T-세포형 급성림프구성백혈병에서 NOTCH1 의 변형과 표적치료제 개발
노정희ㆍ전인상
가천대학교 의학전문대학원 소아과학교실
Alteration of NOTCH1 in T-cell Acute Lymphoblastic Leukemia and Development of Target Therapeutic Agent
Jung Hee Rho, M.D. and In-sang Jeon, M.D.
Department of Pediatrics, Graduate School of Medicine, Gachon University, Incheon, Korea
T-cell acute lymphoblastic leukemia (T-ALL) accounts for approximately 10-15% of entire ALL in children. The outcome of T-ALL has been improved through the intensified ther- apeutic strategy, however, it is still a more aggressive disease. In T-ALL a couple of transcription factor oncogenes are known to be relocated to the juxtaposition of T-cell receptor genes, potent promoter, by chromosome translocation. However the incidence of each chimeric gene formation in T-ALL is less than 5% and their clinical significance as a prognostic marker is lacking. A decade ago it was identified that activating mutations in NOTCH1 in about 60% of T-ALL. After then, activating NOTCH1 mutations present in T-ALL have been extensively investigated with regard to understanding its molecular pathogenesis, its prognostic significance, and developing molecularly tailored novel agents. Small molecule γ-secretase inhibitor, blocking a proteolytic step required for crea- tion of a fragment of NOTCH intracellular domain which actually act as a controller of its target gene expression, was tried as a target therapeutic drug for T-ALL. Although outcome of this drug was not satisfactory, challenges have been launched to develop new drugs which specifically act on the aberrant behavior of mutated NOTCH1 in T-ALL.
pISSN 2233-5250 / eISSN 2233-4580 http://dx.doi.org/10.15264/cpho.2014.21.1.1 Clin Pediatr Hematol Oncol 2014;21:1∼8
Received on March 17, 2014 Revised on April 1, 2014 Accepted on April 5, 2014
Corresponding Author: In-sang Jeon Department of Pediatrics, Gachon University Gil Medical Center, 21, Namdong-daero 774 beon-gil, Namdong-gu, Incheon 405-760, Korea
Tel: +82-32-460-8382 Fax: +82-32-460-3224 E-mail: [email protected]
Key Words: T-ALL, NOTCH1, Gamma-secretase inhibitor
서 론
T-세포형 급성림프구성백혈병(T-cell acute lymphoblastic leukemia, T-ALL)은 전체 급성림프구성백혈병(acute lympho- blastic leukemia, ALL)에서 소아에선 10-15%, 성인에선 25%
를 차지한다. 임상적으로는 진단 시 백혈구수가 크게 증가되 어 있으며 종격동종괴 및 중추신경계 침범이 흔하고 남자에선 고환 침범이 흔하다. 최근에는 강화된 치료법의 개발로 치료 율이 향상되었으나 아직도 상당수의 T-ALL은 항암약물에 저 항성을 보이며 재발한다[1]. 분자생물학적 이해도 미흡하여
B-ALL에서 특이한 염색체전좌가 관찰되며 이들에 의해 변형
된 암유전자들이 백혈병생성에 관여하는 병태생리가 잘 알려
져 있으며 또한 이들이 중요한 예후인자로 알려진 것과 대조
를 이룬다. T-ALL에선 약 35%에서 다양한 염색체전좌가 발견
되며 이로 인해 생성된 융합유전자가 알려져 있다. 이들은 주
로 T-세포 수용체(T-cell receptor, TCR) 유전자와 융합되는
TAL1, LMO1, HOX11과 같은 전사인자들이다. 그러나 이들은
발생 빈도가 각각 5% 내외로 발생 빈도가 높지 않으며 분자생
물학적 연구도 충분하지 못하다[2]. 이들 이외에 염색체전좌
t(7;9)(q34;q34.3)에 의해 TCR-β와 융합된 형태로 처음 알려
진 NOTCH1 유전자가 있는데 이 전좌의 빈도는 약 1% 밖에
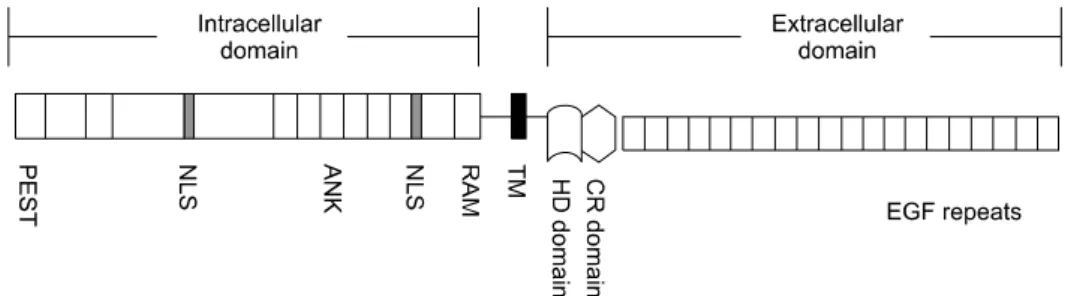
Fig. 1. Schematic drawing of NOTCH1 structure. NOTCH1 is a heterodimer composed of extracellular domain and intracellular domain. These are noncovalently joined through a structural motif, heterodimerization domain (HD). EGF repeats, epidermal growth factor repeats; CR, cysteine rich; TM, transmembrane; RAM, RBP-Jkappa-associated module; NLS, nuclear localization signals; ANK, ankyrin-like; PEST, proline, glutamic acid, serine, threonine rich.
되지 않는다[3]. 그러나 약 10년 전에 이러한 염색체전좌와 무 관하게 NOTCH1 돌연변이가 T-ALL의 약 60%에서 관찰되는 것이 밝혀졌다[4].
NOTCH에 의해 전달되는 신호는 다양한 기관과 조직이 정 상적으로 분화하는데 중심적 역할을 한다. 특히 NOTCH1은 조혈과정에 관여하는데 특히 원시조혈모세포로부터 T-세포 계열로 분화하는데 결정적인 역할을 한다. 또한 NOTCH1의 돌연변이가 T-세포를 형질전환시켜 T-ALL을 유도하는 과정은 비교적 자세히 알려져 있다[5]. 임상적으로는 NOTCH1의 돌 연변이의 동반과 T-ALL의 예후에 대해서는 여러 의견이 있다.
또한 변형된 NOTCH1의 신호전달에 의해 백혈병이 발생하는 과정을 표적으로 하는 표적치료로 NOTCH1의 활성화를 방해 하는 약물인 γ-secretase 방해제(γ-secretase inhibitor, GSI)를 갖고 임상시험이 시행되었으나 치료 효과는 만족스럽지 못했 다[6]. 그러나 새로운 약물을 개발하기 위해 많은 연구가 진행 중으로 가까운 미래에는 NOTCH1에 의해 변형된 신호전달을 차단하는 치료제가 나올 것으로 생각된다.
본 논문에서는 NOTCH1 유전자의 구조, 정상적인 신호전 달 과정을 비롯하여 T-ALL에서 관찰되는 NOTCH1 돌연변이 에 의해 T-세포의 형질전환되는 과정을 설명하고 현재 진행 중인 표적치료에 대해 알아보고자 한다.
NOTCH 신호전달 경로
초파리의 Drosophila NOTCH 유전자에 돌연변이가 생기면 날개에 ‘마디(notch)’가 생긴다. 이 유전자의 인체 상동유전자 가 NOTCH 유전자이다. 인체 NOTCH 유전자에는 4종류가 있 다. 그 중의 하나인 NOTCH1은 T-ALL 환자에서 관찰되는 t(7;9)(q34;q34.3)에서 처음 확인되었다[4]. NOTCH 유전자는 진화학적으로 잘 보존되어 있으며 세포의 분화, 증식, 생존,
세포사멸 과정을 매개되는 신호전달에 관여한다. NOTCH 단 백질은 세포막을 한번만 통과하는 신호전달수용체로 세포막 외 부분과 세포내 부분으로 되어 있으며 이 두 부분은 이형이 합체 영역(heterodimerization domain, HD)에 의해 비공유 결합되어 있다[5]. NOTCH 수용체는 주위 세포의 표면에 위치 하는 Delta-Serrate-Lag2 (DSL)계 리간드에 의해 활성화되어 신호전달을 시작한다. 세포외 부분은 DSL 리간드와의 반응을 매개하는 EGF 반복 구역과 외부 자극 없이 NOTCH가 활성화 되는 것을 방해하는 시스틴풍부 구역(cysteine-rich region, CR)과 HD가 있다. HD에는 금속분해효소(metalloproteinase) 에 의해 분해되고 절단되는 부위가 있는데 CR이 이 부위를 감싸고 있어 리간드-수용체 상호작용이 없으면 NOTCH 단백 질이 분해 절단되는 것을 구조적으로 막아 NOTCH가 활성화 되는 것을 막는다. 세포내 부분은 여러 구역으로 되어 있는데 RBP-Jkappa-associated module (RAM)구역과 ankyline (ANK) 구역은 NOTCH 단백질과 DNA 부착단백질인 DNA-binding protein CBF/RBP-J,Su(H), Log1 (CSL)과의 상호작용을 매개 한다. C-말단 부위에 위치하는 proline, glutamic acid, serine, theonine rich (PEST)구역은 NOTCH 단백질이 분해되는 과정 에 관여한다(Fig. 1).
현재까지 알려진 NOTCH의 신호전달 과정은 다음과 같다
[7]. 먼저 주위 세포의 표면에 위치하는 리간드 DSL이 NOTCH
의 세포외 부분인 EGF 반복 구역에 부착하면서 신호전달은
시작된다. 이 부위에 리간드가 붙게 되면 NOTCH의 활성화를
억제하는 CR과 HD는 구조적으로 바뀌어 ADAM계 금속분해효
소에 의해 NOTCH의 HD의 C-말단부위는 일차적으로는 절단된
다. 일차 절단 이후에 세포외 부위가 짧아진 NOTCH 단백질은
다시 presenilin-의존성 γ-secretase에 의해 이차로 NOTCH는
절단되어 세포내 부위를 세포외 부위와 분리시켜 세포내 부위
로만 구성된 NOTCH 세포내 구역(NOTCH intracellular do-
Fig. 2. The NOTCH1 signalling path- way. (a) Interaction of the NOTCH1 receptor with Delta-Serrate-Lag2 (DSL) ligand triggers two successive proteolytic cleavages, first by an ADAM proteinase and subsequently by the γ-secretase complex, which liberates the intracellular domain (NICD). (b) The NICD translocates to the nucleus, where it interacts with DNA-binding protein RBP-J (CSL) to generates a binding site for coactivator, Mastermind-like proteins (MAML), to activate the expression of NOTCH1 target genes. (c) NICD/ CSL/MAML complex recruits the RNA polymerase II holoenzyme to phosphorylate the PEST domain of NICD, which facilitates ubiquity- lation by FBXW7. Finally ubiquity- lated NICD is degraded by protea- some.
main, N
ICD)을 세포막으로부터 분리시킨다. 분리된 N
ICD는 핵으 로 이동하여 표적유전자의 전사를 조절하게 된다. N
ICD는 핵에 서 CSL이라는 전사인자와 함께 표적유전자의 표현을 조절한다.
CSL은 N
ICD의 신호전달이 없는 상태에서는 NOTCH에 의해 조 절되는 표적유전자의 표현을 억제한다. 그러나 N
ICD이 핵으로 이동하여 연합하게 되면 CSL은 표적유전자의 표현을 억제하는 조-억제자(co-repressor)를 밀어내고 Mastermind-like (MAML) 와 CREB-binding protein (CBP) /p300 등의 조-활성자(co-acti- vator)를 끌어들여 NOTCH의 표적유전자들은 표현된다. 이런 과정에 의해 발현되는 NOTCH1의 표적유전자로는 pre-Tα , HES1, C-MYC, IGF1R, IL7R, NRARP, DELTEX1, RUNX3 등이 있다. 한편 조혈과정에 중요한 역할을 하는 DNA 부착 단백질인 IKAROS는 CSL과 동일한 표적유전자의 염기서열에 경쟁적으로 부착하여 N
ICD의 작용을 방해한다. 실제로 IKAROS의 유전정보 가 입력되어 있는 유전자의 돌연변이가 T-ALL이나 림프종에서 흔하게 발생하여 IKAROS와 NOTCH1의 상호작용이 T-ALL의 발생에 관여하는 것으로 생각된다[8]. 신호전달을 끝낸 NOTCH 는 먼저 CyclinC:CDK8에 의해 NOTCH의 PEST 구역이 인산화 된다. 인산화된 PEST 구역에 F-box 단백질인 FBWX7이 부착하 게 되면 NOTCH는 유비큐틴화되어 프로테아솜에 의해 분해된 다(Fig. 2).
T-세포 발달과정에서 NOTCH1의 역할
NOTCH1 신호전달은 T-세포 분화 과정에서 중요한 역할을
한다. 이러한 사실은 골수에서 NOTCH1이나 CSL의 작용을
억제시켰을 때 가슴샘에서 조기 T-세포로 분화가 일어나지 않
고 이소성 미성숙 B-세포로 분화하는 것으로부터 알 수가 있
다[9]. 골수에서 가슴샘으로 이동한 림프계 전구세포는 T-세포
와 B-세포로 분화할 수 있다. NOTCH1은 이 세포에 작용하여
T-세포계열의 전구세포로 분화시켜 가슴샘 조기 T-전구세포
(earliest T-cell progenitor)로 분화시킨다. 이 세포는 CD4
-CD8
-인
이중음성1 (double negative 1, DN1) T-세포이다. 이후 DN1
은 DN2, DN3, DN4로 분화하는데 이 과정에 NOTCH1이 관
여한다. 특히 NOTCH1은 E2A와 상호작용을 하여 DN3에서
T-세포 수용체 αβ사슬이 형성되게 유도한다. 이후 DN4는 이
중양성(double positive) T-세포로 분화되고 CD4+ T-세포 또
는 CD8+ T-세포로 분화해 간다. 이 과정에서 NOTCH1은 관
여하지 않는 것으로 알려져 있다. NOTCH1은 CD4+ T-세포가
T
H2로 증식하는 것을 촉진하며 T
H1로 분화되는 것을 방해하는
것으로 알려져 있으나, 대체적으로 분화 후반부 세포보다는
전구세포에서 T-세포로 분화되는 과정에 관여하는 것으로 알
려져 있다.
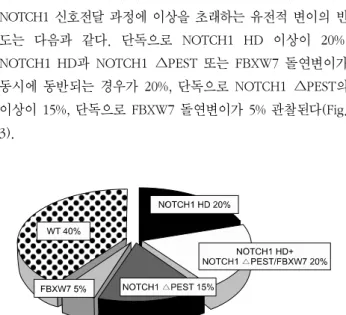
Fig. 3. Prevalence of NOTCH1 mutations observed in T-ALL.
WT, wild type.
T-ALL에서 관찰되는 NOTCH1의 변이
처음으로 NOTCH1 유전자 이상이 T-ALL에서 발견된 것은 1990년대 초반이다. T-ALL에서 t(7;9)(q34;q34.3)에 의해 NOTCH1 유전자가 절단되어 이 유전자의 3’ 부분이 TCR-β 유전자와 합쳐져 TCR-β/NOTCH1 융합유전자가 만들어진다 [3]. 이 융합유전자는 NOTCH1 유전자의 5’ 부분에 있는 HD-CR이 없거나 세포내 부분까지 없는 상태로 외부 리간드 작용 없이 연속적으로 활성화된다. 염색체전좌에 의해 변형된 NOTCH1이 T-ALL의 발생에 관여한다는 사실은 동물 실험에 서 간접적으로 확인이 되었다. 조혈모전구세포를 N
ICD가 표현 되는 레트로바이러스로 감염시킨 뒤 마우스에 이식시키면 신 속하게 T-ALL이 발생한다. 동물 실험에서 NOTCH1의 활성화 는 T-ALL의 발생에 관여한다는 여러 실험적 증거가 있었으나 실제로 NOTCH1의 이상이 동반되는 t(7;9)(q34;q34.3)가 T-ALL 환자에서 1% 정도 밖에 되지 않아 크게 주목을 받지 못 하였다. 그러나 NOTCH1 유전자 자체의 돌연변이가 T-ALL 환자에서 60%나 발견되어 새로운 국면을 맞게 되었다[4]. 리 간드 없이도 NOTCH1 유전자의 돌연변이로 자발적으로 활성 화되거나, 활성화된 NOTCH1을 비활성화시키는 부위의 돌연 변이가 연속적으로 NOTCH1을 활성화시켜 T-ALL가 유도되는 것이 알려졌다. T-ALL에서 관찰되는 NOTCH1의 돌연변이는 다음과 같다[10].
1) HD 돌연변이
전체의 약 40%를 차지하는 가장 흔한 형태로, 돌연변이가 일어나는 부위에 따라 다음과 같이 나눌 수 있다.
(1) NOTCH1 HD 돌연변이 I 군(HD1): T-ALL에서 가장 흔하 게 돌연변이가 일어나는 부위는 NOTCH1 유전자의 엑손 26 과 27로 HD에 대한 유전코드가 있는 부위이다. HD는 CR과 함께 복합체를 이루어 리간드가 없는 상태에서 NOTCH1의 절 단을 방해하지만 이 부위의 돌연변이로 HD-CR 복합체는 안 정성을 잃게 되어 NOTCH1은 절단되어 활동성이 있는 N
ICD을 만들어내어 연속적으로 NOTCH1 신호가 전달된다. 이 때 주 로 일어나는 돌연변이는 하나의 아미노산이 본래와 다른 것으 로 변하거나 유전자틀(in-frame)을 이루는 유전자 결손이나 삽입이 작은 단위로 일어난다. 이 형태는 전체 NOTCH1 이상 의 약 20%를 차지한다.
(2) NOTCH1 HD 돌연변이 II 군(HD1I): HD의 원위 부위에
커다란 크기의 삽입이 일어나 CR-HD 복합체로부터 멀어진 금속분해효소의 작용 부위가 밖으로 노출되어 NOTCH1이 절 단되고 N
ICD가 형성되어 NOTCH1의 활성화가 계속된다.
(3) NOTCH1 H1545P: CR 부위의 돌연변이로 CR-HD 복합 체의 안정성이 약해져 금속분해효소의 작용 부위를 보호하지 못하게 된다.
2) PEST 돌연변이(NOTCH1 ∆PEST)
이 돌연변이는 전체 이상의 20-25%를 차지한다. NOTCH1 의 C-말단 부위에 있는 PEST 부위에서 틀이동 또는 염기치환 에 의해 미리 정지 코돈이 만들어져 정상적인 PEST가 만들어 지지 않는다. 그 결과 N
ICD와 FBXW7의 상호작용이 이루어지 지 않게 되어 활성화된 N
ICD분해가 정상적으로 되지 않게 되 어 NOTCH1의 작용이 지속된다.
3) FBXW7 돌연변이
전체 돌연변이의 약 15%를 차지한다. NOTCH1의 PEST 부 위와 상호작용을 하는 FBXW7의 돌연변이 또는 결손에 의해 활성화된 N
ICD을 분해시키지 못 해 양상이 PEST 돌연변이와 비슷하다. 그러나 이 돌연변이는 NOTCH1 이 외에 FBXW7에 의해 분해되는 다른 암단백질인 C-MYC, JUN, Cyclin-E, m-TOR도 분해되지 않아 PEST 돌연변이에 비해 발암 효과가 크다.
T-ALL에서 이러한 변이들이 단독으로 존재하기도 하나 여 러 변이들이 동반되어 나타나기도 한다. T-ALL에서 관찰되는 NOTCH1 신호전달 과정에 이상을 초래하는 유전적 변이의 빈 도는 다음과 같다. 단독으로 NOTCH1 HD 이상이 20%, NOTCH1 HD과 NOTCH1 ∆PEST 또는 FBXW7 돌연변이가 동시에 동반되는 경우가 20%, 단독으로 NOTCH1 ∆PEST의 이상이 15%, 단독으로 FBXW7 돌연변이가 5% 관찰된다(Fig.
3).
T-ALL로 형질 변경이 일어나기 위해서는 계속적으로 NOTCH1의 표현이 있어야 하는 것이 동물실험에서 알려져 있 다. T-ALL에서 관찰되는 여러 종류의 돌연변이들은 NOTCH1 을 표현하는 정도에 따라 백혈병으로 형질유도하는 정도가 다 르다. 즉, 강력하게 NOTCH1을 활성화시키는 NOTCH1 HD1-
∆PEST형이나 NOTCH1 HD2는 원시 조혈모세포를 T-세포형 전구세포로 형질전환시킨다. 이에 반해 NOTCH1을 활성화시 키는 힘이 약한 NOTCH1 HD1이나 NOTCH1 ∆PEST 돌연변 이형은 조혈모세포를 T-세포 계열로 분화를 유도하지만 백혈 병으로 형질전환시키지는 못 한다. 이러한 돌연변이들은 다른 돌연변이에 의해 백혈병 전 단계에 있는 세포를 형질전환시킨 다. 이러한 예로 k-ras 암유전자를 표현하고 있는 원시조혈모 세포에 이들 약한 돌연변이가 작용하여 T-ALL로 형질전환되 는 것을 촉진시키는 것이 알려져 있다[11]. 이러한 사실은 T-ALL의 치료제로 NOTCH1에 대항하는 약물을 고안하는데 매우 중요하다.
NOTCH1에 의해 조절 받아 T-ALL 형질전환에 관여하는 경로
NOTCH1에 의해 조절 받는 경로와 표적유전자를 아는 것 은 새로운 표적치료제를 개발하는데 매우 중요하다. 지난 수 년 동안 NOTCH1에 의해 조절을 받거나 상호작용을 하여 T-ALL의 발생에 관여하는 유전자와 신호전달 경로에 대한 집 중적인 연구가 있었다. 이들 연구에 의하면 NOTCH1은 여러 직간접적인 방법으로 정상 혈구세포를 백혈병세포로 형질전 환시키고 유지시키는데 중심적인 역할을 하는 것이 알려졌다 [12].
1) C-MYC 유전자를 직접적으로 과표현시킨다.
C-MYC 유전자는 잘 알려진 암유전자성 전사인자로 이 유 전자는 세포의 성장을 촉진시킨다. NOTCH1은 직접적으로 C-MYC 유전자의 표현을 증진시키며 이로 인해 세포성장을 유도한다. NOTCH1/C-MYC에 의해 형성된 전사성 조절망은 T-세포를 형질전환시키는 것이 동물실험에서 알려져 있다.
2) PI3k-AKT-mTOR 경로의 활성화
PI3K-AKT-mTOR 신호전달경로는 성장인자 수용체의 신호 를 받아 다양한 세포반응을 조절한다. 특히 세포성장, 증식과 생존 같은 암형성과 관련이 있는 세포의 여러 작용에 관여한 다. NOTCH1은 PI3K-AKT 신호전달경로를 활성화시켜 T-세 포가 분화하는 과정에 관여한다. NOTCH1은 여러 기전을 통
해 PI3K-AKT-mTOR 신호전달을 항진시키는 것으로 보인다.
Palomero 등에 의하면 T-전구세포나 T-ALL 림프모세포에서 NOTCH1은 전사억제 기능을 갖고 있는 HES1의 발현을 증진 시킨다. 한편 HES1은 PI3K 신호전달경로의 음성 조절자인 PTEN의 표현을 억제한다. 결과적으로 NOTCH1은 T-전구세 포나 T-ALL 림프모세포에서 PI3K 신호전달경로를 활성화시 킨다. 또한 NOTCH1은 T-세포에서 LCK 타이로신키나제릍 통 해 AKT를 활성화시킨다[13]. 한편 NOTCH1은 T-전구세포나 T-ALL 림프모세포에서 PI3K의 상부 신호전달 분자인 인터루 킨 7 수용체 알파체인(IL7RA), 전구 T-세포수용체 알파 (PTCRA), 인슐린 유사 성장인자 1 수용체(IGF1R)를 조절하여 간접적으로 PI3K-AKT-mTOR 신호전달경로를 활성화시킨다.
3) G1/S 세포주기 진행의 촉진
NOTCH1은 조혈모세포를 세포주기 G1기에서 S기로 진행 을 촉진시켜 백혈병세포의 증식을 유도하는 것으로 알려져 있 다. T-ALL 세포에서 NOTCH1은 G1기에서 S로 진행하는 것을 조절하는 CDK4와 CDK6의 발현을 증진시키며 Cyclin D3 유 전자인 CCND3의 발현도 증진시킨다. 또한, CDK 방해제인 p27/KIPI과 p18/INK4C의 파괴를 증진시키고 S기 키나제 연 관 단백질 2 (S phase kinase-associated protein 2)의 전사를 증진시킨다. 이러한 여러 기전을 통해 NOTCH1은 조혈모전 구세포를 S기로 들어가게 하여 모세포의 증식을 유도하는 것 으로 생각된다.
4) NF-κB 신호전달의 활성화
NF-κB는 종양형성에 관여하는 신호에 관여하는 전사인자 인데 NOTCH1은 NF-κB의 활성을 증가시킨다. NOTCH1은 NF-κB의 표현을 증가시키고 NF-κ가 핵 내에 머무는 시간을 연장시키고 NF-κB의 작용을 방해하는 IκB를 분해하는 IκB 키나제의 작용을 활성화시켜 NF-κB의 활성을 증가시킨다.
NOTCH1과 NF-κB의 상호작용이 백혈병에 관여한다는 사실 은 T-ALL에서 NF-κB를 억제하면 백혈병이 호전되거나 NOTCH1이 세포를 형질전환시키는 데는 NF-κB가 반드시 필 요하다는 사실로 확실히 알 수 있다.
5) 에피제네틱 조절
NOTCH1은 Polycomb Regressive Complex 2 (PRC2)의 활
성을 방해한다. PRC2는 히스톤의 하나인 H3K27의 Lys27 부
위를 삼메틸화(trimethylation)시켜 유전자의 표현을 억제하는
역할을 한다. 이러한 작용에 의해 NOTCH1은 자신의 표적유
전자를 표현을 증진시키는 것으로 알려져 있다. 또한 PRC2의
주요 성분인 EZH2, SUZ12와 EED에 대한 유전정보가 코딩되 어 있는 유전자의 돌연변이가 T-ALL에서 흔하게 발견된다. 이 러한 것들을 종합해 보면 NOTCH1은 PRC2와의 상호작용을 통해 유전자의 표현을 조절하고 세포의 형질전환에 관여한다 는 사실을 시사하고 있다.
NOTCH1 변이와 T-ALL 예후와의 관계
T-ALL에서 NOTCH1의 돌연변이의 발생 빈도가 높아 이 변 이가 예후인자로써 어떤 역할을 하는가에 대한 여러 연구가 진행되었다. 연구 초기에는 NOTCH1의 이상을 동반한 T-ALL 이 예후가 양호하다는 보고가 있었다. 독일의 ALL-BFM 2000 연구는 T-ALL 소아 환자 157명을 대상으로 초기 치료반응성 과 장기간의 치료 결과를 분석하였다. 이들에 의하면 NOTCH1 돌연변이를 동반한 소아에서 프레드니솔론 반응성 도 좋았고, 미세잔유병소(minimal residual disease)도 적었고 장기간 생존율도 양호하였다. 이와 유사한 보고가 일본의 JACLS ALL-97 연구에서도 나왔다. 이들은 NOTCH1이나 FBXW7 돌 연변이를 동반한 소아 T-ALL 환자 55명을 분석하여 이들 돌연 변이가 동반된 경우 예후가 양호하였다고 보고 하였다[15]. 성 인에서도 NOTCH1 돌연변이가 동반된 T-ALL은 예후가 양호 한 것으로 보고 되었다[16]. 그러나 영국의 UKALLXII/ECOG E2993 연구는 NOTCH1이나 FBXW7 돌연변이는 환자의 예후 와는 무관하다고 결론을 내렸다[17]. 또한 소아 T-ALL 환자를 대상으로 연구한 DCOG/COALL 연구와 EORTC 58881 연구에 의하면 NOTCH1이나 FBXW7 돌연변이를 동반한 경우 초기 치료 반응성은 좋았으나 장기간 추적 조사에서는 이 요소들은 예후에 영향을 미치지 못 했다[18]. 이렇게 보고자마다 NOTCH1 이나 FBXW7 돌연변이를 동반한 T-ALL의 예후가 차이가 나는 것은 치료 방법의 차이 의해 나타나는 것으로 생각하고 있다.
NOTCH1의 돌연변이가 예후가 양호하다는 보고는 주로 강력 한 치료를 하였고 예후에 차이가 없었던 연구에서는 강력한 치료를 하지 않았다.
NOTCH1을 이용한 T-ALL의 표적치료제 개발
NOTCH1 돌연변이가 T-ALL에서 흔하게 동반되는 것의 중 요성은 NOTCH1의 작용을 방해하는 새로운 약물을 개발하여 치료에 응용할 수 있다는 점이다. 실제로 NOTCH1의 작용을 방해하는 약제로 NOTCH1이 활성화되는데 중요한 역할을 하 는 γ-secretase의 작용을 방해하는 γ-secretase inhibitor (GSI)가 치료제로 사용되는 시험이 있었다. GSI는 NOTCH 수용체 4
종류 모두에 작용하여 억제하는 약물로 실험실에서 T-ALL 세 포주에 투여할 경우 활성화된 NOTCH1 단백질은 신속하게 제 거되고 NOTCH1의 표적유전자들의 표현이 억제되었다. 또한 백혈병세포의 증식이 감소되는 고무적인 결과가 나왔다[19].
이러한 결과에 기초하여 임상시험이 미국의 Dana-Farber 암연
구소에서 GSI의 기능을 갖고 있는 MK-0752라는 경구용 약물
로 시행되었다. 대상 환자는 성인이 6명 소아가 2명으로 모두
8명으로 이중 NOTCH1 돌연변이를 동반한 T-ALL 환자는 4명
이었다. 임상시험 결과는 기대만큼 만족스럽지 못해 1명에서
만 반응이 있어 투여 28일 후 종격동종괴가 감소하는 효과가
있었다. 그러나 이후에는 T-ALL이 다시 진행되는 양상을 보였
다. 다른 7명의 환자 모두에서 이 약물의 부작용으로 약물 복
용을 중지하기 전에 반응을 모두 보이지 않았다[6]. 또한
MK-0752는 심각한 부작용을 초래하여 약물을 복용하는데 문
제가 있었다. 이 약물은 모든 환자에서 심각한 설사를 일으키
어 약물 복용을 장기간 할 수 없었다. 이러한 위장계 부작용을
GSI 약물이 동반하는 것은 예측할 수 없었던 일로 새로운 종
류의 약물을 개발하는데 걸림돌이 되었다. GSI는 정상상피의
증식과 분화에 중요한 역할을 하는 NOTCH1과 NOTCH2의
작용을 방해하여 심각한 위장관계의 부작용을 일으키는 것이
알려졌다. 이러한 부작용을 줄이고 항암 효과는 유지하는 여
러 치료법들이 연구되었다. 이러한 연구의 하나로 Merck 제
약회사는 GSI의 용량을 70%로 줄여 3일간 투여하였더니
T-ALL 이종이식 모델 실험에서 항백혈병 효과도 충분하였고
부작용도 줄일 수 있다고 보고하였다[20]. 비슷한 연구로
Pfizer사에서 개발한 GSI인 PF-0308414를 간헐적으로 투여하
였더니 부작용이 감소하였다. 이러한 연구들은 GSI는 투여 용
량과 투여 시간을 조절을 하면 부작용을 줄일 수 있다는 사실
을 시사한다[21]. 이와 더불어 부작용을 줄이고 치료 효과를
높이기 위한 노력으로 GSI를 다른 화학요법제제나 표적치료
제 또는 글루코코르티코이드 제제와 병행하는 치료에 대한 연
구가 있었다. 위에서 다룬 NOTCH1의 표적유전자들을 위주
로 한 치료법으로 CDK 방해제, NF-κB 신호전달을 표적으로
한 약물, PI3K-AKT-mTOR 경로를 표적으로 하는 약물 등과의
병행 치료는 백혈병 치료 효과를 증진시키는 것이 밝혀졌다
[22]. 주목할 만한 사실은 글루코코르티코이드를 GSI와 병행
투여할 때 부작용도 현저히 줄고 치료효과도 향상된다는 점이
다. T-ALL 세포주를 이용한 시험에서 GSI는 글루코코르티코
이드의 항백혈병 치료 효과를 증진시키며 또한 글루코코르티
코이드에 내성을 보이는 세포의 사멸을 유도하는 것이 알려졌
다. 또한 T-ALL 모형의 마우스를 이용한 생체 시험에서도 GSI
의 심각한 부작용인 위장관 장해가 없었다[23].
그러나 위에서 알아본 바와 같이 NOTCH1의 돌연변이는 T-ALL의 발생과 유지에 중요한 역할을 하는 것이 알려져 있지 만 간이 임상시험이었지만 GSI의 치료 효과는 만족스럽지 못 했다. 여러 연구에 의하면 T-ALL 세포주에 따라 GSI 투여 후 NOTCH1 신호가 확실하게 방해가 되어도 반응을 보이지 않는 저항성을 보이는 경우가 있다. 이러한 사항은 T-ALL에서 동반 되는 NOTCH1의 돌연변이 부위가 다양한 것과 관련이 있거나 T-ALL의 발생 및 유지에 NOTCH1 이외에 다른 이상이 관여하 는 것을 의심해 볼 수 있다. 실제로 T-ALL 세포주를 이용한 시험에서 GSI에 내성을 보이는 T-ALL 세포주는 암억제유전자 인 PTEN의 돌연변이를 동반하는 경우가 흔하였다[13]. 또한 FBXW7의 돌연변이는 N
ICD을 효과적으로 제거하지 못해 NOTCH1 신호를 증가시키나 한편으로 C-MYC, JUN, Cyclin E의 표현을 증가시키는데 이 경우 T-ALL은 GSI에 내성을 보 인다[24]. 이것은 NOTCH1 이외의 다른 경로가 T-ALL과 관련 이 있어 GSI에 내성을 보이는 것을 시사하고 있다.
GSI는 원래 알츠하이머병의 치료제로 이미 개발되어 있어 쉽게 구할 수 있는 약물이지만 NOTCH 수용체 4종류 모두에 비특이적으로 작용하여 T-ALL에서 동반되는 여러 종류의 돌 연변이 형태에 사용하는 데는 한계가 있다. T-ALL의 발생과 관련이 있는 NOTCH1을 표적으로 한 새로운 치료제가 현재 시험 중이다. 이런 약물로 N
ICD가 핵에서 자신의 표적유전자 의 표현을 유도를 돕는 조-활성자(co-activator)인 MAML을 표 적으로 하여 전사를 방해하는 SAHM1이 있다. 실험실 실험에 서도 SAHM1은 NOTCH1 돌연변이를 동반하고 있는 T-ALL 세 포주나 마우스에서 항백혈병 효과를 보여 주었다[25]. 또 다른 접근법으로 세포의 표면에 위치하는 NOTCH1 수용체에 특이 하게 작용하는 NOTCH1 항체를 치료제로 사용하는 방법이 다. 이러한 약물은 NOTCH1에만 특이하게 작용하므로 NOTCH2 억제에 의해 발생하는 위장관 부작용을 줄일 수 있 다. 실제로 NOTCH1 항체는 임상 전 시험에서 백혈병 세포의 성장을 억제하고 부수적으로 혈관생성도 억제하는 것으로 밝 혀졌다[26].
결 론
그 동안 B-ALL에 비해 분자의학적 이해가 미흡했던 T-ALL 에서 NOTCH1의 이상이 상당수에서 동반된다는 것이 알려졌 다. NOTCH1은 T 세포의 분화 과정에서 중요한 역할을 하는 신호전달 수용체로 이 분자의 이상이 T-ALL에서 발견된다는 점은 앞으로 T-ALL의 병태생리를 이해하는데 큰 도움이 될 것이다. 한편 T-ALL의 발생과 유지에 NOTCH1이 관여한다는
사실은 이 분자를 표적으로 하는 치료제를 개발할 수 있다는 점에서 중요하다. 현재 까지 NOTCH1을 표적으로 하는 약물 의 개발은 만족스럽지 못 하나 새로운 시도들이 진행되고 있 어 가까운 미래에 부작용이 없는 새로운 약물이 개발될 것으 로 생각된다. T-ALL은 아직도 치료가 용이하지 않은 질환으로 NOTCH1에 작용하는 새로운 약물이 개발되어 치료율의 향상 을 가져올 것으로 기대된다.
참 고 문 헌
1. Lee KC, Kang IJ, Kang SH, et al. Epidemiology and clinical characteristics of childhood acute lymphoblastic leukemia in Korea. Korean J Pediatr Hematol Oncol 2002;9:9-20.
2. Van Vlierberghe P, Ferrando A. The molecular basis of T cell acute lymphoblastic leukemia. J Clin Invest 2012;122:3398- 406.
3. Ellisen LW, Bird J, West DC, et al. TAN-1, the human homo- log of the Drosophila notch gene, is broken by chromosomal translocations in T lymphoblastic neoplasms. Cell 1991;66:
649-61.
4. Weng AP, Ferrando AA, Lee W, et al. Activating mutations of NOTCH1 in human T cell acute lymphoblastic leukemia.
Science 2004;306:269-71.
5. Demarest RM, Ratti F, Capobianco AJ. It's T-ALL about Notch.
Oncogene 2008;27:5082-91.
6. Paganin M, Ferrando A. Molecular pathogenesis and targeted therapies for NOTCH1-induced T-cell acute lymphoblastic leukemia. Blood Rev 2011;25:83-90.
7. Grabher C, von Boehmer H, Look AT. Notch 1 activation in the molecular pathogenesis of T-cell acute lymphoblastic leukaemia. Nat Rev Cancer 2006;6:347-59.
8. Beverly LJ, Capobianco AJ. Perturbation of Ikaros isoform se- lection by MLV integration is a cooperative event in Notch(IC)-induced T cell leukemogenesis. Cancer Cell 2003;3:
551-64.
9. Tanigaki K, Honjo T. Regulation of lymphocyte development by Notch signaling. Nat Immunol 2007;8:451-6.
10. Ferrando AA. The role of NOTCH1 signaling in T-ALL.
Hematology Am Soc Hematol Educ Program 2009:353-61.
11. Chiang MY, Xu L, Shestova O, et al. Leukemia-associated NOTCH1 alleles are weak tumor initiators but accelerate K-ras-initiated leukemia. J Clin Invest 2008;118:3181-94.
12. Tosello V, Ferrando AA. The NOTCH signaling pathway: role in the pathogenesis of T-cell acute lymphoblastic leukemia and implication for therapy. Ther Adv Hematol 2013;4:
199-210.
13. Palomero T, Sulis ML, Cortina M, et al. Mutational loss of PTEN induces resistance to NOTCH1 inhibition in T-cell leukemia. Nat Med 2007;13:1203-10.
14. Breit S, Stanulla M, Flohr T, et al. Activating NOTCH1 mutations
predict favorable early treatment response and long-term out- come in childhood precursor T-cell lymphoblastic leukemia.
Blood 2006;108:1151-7.
15. Park MJ, Taki T, Oda M, et al. FBXW7 and NOTCH1 muta- tions in childhood T cell acute lymphoblastic leukaemia and T cell non-Hodgkin lymphoma. Br J Haematol 2009;145:
198-206.
16. Asnafi V, Buzyn A, Le Noir S, et al. NOTCH1/FBXW7 muta- tion identifies a large subgroup with favorable outcome in adult T-cell acute lymphoblastic leukemia (T-ALL): a Group for Research on Adult Acute Lymphoblastic Leukemia (GRAALL) study. Blood 2009;113:3918-24.
17. Mansour MR, Sulis ML, Duke V, et al. Prognostic implications of NOTCH1 and FBXW7 mutations in adults with T-cell acute lymphoblastic leukemia treated on the MRC UKALLXII/ECOG E2993 protocol. J Clin Oncol 2009;27:4352-6.
18. Clappier E, Collette S, Grardel N, et al. NOTCH1 and FBXW7 mutations have a favorable impact on early response to treat- ment, but not on outcome, in children with T-cell acute lym- phoblastic leukemia (T-ALL) treated on EORTC trials 58881 and 58951. Leukemia 2010;24:2023-31.
19. Palomero T, Barnes KC, Real PJ, et al. CUTLL1, a novel hu- man T-cell lymphoma cell line with t(7;9) rearrangement, aberrant NOTCH1 activation and high sensitivity to gam-
ma-secretase inhibitors. Leukemia 2006;20:1279-87.
20. Tammam J, Ware C, Efferson C, et al. Down-regulation of the Notch pathway mediated by a gamma-secretase inhibitor in- duces anti-tumour effects in mouse models of T-cell leukaemia.
Br J Pharmacol 2009;158:1183-95.
21. Wei P, Walls M, Qiu M, et al. Evaluation of selective gam- ma-secretase inhibitor PF-03084014 for its antitumor efficacy and gastrointestinal safety to guide optimal clinical trial design. Mol Cancer Ther 2010;9:1618-28.
22. Palomero T, Ferrando A. Therapeutic targeting of NOTCH1 signaling in T-cell acute lymphoblastic leukemia. Clin Lymphoma Myeloma 2009;9 Suppl 3:S205-10.
23. Real PJ, Tosello V, Palomero T, et al. Gamma-secretase in- hibitors reverse glucocorticoid resistance in T cell acute lym- phoblastic leukemia. Nat Med 2009;15:50-8.
24. O'Neil J, Grim J, Strack P, et al. FBW7 mutations in leukemic cells mediate NOTCH pathway activation and resistance to gamma-secretase inhibitors. J Exp Med 2007;204:1813-24.
25. Moellering RE, Cornejo M, Davis TN, et al. Direct inhibition of the NOTCH transcription factor complex. Nature 2009;
462:182-8.
26. Wu Y, Cain-Hom C, Choy L, et al. Therapeutic antibody tar- geting of individual Notch receptors. Nature 2010;464:1052-7.