Several microorganisms directly or indirectly facilitate growth of plants and so called as “plant growth promoting rhizobacteria” (PGPR). Moreover, many reports have shown that PSB can promote plant growth, after their inoculation in soil or plant seeds for that reason (Datta et al., 1982; Gaur and Ostwal, 1972; Jones and Darrah, 1994; Kloepper et al., 1988). A plant growth promoting effect of PSB may also be related to their ability to synthesize plant growth regulating substances other than P solubilization (Azcon et al., 1978; Sattar and Gaur, 1987). There are several reports that the PSB were able to synthesize biologically active substances in liquid culture medium, which may stimulate the plant growth (Brown, 1972; Hussain and Vancura, 1970). Barea et al.
(1976) reported that PSB has synthesized 3 types of plant hormones, such as Indole-3-acetic acid (IAA), gibberellins and cytokinin-like substances. Benizri et al.
(1998) suggested that the production of IAA by P.
fluorescens increased maize growth and also Cheryl et al.
(2002) found that IAA produced by Psuedomonas putida significantly increased the its level in plant roots system.
Umali-Garcia et al. (1980) found that a tremendous stimulation in the growth of lateral roots and root hairs after inoculation with A. brasilense was attributed to production of IAA.
The aim of this study is to isolate PSB from the rhizosphere of crop and is to investigate the relationships among the phosphate solubilization activity, the production of organic acids and pH. Also, this study is to identify plant growth regulating substances from culture broth of PSB and to examine the effect of phosphate- solubilizing bacteria as inoculants on plant growth.
Materials and Methods
Isolation of rhizobacteria Rhizospheres were taken at a depth of 15-20 cm from crop field in Chonnam Province, Republic of Korea. Diluted soils by sterile water at a rate of 10-5 were inoculated on hydroxyapatite (HY) agar medium (glucose 10 g; MgSO
4· 7H
2O 0.4 g;
NaCl 1 g; CaCl
2· 2H
2O 0.2 g; NH
4NO
31.5 g; KCl 0.2 g;
yeast extract 0.5 g; hydroxyapatite 4 g; Agar 15 g per liter), and then incubated at 28 ℃ for 3 days. After incubation, a distinct single colony was picked-up and then routinely subcultured on the same medium until the purity of the strain. Such strains were stored at -70 ℃ for further study.
Identification of PSB The obtained strains were able to solubilize the insoluble phosphates on HY agar plate and were identified as phosphate solubilizing bacteria (PSB) by 16S rRNA analysis. A large fragment of 16S rRNA gene was amplified by polymerase chain reaction (PCR) using the forward primer Y1 (5'-tggctcagaacgaacgctggcggc-3') and the reverse primer Y2 (5'-cccactgctggctcccgtaggagt-3').
The PCR temperature cycle was at 94 ℃ for 30 seconds, 55 ℃ for 1 minute, and 72 ℃ for 1 minute 30 seconds for 30 cycles and 5 minutes at 72 ℃ for extension. The PCR products were purified by Accu Prep Gel purification kit (Bioneer Co., Chungbuk, Korea). The nucleotide sequence of the 16S rRNA gene was determined by Biodye Terminator cycle sequencing kit and compared with published 16S rRNA sequences using Blast search at Gene Bank Data base of NCBI (Bethesda, MD) in order to establish the identity of the isolates.
Determination of P solubilization, organic acid production and pH PSB were grown on 200 ml of HY broth at 30 ℃ for 10 days under the air supply filtered through a 2 μ m Whatman membrane at 30 ml min
-1on a rotary shaker at 200 rpm. Samples were taken at 1, 2, 5, 7 and 10 days for the measurement of pH, P solubilization and organic acid concentration. All these experiments were carried out in triplicate. The pH of broth culture was measured by using pH meter (HANNA-HI 8314, Italy).
For analysis of phosphate concentration, broth culture of each treatment was filtered through a Whatman 0.2 μ m membrane filter. The 50 μ l of filtrated culture was mixed with 1.95 ml extraction solution (0.025 N HCl and 0.03 N NH
4F) and then 2 ml of this solution, 5 ml of distilled water, 2 ml of ammonium paramolybdate solution (12 mM), and 1 ml of SnCl2 solution (5 mM) were mixed well. After 5 min incubation at room temperature (RT), P
2O
5concentrations were measured by a spectrophotometer at 660 nm (Olsen and Sommers, 1982). The productions of organic acid by PSB culture filtrates were analyzed by high performance liquid chromatography (HPLC) with a Shodex RSpak KC-811 column (8 mm × 30 cm) and monitored by UV detector at 210 nm. The operating conditions consisted of 0.1%
phosphoric acid solution as the mobile phase, a flow rate
of 0.5 ml min
-1and sample injection of 20 μ l. The
productions of organic acids were quantitatively
determined by comparing the retention times and peak
areas of chromatograms with those of standards.
Extraction and identification of IAA Production of indole-3-acetic acid (IAA) from the culture filtrates of PSB was done by a modified method of Lindow et al.
(1998). PSB were grown on the medium (0.5M of glucose; 0.4g of MgSO
4· 2H
2O; 1g of NaCl; 0.2g of CaCl
2· 2H
2O, 1.5 g of NH
4NO
3; 2 g of Hydroxy apatite; 0.2g of KCl; 0.15g of Yeast extract; 0.15g of Yeast autolyzate; 0.15g of Trypton pepton, per 1L ; pH 7.0) at 30 ℃ for 3 days under a rotary shaker at 200 rpm until the stationary stage was reached. Then the culture broth was centrifuged at 2,000 rpm for 20 minutes. The supernatants were adjusted with 1N HCl to pH 3 and then partitioned three times with equal volume of ethyl acetate. The organic phases were combined with dehydrates and anhydrous Na
2SO
4. The solvent was evaporated by a rotary evaporator (BUCHI, RotavaporR-114, Germany) and then the residues were dissolved in methanol. HPLC analysis was done by injecting 10 μ l of the purified sample onto a Waters reverse-phase C
18column (7 μ m, 4 mm x 30 cm) and elutes were detected at 220 nm using a UV detector.
Mobile phase was acetonitrile : 50 mM KH
2PO
4(30:70) at flow rate of 1 ml min
-1. The retention times for the peaks obtained in IAA producer were compared with those of authentic IAA and quantity was done by a comparison of peak heights.
Enhancement of plant growth promotion by PSB Cucumber seeds (Sinhuk, Hungnong Seed Co, Republic of Korea) were sterilized with a 2% sodium hypochlorite solution to eliminate seed-infesting organisms and then sowed in sterilized soil mixture (soil:vermiculite:sand, 2:1:1, v/v/v) containing 1% hydroxyapatite. The growth mixture had a pH (1:5, H
2O) of 6.3, total N of 0.1%, organic matter of 1.0%, available P
2O
5of 72 ppm, and CEC of 5 cmol
+kg
-1. Each pot was filled with 500 g of the mixture where a sterilized seed was placed. Five milliliters of each bacterial culture suspension (P.
orientalis, E. asburiae, and Acinetobacter sp., 4 × 10
7CFU ml
-1) was amended to the rhizosphere of each plant in each treatment at 5 and 6 weeks after sowing and control plants received the same amount of culture broth (without bacterial inoculation). After 7 weeks, cucumber plants were harvested and fresh/dry weight of shoot and root, and N and P content in plant were measured in triplicate. Throughout the experiment, the plants were grown at 25-35 and 60-70% relative humidity.
Results
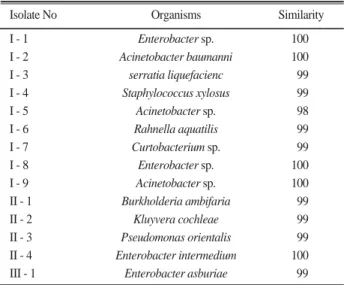
Isolation and identification of bacteria Crop- associated rhizobacteria were isolated from Chonnam Province in Republic of Korea. A total of 14 bacteria were isolated as phosphate solubilizing bacteria (PSB) having P solubilizing ability on HY medium. Among them, 3 strains showing strong P solubilizing ability were selected and then identified as Acinetobacter sp. (I-5), Pseudomonas orientalis (II-3), and Enterobacter asburiae (III-1) based on their morphology and 16S rRNA analysis (Table 1).
Determination of organic acid production Organic acid produced from the culture filtrate of Acinetobacter sp., P. orientalis, and E. asburiae was carried out at 1, 2, 5, 7 and 10 days after cultivation by HPLC analysis. The HPLC analysis of these culture filtrate revealed that four major peaks such as 1, 2, 3 and 4 were represented as
Isolate No
I - 1 Enterobacter sp. 100
I - 2 Acinetobacter baumanni 100
I - 3 serratia liquefacienc 99
I - 4 Staphylococcus xylosus 99
I - 5 Acinetobacter sp. 98
I - 6 Rahnella aquatilis 99
I - 7 Curtobacterium sp. 99
I - 8 Enterobacter sp. 100
I - 9 Acinetobacter sp. 100
II - 1 Burkholderia ambifaria 99
II - 2 Kluyvera cochleae 99
II - 3 Pseudomonas orientalis 99
II - 4 Enterobacter intermedium 100
III - 1 Enterobacter asburiae 99
Organisms Similarity
Table 1. Identification of PSB by 16S rRNA sequence analysis.
Fig. 1. Typical HPLC chromatograms of organic acids produced by PSB on HY broth at 30℃ ℃ for 3 days. Peaks 1, 2, 3 and 4 represent gluconic acid, 2-ketogluconic acid, succinic acid and acetic acid, respectively. A: Acinetobacter sp., B:
Pseudomonas orientalis, and C: Enterobacter asburiae.
¨
gluconic acid, 2-ketogluconic acid, succinic acid, and acetic acid,?respectively by comparing the retention times with those of the authentic standards (Fig. 1). Strain Acinetobacter sp. released only gluconic acid in the culture filtrate and strain E. asburiae released both of acetic acid and succinic acid. Interestingly, P. orientalis produced gluconic acid at 1
stday, and then it was converted into 2-ketogluconic acid rapidly.
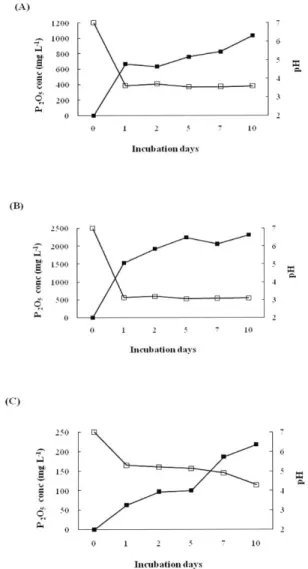
Relationships among pH, organic acid production and P solubilization In order to study the relationship between the production of organic acids and P solubilization, we incubated Acinetobacter sp., P.
orientalis, and E. asburiae on HY broth medium with continuous air supply at pH 7.0 for 10 days. The pH of the media rapidly decreased in 1
stdays and then slowly decreased till 10 days of incubation (Fig. 2). However,
soluble P concentration continuously increased during the incubation periods. The amount of P released in culture filtrate was detected at 1030 mg L
-1(Acinetobacter sp.), 2312 mg L
-1(P. orientalis) and 218 mg L
-1(E. asburiae) after 10 days of inoculation (Fig. 2). Moreover, the rate of acid production significantly increased during the growth periods of Acinetobacter sp., P. orientalis and E.
asburiae. As showed in Fig. 3, strain Acinetobacter sp.
produced gluconic acid with concentration about 11,890 mg L
-1and P. orientalis released 2-ketogluconic acid with concentration about 10,524 mg L
-1accompanied by a small amount of gluconic acid. Also, E. asburiae released both of succinic acid and acetic acid, separately 1100 mg L
-1and 1222 mg L
-1.
Fig. 2. Changes in pH and phosphorus concentration by PSB on HY broth culture at 30℃ ℃ for 10 days. A: Acinetobacter sp., B: Pseudomonas orientalis and C: Enterobacter asburiae.
■: P
2O
5concentration □: pH
Fig. 3. Changes in concentration of organic acids by PSB on HY broth culture at 30℃ ℃ for 10 days. A: Acinetobacter sp., B:
Pseudomonas orientalis, and C: Enterobacter asburiae.
□: gluconic acid, ■: 2-ketogluconic acid, □: succinic acid, ●:
acetic acid.
Determination of IAA production PSB were assessed for their role in plant growth promoting ability of cucumber plants. The production of growth regulating substance was analyzed by HPLC in the culture filtrate of Acinetobacter sp., P. orientalis, and E. asburiae. The presence of IAA in the culture fluids of P. orientalis had 372 mg L
-1and E. asburiae produced about 191 mg L
-1, while Acinetobacter sp. did not produce a detectable amount of IAA during experiment period (Table 2). In further, presence of IAA in the culture fluids of P.
orientalis, and E. asburiae had the major peak coincided with that of authentic IAA (data not shown).
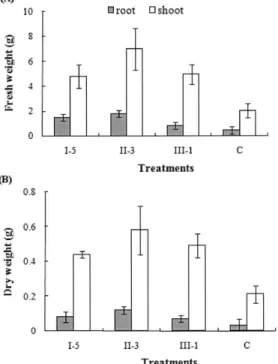
Enhancement of plant growth promotion After 7 weeks of planting, fresh and dry weights of root and shoot were measured. Interestingly, P. orientalis had
highly increased fresh and dry weights of root and shoot, when compared with control (Fig. 4). This data generally showed the plant growth promotion ability by phosphate solubilizing bacteria with the production of IAA.
Furthermore, P
2O
5and total nitrogen (T-N) of plants were highest in P. orientalis treatment, followed by E.
asburiae, Acinetobacter sp., and control (Figs. 5 and 6).
Discussion
Rhizosphere is a natural harbor for phosphate solubilizing bacteria (PSB), which can solubilize the insoluble inorganic P into soluble form in the crop fields.
Many agricultural microbiologists and microbial ecologist have been interested in the ability of these bacteria to dissolve insoluble phosphates such as hydroxyapatite and rock phosphate (Goldstein et al.,
Organism
"
nd ; not detected
Acinetobacter sp. nd
"Pseudomonas orientalis 372
Enterobacter asburiae 191
Concentration of IAA mg L
-1Table 2. Detection of IAA from the culture filtrates of PSB.
The bacteria were grown at 30℃ ℃ for 5 days on HY medium containing 0.4% hydroxyapatite.
Fig. 5. Content of phosphorus in root and shoot of cucumber at 7 weeks after planting as influenced by inoculation with Acinetobacter sp. (I-5), Pseudomonas orientalis (II-3), Enterobacter asburiae (III-1) and control.
Fig. 6. Content of nitrogen in root and shoot of cucumber at 7 weeks after planting as influenced by inoculation with Acinetobacter sp. (I-5), Pseudomonas orientalis (II-3), Enterobacter asburiae (III-1), and control.
Fig. 4. Fresh(A) and dry(B) weights of root and shoot of
cucumber plants at 7 weeks after planting as influenced by
inoculation with Acinetobacter sp. (I-5), Pseudomonas orientalis
(II-3), Enterobacter asburiae (III-1) and control (C).
1999). In addition to, this solubilization activity by PSB is mainly derived from the production of organic acids (Halder et al., 1991; Illmer et al., 1995). Thus, we established a method for assembling and characterizing of rhizobacteria from crop rhizospheres. Also, we have demonstrated that among isolated PSB, Acinetobacter sp., Pseudomonas orientalis, and Enterobacter asburiae did show strong P solubilization activity and they employed the similar mechanism of P solubilization.
Indeed, PSB produced various organic acids in the culture medium, such as gluconic acid, 2-ketogluconic acid, succinic acid, and acetic acid by Acinetobacter sp., P. orientalis, and E. asburiae (Fig. 1). During the growth period of these PSB, the soluble P concentration was inversely correlated with pH in culture medium, which was concomitant with increased organic acid concentrations. Interestingly, the rate of P solubilization by P. orientalis showed as two fold increased, when compared to Acinetobacter sp. However, P. orientalis was found to produce 2-ketogluconic acid, which was almost same concentration of gluconic acid produced by Acinetobacter sp (Figs. 2 and 3). It was known that 2- ketogluconic acid has strong ionic strength (pKa = 2.66), which was 100 times higher than citric acid (pKa = 4.74) (Moghimi and Tate, 1978). The ionized proton ions should easily solubilize rock P into soluble forms (Hwangbo et al., 2003; Kim et al., 1997b).
P solubilization is a well known mechanism that involved in growth promotion of crops by PSB (Hilda and Reynaldo, 1999). P solubilizer showed a great ability to produce growth regulators and was frequently found in the rhizosphere (Vandana and Reeta, 2003). Growth promotion in cucumber caused by PGPR treatment may be due to the production of phytohormones. In this study, we have been able to identify IAA from the culture filtrate of PSB. Treatment of strain P. orientalis did cause increased root (2-3 folds) and shoot (1-2 folds) weights of cucumber seedling compared with untreated control (Fig.
4). This result was same that the inoculation with PSB in soils can increase crop yields by growth promoting regulators of PSB (Freitas et al., 1997). This investigation suggested that the strain P. orientalis was an efficient candidate for plant growth promoting function by the instance of IAA production. Moreover, P
2O
5and total nitrogen (T-N) of P. orientalis had significantly increased, when compared with others and control (Figs. 5 and 6).
Through all these experiments, the importance of P solubilization as the mechanism that mediates the
solubilization of insoluble P was established. Moreover, the production of IAA appeared to major factors that may play a vital role in the PGPR activity and also contribute to the growth promotion in cucumber plant.
Choice of useful PSB candidates is indeed one of the most important factors for the success of P solubilization programs on a commercial basis. It is therefore important to consider cost effective, eco-friendly and environment management. This could be further exploited or accelerated to the advantage of crop farmers by developing superior PSB strains as microbial inoculants/biofertilizer for crop productivity.
Acknowledgements
This study was supported by the Korea Research Foundation second step Brain Korea 21 project, Republic of Korea.
References
Azcon, R., G. Azcon, D. Aghilar, and J.H. Barea. 1978. Effect of plant hormones present in bacterial cultures on the formation and response to vamendomycorrhiza. New Phytol. 80:359-364.
Banik, S., and B.K. Dey. 1982. Available phosphate content of and alluvial soil as influenced by inoculation of some isolated phosphate solubilizing bacteria. Plant Soil. 69:353-364.
Barea, J.M., E. Navarro and E. Montoya. 1976. Production of plant growth regulators by rhizosphere phosphate-solubilizing bacteria.
J. Appl. Bact. 40:129-134.
Benizri, E., A. Courtade, C. Picard and A. Guckert. 1998. Role of maize exudates in the production of auxins by Pseudomonas Fluorescens M.3.1. Soil Biol. Biochem. 30:1481-1484.
Brown, M.E. 1972. Plant growth substances produced by micro- organisms of soil and rhizosphere. J. Appl. Bact. 35:443-451.
Cheryl, L.P., and R.G. Bernard. 2002. Role of Pseudomonas putida indoleacetic acid in development of the host plant root system.
Appl. Environ. Microbiol. 68:3795-3801.
Datta, M., S. Banik and R.K. Gupta. 1982. Studies on the efficacy of a phytohormone producing phosphate solubilizing Bacillus firmus in augmenting paddy yield in acid soils of Nagaland. Plant and Soil 69:365-373.
Freitas, J.R., M.R. Banerjee and J.J. Germida. 1997. Phosphate- solubilizing rhizobacteria enhance the growth and yield but not phosphorus uptake of canola (Brassica nupus L.). Biol. Fertil.
Soils. 24:358-364.
Gaur, A.C., and K.P. Ostwal. 1972. Influence of phosphate dissolving Bacilli on yield and phosphate uptake of wheat crop.
Indian J. Exp. Biol. 10:393-394.
Goldstein, A.H. 1986. Bacterial mineral phosphate solubilization:
Historical perspective and future prospects. Am. J. Altern. Agric.
1:57-65.
Goldstein, A.H., K. Braverman and N. Osorio. 1999. Evidence for mutualism between a plant growing in a phosphate-limited desert environment and a mineral phosphate solubilizing (MPS) rhizobacterium. FEMS Microbiol. Ecol. 30:295-300.
Halder, A.K., A.K. Misra and P.K. Chakrabarty. 1991.
Solubilization of inorganic phosphates by Bradyrhizobium. Indian J. Exp. Biol. 29:28-31.
Halder, A.K., and P.K. Chakrabartty. 1993. Solubilization of inorganic phosphate by Rhizobium. Folia Microbiol. 38:325-330.
Hilda, R., and F. Reynaldo. 1999. Phosphate solubilizing bacteria and their role in plant growth promotion. Biotechnol. Adv.17:319- 339.
Hu, H.Q., J.Z. He, X.Y. Li, and F. Liu. 2001. Effect of several organic acids on phosphate adsorption by variable charge soils of central China. Environ. Int. 26:353-358.
Hussain, A., and V. Vancura. 1970. Formation of biologically active substances by rhizosphere bactieria and their effect on plant growth. Folia Microbiol. 15:468-478.
Hwangbo, H., D.R. Park, W.Y. Kim, Y.S. Rim, H.K. Park, H.T.
Kim, J. SunSuh and Y.K. Kim. 2003. 2-Ketogluconic acid production and phosphate solubilization by Enterobacter intermedium. Curr. Microbiol. 47:87-92.
Illmer, P., and F. Schimmer. 1992. Solubilization of inorganic phosphates by microorganisms isolated from forest soil. Soil Biol.
Biochem. 24:389-395.
Illmer, P., A. Barbato, and F. Schinner. 1995. Solubilization of hardly-soluble AlPO
4with P-solubilizing microorganisms. Soil Biol. Biochem. 27:265-270.
Jones, D.L., and P.R. Darrah. 1994. Role of root derived organic acids in the mobilization of nutrients from the rhizosphere. Plant and Soil 166:247-257.
Kim, K.Y., D. Jordan and H.B. Krishnan. 1997a. Rahnella aquatilis, a bacterium isolated from soybean rhizosphere can solubilize hydroxyapatite. FEMS Microbiol. Lett. 153:273-277.
Kim, K.Y., G.A. McDonald and D. Jordan. 1997b. Solubilization of hydroxyapatite by Enterobacter agglomerans and cloned Escherichia coil in culture medium. Biol. Fertil. Soils. 24:347-352.
Kloepper, J.W., K. Lifshitz and M.N. Schroth. 1988. Pseudomonas inoculants to benefit plant production. ISI Atlas Sci. Anim. Plant.
60-64.
Lindow, S.E., C. Desurmont, R. Elkins, G. McGourty, E. Clark and M.T. Brandl. 1998. Occurrence of indole-3-acetic acid producing bacteria on pear trees and their association with fruit russet.
Phytopathology 88:1149-1157.
Liu, S.T., L.Y. Lee, C.Y. Tai, C.H. Horng, Y.S. Chang, J.H.
Wolfram, R. Rogers and A.H. Goldstein. 1992. Cloning of and Erwinia herbicola gene necessary for gluconic acid production and enhanced mineral phosphate solubilization in Escherichia coli HB101. J. Bacterial. 174:5814-5819.
Moghimi, A., and M.E. Tate. 1978. Does 2-ketogluconate chelate calcium in the pH range 2.4 to 6.4. Soil Biol. Biochem. 10:289- 292.
Satter, M.A., and A.C. Gaur. 1987. Production of auxins and gibberellins by phosphate dissolving microorganisms. Zentralbl.
Microbiol. 142:393-395.
Shekhar, N.C., S. Bhadauria, P. Kumar, H. Lal, R. Mondal and D.
Verma. 2000. Stress induced phosphate solubilization in bacteria isolated from alkaline soils. FEMS Microbil. Lett. 182:291-296.
Umali-Garcia, M., D.H. Hubbell., M.H. Gaskins., F.B. Dazzo. 1980.
Association of Azospirillum with grass roots. Appl. Environ.
Microbiol. 39:219-226.
Vandana, K., and G. Reeta. 2003. Solubilization of inorganic phosphate and plant growth promotion by cold tolerant mutants of Pseudomonas fluorescens. Microbiol. Res. 158:163-168.
Olsen, S.R., and L.E. Sommers. 1982. Phosphorus. p. 403-430. In A.L. Page et al. (ed.), Method of soil analysis, Chemical and microbial properties. American Society of Agronomy, Madison, WI, U.S.A.
Yadav, K.S., and K.R. Dadarwal. 1997. Phosphate solubilization and mobilization through soil microorganisms. p. 293-308. In K.R.
Dadarwal, (ed.), Biotechnological approaches in soil
microorganisms for sustainable crop production. Scientific
Publishers, Jodhpur, India.
14종의 인산용해미생물을 근권으로부터 분하였고, 16S rRNA gene 염기서열에 의하여 동정하였다 그 중 hydroxyapatite를 첨가한 배지에서 인산용해능력이 가장 뛰어난 Acinetobacter sp., Pseudomonas orientalis, Enterobacter asburiae 3종을 선택하였다. 선택된 3종의 미생물에 의해 용해된 인산의 농도는 200 mg L
-1에서부 터 2300 mg L
-1까지 이르렀으며, 증가된 인산 농도는 배양액의 pH와 역으로 비례하였다. HPLC를 사용하여 유 기산을 측정한 결과 Acinetobacter sp.는 gluconic acid를, P. orientalis는 gluconic acid와 2-ketogluconic acid를 그 리고 E. asburiae는 acetic acid와 succinic acid를 분비하였다. 한편 P. orientalis와 E. asburiae는 각각 372 mg L
-1와 191 mg L
-1의 IAA 분비하였고, Acinetobacter sp.는 IAA를 생성하지 못했다. 인산용해미생물이 오이의 생장 에 미치는 효과를 조사한 결과, P. orientalis를 처리한 시험구가 가장 높았고, E. asburiae, Acinetobacter sp., control 순으로 나타났다. 이러한 식물생장 효과는 인산용해미생물에 의한 불용성 인산용해와 IAA 생산과 관련 이 있다고 생각된다.
인산용해미생물에 의한 불용성 인의 용해와 식물생장에 미치는 영향
나정행·최진호1·김영덕·고현선·박노동·김길용*