미강의 섭취가 한계유영운동에 미치는 영향
김신태1*․박정진1,2*․김경미3․전우진1,2
1전남대학교 식품영양학과
2전남대학교 생활과학연구소
3한국폴리텍대학 바이오캠퍼스 바이오식품분석학과
Effect of Rice Bran Supplementation on Endurance Exercise Capacity in Mice
Shintae Kim
1*, Jeongjin Park
1,2*, Kyungmi Kim
3, and Woojin Jun
1,21
Division of Food and Nutrition and
2Research Institute for Human Ecology, Chonnam National University
3
Department of Biofood Analysis, Korea Bio Polytechnic
ABSTRACT This study examined the effects of the endurance exercise capacity of rice bran supplementation in mice.
For the endurance exercise capacity, mice were divided into three groups: sedentary control (CON), exercise control (Ex-CON), and exercise and rice bran supplementation (Ex-RB). Ex-RB showed a significantly prolonged swimming time compared to Ex-CON. For the Ex-RB group, the blood lactate level was lower than that of the Ex-CON group, and the blood non-esterified fatty acids level was higher. The Ex-RB group increased the glycogen level and antioxidant enzyme activities significantly compared to Ex-CON. These results suggest that rice bran can increase the endurance exercise capacity by delaying the accumulation of lactate, saving glycogen, and elevating the antioxidant defense activities.
Key words: endurance exercise, swimming, antioxidant activities, rice bran
Received 28 August 2019; Accepted 21 November 2019 Corresponding author: Woojin Jun, Division of Food and Nutrition, Chonnam National University, Gwangju 61186, Korea
E-mail: [email protected], Phone: +82-62-530-1337
*These authors contributed equally to this work.
Author information: Shintae Kim (Graduate student), Jeongjin Park (Instructor), Kyungmi Kim (Professor), Woojin Jun (Professor)
서 론
현대사회의 급격한 경제성장과 과학의 발달로 인간의 수 명이 증가하고 있는 것은 전 세계적으로 공통된 현상이다.
그러나 이러한 인간 수명의 연장과 복잡한 사회구조에 따른 스트레스의 증가, 식생활의 변화 등으로 인류 건강은 위협받 고 있다(Kim 등, 2013). 이로 인해 단순한 식사보다는 인체 에 유용한 기능을 가진 원료와 성분을 사용하여 제조한 건강 기능식품에 대한 관심이 지속해서 증가하고 있으며, 질병 예방에도 도움을 주는 규칙적인 운동은 건강에 유익한 효과 를 주므로 삶의 질 향상을 위해 권장되고 있다(Cha와 Kim, 2008).
운동수행능력이란 근육이나 근조직이 단 한 번에 발휘할 수 있는 최대의 힘을 이용하여 운동을 수행하는 능력을 말한 다. 운동 부족으로 인하여 활동량이 적어지게 되면 근육이 약화하므로 운동수행능력이 저하될 수 있으며, 이로 인해 체
력 저하 및 비만, 고지혈증, 고혈압 등의 문제를 일으킬 수 있다(Fisher-Wellman 등, 2009). 운동수행능력을 향상하 려면 규칙적인 운동을 하고 근육의 에너지원으로 쓰이는 단 백질, 지방, 탄수화물 등의 영양소를 적절하게 섭취하는 것 이 중요하다.
운동시간을 향상시키기 위해서는 여러 가지 메카니즘이 복합적으로 작용한다(Lamou 등, 2016). 우선 에너지의 고 갈을 막기 위해 혈중에 있는 글루코오스를 사용하는 대신 많은 에너지를 생산할 수 있는 지방을 에너지원으로 사용해 야 운동시간이 증가하게 된다(Kim 등, 2015). 이러한 이유 는 글루코오스의 소모가 증가할수록 에너지 고갈이 빨리 오 고 혈액을 산성화시켜 근육에 축적되어 근육을 뭉치게 만들 고 피로물질을 생성하는 젖산의 축적으로 인해 운동시간이 짧아지게 되므로 글루코오스보다는 지방을 소모하여 에너 지원으로 사용해야 한다(Durkot 등, 1995). 또한 운동을 하 게 되면 세포 내 에너지 생성 기관인 미토콘드리아가 활발하 게 작동이 된다. 미토콘드리아는 에너지 생성을 위해 해당과 정과 TCA 회로를 통해 생성된 NADH와 FADH2를 전자전 달계로 이동을 시키게 되고 전자전달계를 거치면서 많은 ATP가 생성되며, 활성산소를 생성하게 된다(Flis 등, 2018).
이렇게 생성된 활성산소는 체내에 산화효소인 superoxide dismutase(SOD)에 의해 독성이 덜 한 H2O2가 되며, 생성된
H2O2는 catalase(CAT)나 glutathione peroxidase(GPx) 에 의해 물과 산소로 된다(Powers와 Jackson, 2008). 운동 을 하게 되면 많은 에너지가 필요하여 근육에서 미토콘드리 아 활성이 높아지게 되며 끊임없이 활성산소가 생성된다.
이로 인해 활성산소와 산화효소 사이에 불균형이 일어나게 되고 활성산소가 많이 증가하게 된다. 증가한 활성산소는 체내에 독소로 작용하여 세포에 손상을 입히고 이로 인해 운동시간이 짧아지게 된다(Finsterer, 2012). 운동수행능력 을 향상하기 위해서는 반복적인 운동도 중요하지만 적절한 식품 섭취 또한 중요하다(You 등, 2015a). 운동 시 적절한 식품 섭취는 에너지원으로 사용되거나 젖산, 이산화탄소와 같은 대사산물 생성을 억제하여 몸을 회복하는 데 중요한 역할을 한다.
쌀은 왕겨층과 과피, 종피 및 호분층으로 되어 있는 쌀겨 층과 배유와 배아로 구성되어 있다. 미강은 현미에서 백미를 만들 때 떨어져 나오는 부분을 가리킨다. 미강은 단백질, 식 이섬유, 각종 비타민, 미네랄뿐만 아니라 생리활성 물질인 오리자놀, 토코페롤, 레시틴, 스테롤 등을 함유하고 있다 (Nagendra Prasad 등, 2011). 이러한 생리활성물질은 항산 화 능력을 가지고 있어 운동 시 생성되는 산화적 스트레스를 완화하여 운동수행능력을 향상시키는 것으로 알려져 있다 (Howes와 Simmonds, 2014). 또한 쌀 단백질은 근손실을 막고 근육량을 증가시키는 데 중요한 영양분으로 작용하여 운동수행능력 향상에 좋은 소재로 알려져 있다(Joy 등, 2013).
이러한 유용한 소재임에도 불구하고 생산되는 미강의 30%
정도만 미강유 제조에 사용되고 나머지는 사료나 비료 등에 이용되거나 폐기물화되고 있는 실정이다(Seon 등, 2016).
이에 본 연구에서는 미강이 마우스의 한계유영운동에 미 치는 영향을 확인하고자 하였다. 또한 한계유영운동 상태 지표인 젖산(lactic acid), 유리지방산(non-esterified fatty acids, NEFA), 글리코겐 및 항산화 능력을 측정하여 운동수 행능력 개선 기능성 식품의 소재 및 제품 개발의 기초자료로 써 활용하고자 본 연구를 수행하였다.
재료 및 방법
실험소재
본 연구에 사용된 미강은 (주)아이앤비(Yesan, Korea)의 파우더 형태의 제품을 농업회사법인(주)산들촌(Damyang, Korea)에서 구매한 것을 제공받아 사용하였다. 전체 1 kg 미강을 약 50 g씩 소분하여 -20°C에 보관 후 실험에 사용하 였다.
실험동물 및 사육조건
본 실험에 사용된 4주령 ICR 수컷 마우스(18~20 g)를 Orient Bio(Gwangju, Korea)에서 구입하여 조명시간은 오 전 8시부터 오후 8시까지 12시간으로 설정, 실내온도 23±
3°C, 습도는 50%로 조절한 동물 사육실에서 사육하였다.
일주일간 사육 환경에 적응시킨 후 실험에 사용하였다. 실험 기간 동안 5L79(Orient Bio, Inc., Seongnam, Korea) 일반 식이와 수돗물을 급수로 제공하였다. 실험 기간 중 사료와 급수는 제한하지 않았다. 본 연구의 동물실험은 전남대학교 동물실험윤리위원회의 승인 하에 수행되었다(CNU IACUC- YB-R-2017-03).
실험소재 투여 및 유영운동 실험
실험동물은 실험 시작 전 체중을 동일하게 하여 비운동군 (CON)과 한계유영운동을 실시한 운동군(Ex-CON), 한계유 영운동 실시 및 미강을 투여한 군(Ex-RB) 등 총 3군으로 나누었다. CON군과 Ex-CON군은 증류수를, Ex-RB군은 미강 분말을 증류수에 녹여 1 g/kg b.w./d로 경구투여 하였 으며 각 그룹당 8마리씩 21일간 실험을 진행하였다. 마우스 의 행동과 외형은 매일 관찰하였고 한계운동은 7일 간격으 로 진행하였으며, 운동 시 3시간 전 식이를 제거하였고 2시 간 전 경구투여를 시행하였다(You 등, 2015a). 한계운동수 행능력 평가를 위해 유영운동을 진행하였다. 유영운동은 투 명하게 제작한 아크릴 수조(90×45×45 cm)에 35 cm 높이 까지 물을 채운 후 수온을 34°C로 유지하여 펌프와 유량계 (type F45500, Blue White Co., Westminster, CA, USA) 를 이용해 물을 순환시키고, 8 L/min의 유속을 유지하며 실 험을 진행하였다(Matsumoto 등, 1996). 한계운동시간은 마우스가 물속에 빠져 7초가 경과할 때까지 물 표면으로 올 라오지 못하는 때를 최대 유영운동시간으로 하였다.
혈액 내 젖산 및 유리지방산 측정
혈중 젖산과 유리지방산은 운동 전과 후에 미정맥에서 혈 액을 채취하여 측정하였다. 혈중 젖산은 상용화된 테스트 스트립(Arkray, Tokyo, Japan)을 사용했으며, 유리지방산 은 혈액을 채취하여 3,000 rpm에서 5분간 원심분리 후 상층 액인 혈청만 채취하여 상용화된 키트(Arkray)를 이용해 측 정하였다.
근육 조직 글리코겐 함량 측정
글리코겐 함량은 마지막 한계운동 후 4시간이 지나고 마 우스를 해부하여 얻은 근육 조직을 이용해 측정하였다. 막자 사발에 액체질소를 이용해 균질화한 후 anthrone 법을 이용 해 측정하였다(Gierus와 Rocha, 1997). 근육 조직 100 mg 에 30% KOH 1 mL를 첨가하여 100°C에서 30분 가열한 후 실온에서 10분간 방치하고 증류수 0.4 mL 첨가한 다음 0.2% anthrone 용액 2 mL를 첨가하여 실온에서 15분간 반응시킨 후 620 nm에서 흡광도를 측정하고 표준곡선 계산 식에 대입하여 mg glycogen/g tissue로 표기하였다. 사용 된 표준곡선은 bovine liver glycogen을 이용해 작성하였 다.
0 10 20 30 40 50 60 70
0 days 7 days 14 days 21 days
Swimming time (min) .
Ex-CON
Ex-RB *
Fig. 1. Effect of rice bran on exhaustive swimming capacity in
mice. The Ex-CON and Ex-RB did swimming exercise until they were exhausted. The Ex-RB was orally given rice bran at a dose of 1 g/kg b.w./d for 21 days. Data are expressed the mean±SE.The asterisk above the bar is statically different from the Ex- CON group by Student’s t-test (*
P<0.05).
0 0.5 1 1.5 2 2.5 3 3.5 4 4.5 5
Ex-CON Ex-RB
Blood lactate level (mM/L) .
Pre-swimming Post-swimming
*
Fig. 2. Effect of rice bran on blood lactate level. The Ex-CON
and Ex-RB of lactate levels was evaluated before and 10 min after forced swimming with a current flow of 8 L/min. The Ex-RB was orally given rice bran at a dose of 1 g/kg b.w./d for 21 days. Data are expressed the mean±SE. The asterisk above the bar is statistically different from Ex-CON group by Student’st-test (
*P<0.05).
근육 조직 항산화 효소 활성 평가
조직 희석 및 단백질 정량: 마지막 한계운동 후 4시간이 경과했을 때 마우스를 해부하여 채취한 근육 조직에 액체질 소를 첨가하여 막자와 막자사발을 이용해 잘게 분쇄하였다.
분쇄한 근육 조직에 용해액(1×KCl)을 첨가하여 아이스 위 에서 2시간 반응시키고, 용해된 조직을 13,000 rpm에서 30 분간 원심분리 하였다. 원심분리 후 투명한 상층액을 1×
PBS로 10배 희석하여 단백질 정량 용액(bradford)을 첨가 하여 15분간 반응시킨 후 595 nm에서 흡광도를 측정하고 계산식에 대입하여 mg/mL로 나타내었다.
항산화 효소 활성 평가: 과산화수소를 물과 산소로 변화 시키는 CAT 활성은 spectrophotometric method로 분석 하였다(Aebi, 1984). 조직 균질액에 20 mM H2O2 용액을 첨가하여 microplate reader의 kinetic method를 이용하 여 20분간 240 nm에서 흡광도의 변화를 측정하여 활성을 평가하였다. SOD 활성은 spectrophotometric method로 분석하였다(McCord와 Fridovich, 1969). 조직 균질액에 3 mM xanthine을 첨가하고 0.2 mM WST-1, xanthine oxi- dase 용액을 첨가하여 37°C에서 15분간 반응시키고 450 nm에서 흡광도를 측정하여 활성을 평가하였다. GPx 활성 은 spectrophotometric method로 분석하였다(Smith 등, 1988). 조직 균질액에 3.5 mM reduced glutathione을 첨 가하고 0.5 U/mL glutathione reductase를 첨가한 후 2.5 mM NADPH를 첨가하였다. 마지막으로 30 mM tert-butyl hydroperoxide를 넣어 반응시킨 후 340 nm에서 1분 간격 으로 20분 동안 흡광도를 측정하여 활성을 평가하였다.
CAT, SOD, GPx 활성은 U/mg protein으로 나타내었다.
통계 분석
실험 결과는 평균±표준오차로 나타내었고, 실험 결과들 의 유의성을 검정하기 위해 SPSS(PASW Statistics 23) 프 로그램(IBM Inc., Armonk, NY, USA)의 Student’s
t
-test 와 Duncan’s multiple range test를 이용하여 사후검정을 실시하였다(P
<0.05).결과 및 고찰
미강의 한계운동수행능력 향상 효과
미강의 한계운동수행능력을 측정하기 위해 Ex-CON, Ex-RB로 나누었으며 두 군 간에 마우스의 몸무게는 유의차 가 없게 하였으며 각 군들은 한계운동수행능력 평가전 유영 운동시간에 있어 유의차가 없게 그룹화하였다.
실험기간 동안 매일 미강을 투여하였으며 한계유영운동 은 실험기간 동안 세 번 측정하였다. 실험군 간에 체중과 조직(간, 신장, 비장, 근육) 무게 및 간 독성 지표인 aspartate amino transferase(AST)와 alanine amino transferase (ALT) 활성은 유의적인 차이를 나타내지 않았다(data not shown). 최대유영운동 시간을 측정한 결과 미강 투여 후
14일까지는 유의적 차이가 나타나지 않았으나 21일째에 Ex-CON과 비교하여 Ex-RB의 최대유영운동 시간이 유의 적으로 증가(
P
<0.05)함을 나타내었다(Fig. 1). 이는 미강 섭 취가 최대유영운동시간을 증가시켰다는 것을 확인할 수 있 었다.운동 후 혈중 젖산 농도의 변화
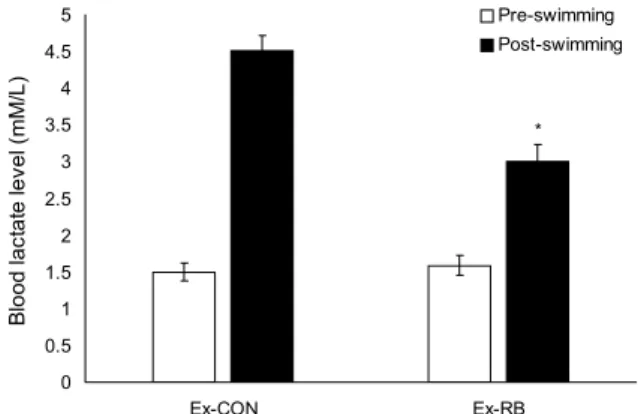
운동이나 무산소 대사 중에 빠른 에너지 생산을 위해 글루 코오스가 완전히 분해되지 못하고 해당과정만 거쳐 에너지 를 생성하게 되며, 이때 발생하는 대사산물이 젖산이다. 젖 산은 운동 후에 피로의 정도를 알 수 있는 하나의 지표이며 젖산의 과다 축적은 조직과 혈액에 산성화로 근육의 뭉침을 유발하고 지속적인 운동을 하지 못하게 한다(Finsterer, 2012). Fig. 2에서 운동 전 젖산의 농도는 Ex-CON과 Ex- RB에 있어 서로 유의적 차이를 나타내지 않았으나 10분 동
0 0.2 0.4 0.6 0.8 1 1.2 1.4 1.6
Ex-CON Ex-RB
Serum NEFA level (mEq/L) .
Pre-exercise
Post-exercise *
Fig. 3. Effect of rice bran on serum NEFA level. The Ex-CON
and Ex-RB of NEFA level was evaluated before and 10 min after forced swimming with a current flow of 8 L/min. The Ex-RB was orally given rice bran at a dose of 1 g/kg b.w./d for 21 days. Data are expressed the mean±SE. The asterisk above the bar is statistically different from Ex-CON group by Student’st-test (
*P<0.05).
0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8
CON Ex-CON Ex-RB
Muscle glycogen (mg/g tissue) .
a
a
b
Fig. 4. Effect of rice bran on muscle glycogen level. The CON
was sedentary group. The Ex-CON and Ex-RB were sacrificed 4 after exhaustive swimming with current flow at 8 L/min. The Ex-RB group was orally given rice bran at a dose of 1 g/kg b.w./d for 21 days. Data are expressed the mean±SE. Means with different letters above the bars are statistically different by Duncan’s multiple range test (P<0.05).안 운동 후 젖산의 수치는 Ex-CON과 Ex-RB가 모두 증가 하였다. Ikeuchi 등(2006)의 연구에서도 운동 후에 혈중 젖 산의 양이 높아지는 것을 나타내었다. 그러나 증가하는 양은 Ex-CON보다 Ex-RB가 유의적으로(
P
<0.05) 낮았다. 이러 한 결과는 미강의 섭취가 운동 중 생성되는 혈중 젖산의 축 적을 억제하여 젖산으로 생기는 근육 피로를 막아 장시간 운동을 할 수 있게 한 것으로 생각된다.운동 후 혈중 유리지방산 농도의 변화
한계운동을 할 경우 장시간 원활한 운동을 위해서는 글루 코오스보다 지방의 소모가 높아야 젖산 축적을 억제할 수 있 으며 많은 에너지를 생성할 수 있다(You 등, 2015b). NEFA 는 혈장 중의 유리형 지방산으로 여러 조직으로 운반되어 β-산화에 의해 에너지를 생성하거나 재에스테르화에 이용 된다(Ikeuchi 등, 2006). 특히 운동 시에는 NEFA가 근육으 로 운반되어 지속적인 운동을 하는 데 에너지원으로 사용된 다. 운동 전과 후 NEFA 수준을 측정한 결과 운동 전 Ex- CON과 Ex-RB의 NEFA 수준은 유의적인 차이가 없었으나, 운동 후에는 Ex-CON보다 Ex-RB가 유의적인 증가(
P
<0.05) 를 보였다(Fig. 3). 이러한 결과는 Murase 등(2005)의 연구 에서도 녹차 추출물 섭취로 인한 장시간 운동 시 NEFA 양이 증가하는 것과 유사한 경향을 보인다. 이는 미강의 섭취가 운동 시 혈중 젖산 수준을 낮출 뿐만 아니라 NEFA 수준을 평소보다 빠르게 증가시켜 에너지원으로 사용하여 운동수 행능력을 향상시킨 것으로 생각된다.운동 후 근육 내 글리코겐 함량의 변화
글리코겐은 혈중 글루코오스가 높을 때 간이나 근육 등 여러 조직에 글루코오스를 저장하는 형태이다(Xirouchaki 등, 2016). 글리코겐은 운동 시 중요한 에너지원으로 오랜 시간 동안 운동하게 되면 조직 내 글리코겐 함량이 감소하게
된다. 한계운동 후 근육 내 글리코겐의 함량은 Ex-CON이 CON보다 유의적으로(
P
<0.05) 낮았다(Fig. 4). 그러나 Ex- RB의 글리코겐 함량은 CON과 서로 유의적인 차이를 보이지 않았으며 Ex-CON과 비교했을 때 유의적인 증가(P
<0.05) 를 보였다. 이러한 결과는 Cantó 등(2010)의 연구에서도 운동 후 글리코겐이 감소하는 결과를 보여주었으며, You 등 (2015b)의 연구에서는 복분자 추출물에 의해 글리코겐이 보호되는 결과를 나타내었다. 결과적으로 운동을 통해 근육 내 글리코겐 함량이 감소하였으나 미강의 투여로 운동 후 글리코겐의 소모를 줄여주는 결과를 보였다. 이는 미강이 NEFA 수준을 증가시켜 지방의 소비는 증가시키고 글리코겐 은 절약함으로 에너지 생성량이 더 많은 지방을 에너지원으 로 사용하여 지속적인 운동을 가능하게 한 것으로 생각된다.운동 후 근육 내 항산화 활성 변화
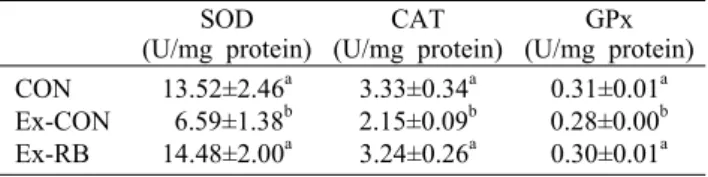
생물체에서 에너지를 생산하기 위해서 산화는 필수적이 다. 평상시에는 활성산소가 형성되어도 체내 효소에 의해 평 형상태를 유지하여 영향을 미치지 않지만 한계유영운동을 하게 되면 미토콘드리아에서 에너지 생성을 하거나 β-산화 로 인해 활성산소가 증가하여 산화효소와 불균형을 이루게 된다(Ji, 1999). 이렇게 활성산소와 산화효소의 불균형은 장 시간 동안 운동을 하지 못하게 한다(Lima 등, 2013). SOD는 활성산소를 H2O2와 산소로 만드는 효소이고 CAT와 GPx는 H2O2를 물과 산소로 분해하는 효소이다. 이러한 기전은 정 상 세포에서 잘 유지된다. 한계유영운동 후 항산화 활성을 나타내는 SOD, CAT, GPx는 Ex-CON에서는 CON보다 유 의적으로 감소하였으나 Ex-RB는 CON과 유의적인 차이를 볼 수 없었으며 Ex-CON에 비해 유의적 증가(
P
<0.05)를 나타내었다(Table 1). 많은 연구들에서 한계운동은 효소 활 성에 영향을 주어 낮은 항산화 활성을 나타내지만(Flis 등, 2018; Yan와 Hao, 2016), 인체에 유용한 생리활성물질인Table 1. Effects of rice bran on antioxidant enzyme activities
in muscle(U/mg protein)SOD CAT
(U/mg protein) GPx (U/mg protein) CON
Ex-CON Ex-RB
13.52±2.46a 6.59±1.38b 14.48±2.00a
3.33±0.34a 2.15±0.09b 3.24±0.26a
0.31±0.01a 0.28±0.00b 0.30±0.01a The CON was sedentary group. The Ex-CON and Ex-RB were sacrificed 4 after exhaustive swimming with current flow at 8 L/min. The Ex-RB was orally given rice bran at a dose of 1 g/kg b.w./d for 21 days. Data are expressed the mean±SE. The alphabet within a column are statistically different by Duncan’s multiple range test (P<0.05).
토코페롤, 레시틴 등은 항산화 능력을 향상시켜준다(Nor Azman 등, 2018; Nasab 등, 2019)고 보고되어 있다. 결국 한계유영운동에 의해 증가한 활성산소가 체내에 낮은 항산 화 활성을 나타내지만 미강을 섭취함으로써 생리활성물질 인 오리자놀, 토코페롤, 레시틴, 스테롤 등(Nagendra Pra- sad 등, 2011)이 체내 항산화 효소 활성을 유지시켜 한계운 동수행능력을 향상한 것으로 보인다.
요 약
본 연구에서는 미강을 마우스에 투여한 후 한계운동수행능 력을 평가하였다. 한계유영운동 시간은 비투여군(Ex-CON) 보다 미강 투여군(Ex-RB)에서 유의적으로 높게 나타났다.
미강 투여로 인해 체내에 피로물질인 젖산의 축적을 억제하 였으며, 혈청 유리지방산의 농도를 높여 글리코겐의 소모를 줄이고 유리지방산을 에너지원으로 하여 에너지 고갈을 낮 추었다. 이로 인해 근육 내 글리코겐 함량이 운동군에 비해 미강 투여군에서 높게 나타났다. 또한 항산화 효소 활성을 증가시켜 한계유영운동이 증가함을 확인하였다. 이상의 결 과들로 미강의 섭취가 한계유영운동 수행능력을 향상시키 는 것을 확인하였다.
감사의 글
본 연구는 산업통상자원부와 한국산업기술진흥원이 지원하 는 경제협력권산업육성사업으로 수행된 연구 결과입니다 (과제번호: R0004933).
REFERENCES
Aebi H. Catalase in vitro. Methods Enzymol. 1984. 105:121-126.
Cantó C, Jiang LQ, Deshmukh AS, Mataki C, Coste A, Lagouge M, et al. Interdependence of AMPK and SIRT1 for metabolic adaptation to fasting and exercise in skeletal muscle. Cell Metab. 2010. 11:213-219.
Cha MH, Kim YK. Moderating effect of health motivation, health concern and food involvement on the relationship be- tween consumption value and purchasing intentions of healthy functional food. J Korean Soc Food Sci Nutr. 2008. 37:1435-
1442.
Durkot MJ, De Garavilla L, Caretti D, Francesconi R. The effects of dichloroacetate on lactate accumulation and endurance in an exercising rat model. Int J Sports Med. 1995. 16:167-171.
Finsterer J. Biomarkers of peripheral muscle fatigue during ex- ercise. BMC Musculoskelet Disord. 2012. 127:218. doi: 10.
1186/1471-2474-13-218.
Fisher-Wellman K, Bell HK, Bloomer RJ. Oxidative stress and antioxidant defense mechanisms linked to exercise during car- diopulmonary and metabolic disorders. Oxid Med Cell Lon- gev. 2009. 2:43-51.
Flis DJ, Dzik K, Kaczor JJ, Halon-Golabek M, Antosiewicz J, Wieckowski MR, et al. Swim training modulates skeletal mus- cle energy metabolism, oxidative stress, and mitochondrial cholesterol content in amyotrophic lateral sclerosis mice. Oxid Med Cell Longev. 2018. doi: 10.1155/2018/5940748.
Gierus M, Rocha JBT. Forage substitution in a grain-based diet affects pH and glycogen content of semimembranosus and semitendinosus rabbit muscles. J Ani Sci. 1997. 75:2920-2923.
Howes MJ, Simmonds MS. The role of phytochemicals as mi- cronutrients in health and disease. Curr Opin Clin Nutr Metab Care. 2014. 17:558-566.
Ikeuchi M, Koyama T, Takahashi J, Yazawa K. Effects of astax- anthin supplementation on exercise-induced fatigue in mice.
Biol Pharm Bull. 2006. 29:2106-2110.
Ji LL. Antioxidants and oxidative stress in exercise. Proc Soc Exp Biol Med. 1999. 222:283-292.
Joy JM, Lowery RP, Wilson JM, Purpura M, De Souza EO, Wilson SM, et al. The effects of 8 weeks of whey or rice protein supplementation on body composition and exercise performance. Nutr J. 2013. 12:86.
Kim HK, Chang HK, Lee MR, Son YJ, Han SJ, Yang NY, et al. Influence of malnutrition and social network on health-re- lated quality of life in elders. J Korean Acad Fundam Nurs.
2013. 20:98-107.
Kim JH, Pan JH, Lee ES, Kim YJ. L-Carnitine enhances ex- ercise endurance capacity by promoting muscle oxidative me- tabolism in mice. Biochem Biophys Res Commun. 2015. 464:
568-573.
Lamou B, Taiwe GS, Hamadou A, Houlray J, Atour MM, Tan PV. Antioxidant and antifatigue properties of the aqueous ex- tract of Moringa oleifera in rats subjected to forced swim- ming endurance test. Oxid Med Cell Longevity. 2016. Article ID: 3517824. http://dx.doi.org/10.1155/2016/3517824 Lima FD, Stamm DN, Della-Pace ID, Dobrachinski F, de Car-
valho NR, Royes LF, et al. Swimming training induces liver mitochondrial adaptations to oxidative stress in rats submitted to repeated exhaustive swimming bouts. PLoS One. 2013.
8:e55668. doi: 10.1371/journal.pone.0055668.
Matsumoto K, Ishihara K, Tanaka K, Inoue K, Fushiki T. An adjustable-current swimming pool for the evaluation of endur- ance capacity of mice. J Appl Physiol. 1996. 81:1843-1849.
McCord JM, Fridovich I. Superoxide dismutase. an enzymic function for erythrocuprein (hemocuprein). J Biol Chem. 1969.
244:6049-6055.
Murase T, Haramizu S, Shimotoyodome A, Nagasawa A, Toki- mitsu I. Green tea extract improves endurance capacity and increases muscle lipid oxidation in mice. Am J Physiol Regul Integr Comp Physiol. 2005. 288:R708-R715.
Nagendra Prasad MN, Sanjay KR, Shravya Khatokar M, Vis- maya MN, Nanjunda Swamy S. Health benefits of rice bran-
A review. J Nutr Food Sci. 2011. 1:1-7.
Nasab ME, Takzaree N, Saffari PM, Partoazar A. In vitro anti- oxidant activity and in vivo wound-healing effect of lecithin
liposomes: a comparative study. J Comp Eff Res. 2019. 8:
633-643.
Nor Azman NHE, Goon JA, Abdul Ghani SM, Hamid Z, Wan Ngah WZ. Comparing palm oil, tocotrienol-rich fraction and α-tocopherol supplementation on the antioxidant levels of old- er adults. Antioxidants. 2018. 7:74. doi: 10.3390/antiox70600 74.
Powers SK, Jackson MJ. Exercise-induced oxidative stress: cel- lular mechanisms and impact on muscle force production.
Physiol Rev. 2008. 88:1243-1276.
Seon YK, Goo HM, Park KK, Yang EJ. Characteristics of enzy- matic hydrolysates of rice bran and rice protein by mixing ratio and hydrolysis times. J Korean Soc Food Sci Nutr. 2016.
45:1460-1466.
Smith IK, Vierheller TL, Thorne CA. Assay of glutathione re- ductase in crude tissue homogenates using 5,5’-dithiobis(2-ni-
trobenzoic acid). Anal Biochem. 1988. 175:408-413.
Xirouchaki CE, Mangiafico SP, Bate K, Ruan Z, Huang AM, Tedjosiswoyo BW, et al. Impaired glucose metabolism and exercise capacity with muscle-specific glycogen synthase 1 (gys1) deletion in adult mice. Mol Metab. 2016. 5:221-232.
Yan F, Hao H. Effects of Laminaria japonica polysaccharides on exercise endurance and oxidative stress in forced swim- ming mouse model. J Biol Res. 2016. 23:7. doi: 10.1186/s40 709-016-0049-4.
You Y, Kim K, Jeong H, Park J, Kim OK, Jun W. Ethanolic extract from Rubus coreanus enhanced swimming capacity in mice. Food Sci Biotechnol. 2015a. 24:2213-2217.
You Y, Kim K, Yoon HG, Choi KC, Lee YH, Lee J, et al. Sasa