG ODN (CpG oligodeoxynucleotides) synergism 1984년 Tokunaga 등이 mycobacterium bovis BCG DNA가 type I IFN 생산과 natural killer cell을 증가시켜 항암 효과가 있다는 사실을 밝힌 이래, 현재 CpG ODN이 합성되어 사용되 고 있다[82].
Unmethylated CpG motif ODN은 미생물 유전체에 많이 존재하고 있으며, B 세포와 수지상 세포(pDCs)의 TLR9에 의 해 인식 되어 선천성(innate)과 적응성 면역 반응(adaptive im- mune responses)을 유도한다. CpG ODN은 B 세포를 cell cy- cle G1 phase로 들어가게 하여 IL-6와 IL-10, NF-κB를 분비하 게 하고, 분비된 IL-6는 B 세포의 증가와 활성을 유도하여 항체 생산을 강화시킨다[44]. 또한 CpG ODN에 의해 활성화된 수 지상 세포는 IL-12와 type I IFN, TNF-α를 분비하고, IL-12는 Th0에서 Th1으로 분화를 촉진시켜 IFN-γ를 증가시킨다. 또한 IFN-α는 naive T cell의 CD8-positive cytotoxic T lymphocyte 분화를 촉진시킨다(Fig. 8) [72].
CpG ODN은 exonuclease(DNase)에 의해 빨리 분해되기
때문에 DNase가 작용하는 nucleic acid의 phosphate group의 oxygen을 교체하여 phosphorothioate backbone을 가지는 DNase-resistant CpG ODN이 개발되었다. 그러나 면역반응 감소와 같은 부작용(lymphoid follicle destruction과 organ enlargement)이 나타나[27] phosphodiester backbone을 자연 상태 그대로 유지하는 DNase-resistant natural CpG ODN을 encapsulating 이나 NP (nanoparticle)로 sealing 하는 방법들 이 연구되고 있다[91].
CpG ODN은 크게 4가지 유형 즉, class A, B, C, P로 나누며, class A CpG ODN은 phosphodiester backbone과 중앙 palin- dromic CpG motifs, 3‘과 5’ 말단에 poly (G) sequence가 있으 며, pDC의 TLR9을 활성화 시켜 IFN-α 분비를 유도하나 B cell multiplication은 유도하지 않는다. Class B CpG ODN은 전체 sequence가 phosphorothioate backbone로 구성되어 있으며, B cell activation과 proliferation을 유도하나 pDC로부터 IFN- α 분비 능력이 낮다. Class C CpG ODN은 5‘ 말단에 1-2 개의 CpG motif를 가지고 있으며, 3’ 말단에 phosphorothioate backbone을 가지는 palindromic sequence로 구성되어 B cell proliferation과 pDC IFN-α 분비를 유도하는 Class A와 B의 사이의 능력을 가지고 있다. Class P CpG ODN은 phosphor- othioate backbone과 두 개의 palindromic sequence로 구성되 어 있으며, IFN-α 분비와 nuclear factor-kappa B 활성화 능력 이 높다(Table 3, Fig. 9) [25].
한편, CpG ODN의 분해가 주로 DNase에 의해 일어나기
때문에 plasmid와 같은 ring 구조가 DNase에 저항성이 있을
것이라 착안하여 dumbbell-like structure와 hairpin-loop에
CpG motif를 갖게 한 결과 sequence 양 말단에 phosphor-
othioate backbone을 가지는 linear-structure CpG ODN과 같
은 면역 자극 효과를 가져왔다(Fig. 10A) [71]. 세 개의 sin-
Table 1. General features of sequenced Bifidobacterium and Lactobacillus genomes [87]
Species Genome size (bp) %GC Gene numbers
Bifidobacterium strains B. longum subsp longum NCC2705 B. longum subsp longum DJ010A B. adolescentis ATCC15703 B. animails subsp lactis AD011 B. animalis subsp lactis BI-04 B. animalis subsp lactis DSM 10140 B. bifidum PRL2010
B. bifidum S17 B. dentium Bd1
B. longum subsp infantis 157F B. longum subsp infantis ATCC 15697 B. longum subsp longum BBMN68 B. longum subsp longum JCM 1217 B. longum subsp longum JDM 301 B. breve UCC2003
2,256,640 2,375,792 2,089,645 1,933,695 1,938,709 1,938,483 2,214,656 2,186,822 2,636,367 2,400,312 2,834,748 2,265,943 2,385,164 2,477,838 2,422,684
60%
59%
59%
60%
60%
60%
62%
62%
58%
60%
59%
59%
60%
59%
59%
1798 2061 1701 1603 1631 1629 1791 1845 2197 2062 2588 1878 2009 2035 1642 Lactobacillus strains
L. acidophilus NCFM L. casei ATCC334 L. gasseri ATCC 33323 L. johnsonii NCC533 L. plantarum WCFS1 L. fermentum IFO 3956 L. salivarium UCC118 L. amylovorus GRL 1112 L. brevis ATCC 367 L. casei BL 23 L. casei Zhang L. crispatus ST1
L. delibrueckii subsp bulgaricus ATCC11842 L. delbrueckii subsp bulgaricus ATCC BAA-365 L. delbrueckii subsp bulgaricus ND02
L. helveticus DPC 4571 L. johnsonii FI9785 L. plantarum JDM1 L. plantarum ST-III L. reuteri DSM 20016 L. reuteri JCM 1112 L. rhamnosus GG L. rhamnosus Lc 705 L. sakei 23K
1,993,560 2,895,264 1,894,360 1,992,676 3,308,274 2,098,685 1,827,111 2,067,702 2,291,220 3,079,196 2,861,848 2,043,161 1,864,998 1,856,951 2,125,753 2,080,931 1,755,993 3,197,759 3,254,376 1,999,618 2,039,414 3,010,111 2,968,598 1,884,661
34%
46%
35%
34%
44%
51%
32%
38%
46%
46%
46%
36%
49%
49%
49%
37%
34%
44%
44%
38%
38%
46%
46%
41%
1938 2909 1898 1918 3135 1912 1864 2126 2314 3090 2906 2100 2184 2033 2177 1838 1780 3029 3137 2027 1901 2985 2954 1963
gle-strand DNA로 만들어진 Y-DNA (Y-shaped DNA) 구조는 TNF-α와 IL-6의 분비를 유도하였으나 DNase에 쉽게 분해되 었으며[61, 62], Y-DNA 연결에 의한 DL-DNA (dendrimer- like DNA) 구조는 DNase에 대한 저항성과 면역자극 효과가 나타났다(Fig. 10B) [68]. 네 개의 55-mer ODN으로 구성된 CpG-bearing DNA tetrahedral nanostructure는 DNase 대한 저항성을 나타내고 macrophage-like cell에 효과적으로 결합 하여 TLR9 활성화를 통한 TNF-α와 IL-6, IL-12과 같은 다양한
pro-inflammatory cytokine의 분비를 유도하였다(Fig. 10C) [47].
HT-29 monolayer에 CpG-ODN과 nondigestible oligo- saccharide (scGOS/lcFOS)를 함께 처리했을 때 galectin-9의 발현이 증가하고 Treg와 Th1 polarization이 유도되었다[15].
쥐에게 바퀴벌레 allergy 유발 물질(cockroach allergen)을 코
(intra-nasally)를 통하여 자극하고 CpG-ODN을 처리했을 때
eosinophilic inflammation, goblet cell hyperplasia, airway
Table 2. In silico search of immunostimulatory motifs on sense and antisense strands of Bifidobacterium genomesa [053]
CpG motif Sequence
B. longumNCC2705
B. longum
DJO10A
B. longum
subsp. infantis
ATCC 15697
B. adolescentis
ATCC 15703
B. animalis
subsp. lactis
AD011
Type 1
bAACGTT
AACGTC AACGCT AACGCC AGCGTT AGCGTC AGCGCT AGCGCC GACGTT
cGACGTC GACGCT GACGCC GGCGTT GGCGTC GGCGCT GGCGCC
296 541 424 1,203 485 629 469 995 528 648 527 1,516 1,186 1,538 975 1,014
313 611 414 1,253 492 661 486 1,054 571 668 660 1,651 1,300 1,661 1,070 1,054
375 698 560 1,546 501 817 485 1,174 729 865 859 2,097 1,540 2,046 1,179 1,490
433 703 589 1,191 476 705 370 928 730 844 744 1,371 1,144 1,460 933 1,112
151 386 285 894 320 572 361 838 385 778 541 1,294 894 1,353 921 1,433
Type 1 total 12,974 13,919 16,961 13,733 11,406
Type 2 ATCGTT
ATCGTC ATCGCT ATCGCC GTCGTT
dGTCGTC GTCGCT GTCGCC
553 1,250 501 2,079 663 960 526 1,562
(596) (1,236) (435) (2,044) (632) (931) (508) (1,496)
564 1,312 516 2,171 693 1,076 576 1,638
(640) (1,312) (455) (2,196) (692) (999) (514) (1,597)
666 1,591 545 2,460 898 1,275 570 1,949
(718) (1,600) (508) (2,513) (834) (1,286) (628) (1,964)
573 1,408 425 1,951 661 1,011 537 1,338
(593) (1,406) (434) (1,863) (681) (988) (550) (1,290)
405 1,204 350 1,642 672 1,100 536 1,384
(341) (1,152) (295) (1,715) (594) (1,138) (507) (181)
Type 2 total 8,094(7,878) 8,546(8,405) 9,954(10,051) 7,904(7,805) 7,293(7,123)
Total 21,068 22,465 26,915 21,637 18,699
Published motifs
eBL 07 OL-LB7 ID35
Active motif of ID35 AT ODN
GCGTCGGTTTCGGTGCTCAC CGGCACGCTCACGATTCTTG ACTTCGTTTTCTGCGTCAA TTTCHTTT
ATTTTTAC
1 0 0 16 6
(0) (0) (0) (22) (4)
1 0 0 20 8
(0) (0) (0) (21) (3)
1 0 0 26 8
(0) (0) (0) (28) (13)
0 0 0 42 11
(0) (0) (0) (46) (9)
0 0 0 13 2
(0) (0) (0) (13) (1)
a
Values indicate the number of the times that each sequence appears in the respective genome. The values in parentheses indicate the results for the antisense strand.
bPalindromic sequences, found equally on antisense strand.
cSequence for optimal im- munostimulatory motif of murine cells.
dSequence for optimal immunostimulatory motif of human cells.
eImmunostimulatory motifs described in literature on lactic acid bacteria, including Bifidobacterium BL07 (79), Lactobacillus OL-LB7 (40), ID35 (30), and AT ODN (74).
hyper-responsiveness, IL-13, IL-5, serum IgE, cockroach spe- cific IgE, IgG1/IgG2a ratio를 감소시켰는데 이러한 억제 기능 은 폐에서 CD4
+Foxp3
+regulatory T 세포와 IL-10 up-regu- lation에 기인하였다[38]. 쥐의 기관지에 house dust mite-in- duced allergy를 유발 한 후 NP-conjugated CpG를 처리하고 폐 면역을 조사하였을 때 free CpG 처리에 비하여 NP-CpG 처리가 수지상 세포 활성 및 Th1 면역을 증가시켰으며, 예방 효과로서 NP-CpG 처리 후 house dust mite-induced allergy 를 유발하였을 때 eosinophilia, IgE level, mucus production, Th2 cytokine이 유의적으로 감소되었다[5].
Allergy 면역질환의 효과적인 치료를 위하여 CpG motif ODN과 probiotics, prebiotics를 혼합하는 방법 또는 CpG mo-
tif ODN의 경구용 투여와 기관지 계통 분무 투여에 대한 다양 한 방법이 고려 될 수 있다.
Clostridium butyricum synergism
사람의 대장 내에는 Firmicutes와 Bacteroidetes, Proteobacteria,
Actinobacteria가 98% 이상을 차지하며, 여기에 500-1,000 spe-cies와 약 7,000 strains이 존재하는데 그 중 약 80%는 배양할
수 없는 균이다. 장내에서 Bifidobacteria가 fructose, oligo-
fructose, inulin 등과 같은 저항성 탄수화물을 이용하여 중간
대사 물질 lactate나 acetate로 생산하고, Clostridium butyricum
을 포함하는 Clostridia 계통이나 Roseburia, Faecalibacterium,
Butyrivibrio 같은 colon butyrate producer가 기능성 대사물질Table 3. Features of each class of cytosine-phosphate-guanosine oligodeoxynucleotides (CpG ODNs) [25]
Class A (type D) Class B (type K) Class C Class P
ODN structure
Central phosphodiester region containing one or more CpG motifs in a palindrome and 5’ and/or 3’
ends consisting of poly(G) motifs with phosphorothioate backbone
Completely phosphorothioate backbone
One or more 5’ CpG motif(s) and a 3’ palindrome
Two palindromes consisting of phosphorothioate backbone
Examples ODN2216 (for human) ODN2336 (for human) ODN1585 (for mouse)
ODN2006 (also know as PF-3512676 and CpG7909, for human) ODN1668 (for mouse) ODN1826 (for mouse)
ODN2395 (for human and mouse) ODN M362 (for human and mouse)
ODN21798
Mainly stimulated cell types
pDCs B cells pDCs and B cells pDCs
Actions Innate immune responses:
IFN-α, TNF-α, and IL-12 secretion
Innate immune responses:
IL-6, IL-10, and IL-12 secretion
Intermediate between
the A and B classes
Potency for IFN-α secretion is higher than that of CpG ODN in class C
Adaptive immune responses:
IL-12 and IP10 secretion
Adaptive immune responses:
antibody production:
IL-6 and IL-12 secretion
Abbreviations: IFNα, interferon-alpha; IL, interleukin; IP10, interferon-gamma-inducible protein of 10 kDa; pDC, plasmacytoid denddritic cell; TNF-α, tumor necrosis factor-alpha.
Fig. 9. Features of cytosine-phosphate-guanosine oligodeox- ynucleotide (ODN) sequences in each class. Note:
Underlining indicates palindromic sequence; black and red hyphens indicate phosphodiester and phosphor- othioate bonds, respectively [25].
butyrate를 생산한다[19].
Clostridium butyricum은 막대 모양의 혐기성 그람양성 포자
형성 균이며 사람과 동물의 장에서 발견되는 미생물로 대장 내에서 Bifidobacteria에 의해 생산된 lactate나 sugar, succinate 를 이용하여 탄소수가 2-5개로 된 단쇄지방산을 생산한다 (Table 4) [16].
2002년 FDA는 probiotics로 사용되는 미생물에 대하여 항 생제 저항성이나 독소 생산에 대한 가이드라인을 제시하였다 [21]. Clostridium butyricum MIYARI 588 (CBM 588)은 1960년 대 일본 나고야 토양에서 분리한 미생물로 EU를 포함하여 많 은 나라에서 건강기능성 식품이나 동물사료 첨가제로 사용되 고 있다. 이 미생물은 α, β, ε toxin 유전자나 botulin neuro- toxins type A, B, E, F가 존재하지 않으며 사람과 동물에서 중요하게 사용되는 모든 항생제에 대한 민감성을 나타내어 probiotics 균주로 적합한 것으로 나타났다[33].
쥐에서 장염을 유발하고 Clostridium butyricum MIYAIRI 588을 처리했을 때 TLR2나 MyD88 (myeloid differentiation primary response gene 88) signaling pathway를 통하여 in- testinal macrophage를 직접 자극하고 IL-10의 생산을 유도 하 였으며, human lamina propria 유래 CD14
+⁻CD33
+macro- phage에 Enterococcus faecalis를 처리하면 proinflammatory cytokine (IL-12/IL-23p40, IL-23, TNF-a, IL-6)을 분비 하지만,
Clostridium butyricum MIYAIRI 588을 처리하면 human IBDtissue 유래 CD14
+CD33
+lamina propria macrophage는 높은 수준의 IL-10 분비를 유도하였다(Fig. 11) [35].
Clostridium butyricum과 Bifidobacterium infantis probiotic
mixture를 장기간 급여 시 AAD (antibiotic-associated diar-
rhea)로 파괴된 장내 미생물 균총과 IL-10, IFN-γ, TNF-α가
정상으로 회복되었으며[49], Clostridium butyricum은 in-
Table 4. Major metabolic features of Clostridium [16]
Fermentation pathway Substrate(s) Products Representative species
Homoacetate (Wood-Ljungdahl) Propionate Acrylyl-CoA
Succinate decarboxylation Butyrate
Acetone/butanol
Alcoholotrophic
Proteolytic, peptolytic, use of amino acids Stickland reaction Purinolytic Pyrimidinolytic
Sugars (e.g., fructose), CO
2+H
2, CO
Lactate Succinate
Sugars (e.g., glucose) Sugars (e.g., glucose)
Various alcohols, alcohol + acid Proteins, peptides, amino acids Pairs of amino acids Purines
Pyrimidines
Acetate (under special conditions also ethanol)
Propionate, acetate, CO
2Propionate, CO
2Butyrate, acetate, CO
2, H
2Butanol, acetone,
CO
2, H
2, butyrate, acetate Various acids and solvents Various acids
Acetate, propionate, butyrate, CO
2, NH
4+Acetate, CO
2, NH
4+Acetate, formate, CO
2, NH
4+Acids, CO
2, NH
4+C. aceticum. C. ljungdahlii
C. propionicum C. mayombei C. butyricum C. acetobutylicum
C. aceticum C. kluyveri C. propionicum, C. tetanomorphum C. sticklandii C. purinilyticum C. oroticum
Fig. 10. Structures of cytosine-phosphate-guanosine (CpG) oligo- deoxynucleotides (ODNs) consisting of entirely phospho- diester backbone. (A) Sequence and structure of CpG ODN with a dumbbell-like structure: the CpG ODN with a dumbbell llike structure has 30 nucleotides in both loops that contain three CpG dinucleotide motifs.a (B) Structure of dendrimer-like DNA: Y-shaped DNA consists of three single-stranded DNA with 30 nucleotides containing CpG dinucleotide motifs; G1, G2, and G3 dendrimer-like struc- tures were synthesized by ligation of Y-shaped DNA; the sizes of CpG ODNs with G1, G2, and G3 dendrimer-like structures were about 12, 20, and 36 nm, respectively. (C) Assembly of CpG bearing DNA tetrahedral nanostructure:
the core tetrahedral nanostructure consists of assembly with four 55-mer ODNs. The CpG motif is linked to each ODN via a 7-mer oligothyamine spacer [25].
testinal allergic inflammation에서 SIT (specific immunoth- erapy) 효과를 강화시켰다[75]. 특히 천식환자에서 Der p (dermatophagoides pteronyssinus) 1 specific B를 regulatory B로 전환시켰다[48]. 건강한 사람의 장내에서 분리한 17 개의
Clostridia strain 급여는 Treg 세포의 수와 기능을 증가시켜allergy-설사와 장염의 증상을 완화 시키는 결과를 나타냈으 며, C lostridia가 생산하는 단쇄지방산 중 butyrate는 수지상 세포의 anti-inflammatory 유전자의 발현을 유도하기 위해 GPR109a signaling pathway를 활성화시켰고, 또는 GPR43 활 성화를 통하여 tTreg 세포 proliferation을 직접 자극하거나 HADC 활성을 억제시켜 Foxp3 gene intronic enhancer의 his- tone H3 acetylation을 통해 naive CD4⁺ T 세포를 pTreg 세포 로 분화시켰다[3, 58].
따라서 장내에서 bacterial antigen이나 butyrate를 생산하
고 TGF-β 환경을 제공하여 Treg 세포의 분화와 확장을 돕는
Clostridium butyricum이 allergy 증상 억제에 효과가 있는 것으로 나타났으며, lactate와 acetate를 생산하는 Bifidobacteria와 함께 probiotics로 이용할 경우 synergism이 나타났다.
Butyrate synergism
Butyrate는 장 건강에 중요한 역할을 하고 있으며, 특히 장 속에 항염증 환경을 만드는데 필수적인 요소로 알려져 있다.
Butyrate는 주로 대장에서 Clostridium butyricum 등의 미생물 에 의해 생성되어 세포내로 흡수되며, TCA cycle을 거쳐 ATP 생성에 이용될 뿐만 아니라 장내 항염 효과 및 T 세포 분화와 증식을 조절하는 면역조절 기능이 있는 것으로 알려져 있다.
또한 장 상피 세포막을 보호하는 역할까지 하고 있는 것으로
알려져 궤양성 대장염(ulcerative colitis; UC), 염증성 장 질환
A B
Fig. 11. (A) Dextran sodoim sulfate treatment of mice allowed bacteria to invade the colon mucus layer and penetrate the epithelial barrier into the lamina propria. In the inflamed condition, Clostridium butyricum MIYAIRI 588 directly stimulated intestinal macrophages through the Toll-like receptor 2 (TLR2)/myeloid differentiation primary response gene 88 (MyD88) signaling pathway and induced interleukin (IL)-10 production by macrophages in mice. (B) CD14
+⁻CD33
+macrophages isolated from human lamina propria stimulated with Enterococcus faecalis induced proinflammatory cytokines, such as IL-12/IL-23p40, IL-23, tumor necrosis factor a (TNF-a), and IL-6. However, Clostridium butyricum MIYAIRI 588 stimulation induced higher levels of IL-10 in CD14
+CD33
+lamina propria macrophages from human inflammatory bowel disease (IBD) tissue [35].
Fig. 12. Dietary fiber effect involves reshaping of the gut micro- biota as well as increased levels of short-chain fatty acids and activity of their receptors GPR43 and GPR109a. High-fiber feeding also increased tolerogenic CD103
+DCs activity, leading to increased Treg cell dif- ferentiation [78].
(inflammatory bowel disease; IBD), 크론병(Crohn’s disease), 대장암(colorectal cancer), 설사(diarrhea) 등의 질병의 치료에 효과적이라는 연구결과가 지속적으로 보고되고 있다[69].
높은 수준의 섬유소 급여(high-fiber feeding)는 장내 미생 물 환경(gut microbial ecology)을 변화시켜 butyrate와 같은 단쇄지방산을 생산하고 CD103⁺ DC의 retinal dehydrogenase
activity를 증가 시켜 음식 allergy를 억제한다(Fig. 12) [78].
6개월령 유아에서 조사한 결과 장내 butyrate 생산 미생물 (Coprococcus eutactus)과 미생물 다양성이 증가할수록 아토피 증상이 완화 되었다[63]. 최근 allergy 비염 마우스 모델에서 Na-butyrate가 TSLP 발현을 억제함과 동시에 Th1/Th2 bal- ance를 유지하는 효과를 통해 allergy-비염증세를 완화시킨다 고 보고되었다[88].
따라서 Bifidobacteria와 Clostridium butyricum의 synergism 에 의해서 생성된 기능성 대사체 butyrate는 probiotics 또는 prebiotics와 함께 이용될 경우 allergy 억제에 대한 더 효과적 인 synergism 발생 한다.
결 론
Allergy는 중요한 만성질환으로서 유아에서 가장 많이 발 생하고 최근에는 청년기나 성인에서도 일반적으로 발생하고 있어 개발도상국가에서는 공중 보건의 부담(public health burden)이 되고 있다. Allergy와 같은 면역 관련 질병과 미생 물과의 연관성을 규명하는 과학적 연구는 미생물 분류(catalo- gue)나 미생물 집단 불균형(dysbiosis)을 분석하는 기술 개발 능력과 함께 발전해 왔다.
장내 미생물은 점막 항상성(mucosal homeostasis)에 중요 한 상피세포 총체성과 Treg 세포를 증가 시키며, 특히 GOS/
FOS와 같은 저항성 탄수화물 prebiotics와 Bifidobacteria와 같
은 probiotics는 장내 미생물 집단 불균형을 정상으로 회복시
키고, galactin-9 발현 증가, Th1과 Treg 분화 증가를 통하여
allergy 증상을 감소시키거나 예방한다.
Bifidobacteria는 Lactobacillus 보다 많은 GC 함량(60.1% vs
46.61%)과 CpG motif를 가지고 있는데, 이러한 CpG ODN은 TLR9에 의해 인식 되어 선천성 면역과 적응성 면역 반응을 유도한다. CpG ODN은 B 세포의 증가와 활성을 유도하여 항 체 생산을 강화시키고, 수지상 세포 활성화와 Th1 세포 분화 촉진, CD8-positive cytotoxic T lymphocyte 분화를 촉진시키 는 면역조절자(immunoregulator) 기능을 가진다. Clostridium genus는 이질적(heterogeneous)인 특성을 가지는 박테리아로 서 사람과 동물에게 병원성인 미생물(C. botulinum, C. difficile,
C. tetani, C. perfringens)이 있는 반면, 기능성 대사체 butyrate를 생산하여 Treg 세포 수나 기능을 증가 시키는 유익한 미생 물(IV, XIVa, XVIII species; Clostridium clostridioforme, Clostri-
dium innocuum, Clostridium ramosum)이 있다.따라서 Bifidobacteria가 중간대사 물질로 acetate와 lactate를 생산하고, 이러한 중간 대사물질을 이용하여 생체 내 기능성 물질 butyrate를 생산하는 Clostridium butyricum을 probiotics 로 사용하거나 CpG ODN이나 butyrate를 조합하여 사용하면 장내 면역조절을 위한 synergy 효과를 나타낼 수 있다.
감사의 글
이 논문은 2015년도 경남과학기술대학교 대학회계 연구비 지원에 의하여 연구되었음.
References



![Table 1. General features of sequenced Bifidobacterium and Lactobacillus genomes [87]](https://thumb-ap.123doks.com/thumbv2/123dokinfo/5005268.548561/9.892.82.806.182.939/table-general-features-sequenced-bifidobacterium-lactobacillus-genomes.webp)
![Table 2. In silico search of immunostimulatory motifs on sense and antisense strands of Bifidobacterium genomesa [053]](https://thumb-ap.123doks.com/thumbv2/123dokinfo/5005268.548561/10.892.82.807.184.775/table-silico-search-immunostimulatory-antisense-strands-bifidobacterium-genomesa.webp)
![Table 3. Features of each class of cytosine-phosphate-guanosine oligodeoxynucleotides (CpG ODNs) [25]](https://thumb-ap.123doks.com/thumbv2/123dokinfo/5005268.548561/11.892.97.807.180.600/table-features-class-cytosine-phosphate-guanosine-oligodeoxynucleotides-odns.webp)
![Table 4. Major metabolic features of Clostridium [16]](https://thumb-ap.123doks.com/thumbv2/123dokinfo/5005268.548561/12.892.85.805.181.486/table-major-metabolic-features-clostridium.webp)
