400
Copyright © 2019 The Korean Society of Fisheries and Aquatic Science pISSN:0374-8111, eISSN:2287-8815
서 론
돌기해삼
(Apostichopus japonicus)
은주로우리나라의모든 연안,
중국랴오닝성의대련연해,
일본홋가이도연안그리고 러시아오호츠크해역까지북동태평양한대전역에분포하고 있으며,
한국과중국그리고일본에서중요한수산양식생물이 다(Slater and Carton, 2007; Jin et al., 2016a).
해삼은예로부터 바다의인삼으로불리어지고있으며,
최근소비가확대되면서중국을중심으로새로운양식대상종으로각광받고있다
(Jin
et al., 2016b).
특히중국에서해삼을먹는음식문화가발달되 어있어전세계약90%
의해삼을소비하고있다(Wen et al., 2016; Shahabuddin et al., 2017).
해삼공급량이수요를충족시 키지못하고있으며,
이러한수요공급의불균형은중국해삼양 식뿐만아니라일본과우리나라의해삼양식을유발시키는촉진제가되었다
(Yu et al., 2014).
우리나라에서해삼은마을어 장의대표적인어획대상종이며,
생산량이1990
년에는2,491
톤이나달하였으나연안어장의환경변화,
해안매립등으로산란장과서식장이축소됨에따라
2002
년에는833
톤까지격감하였다
.
이에2000
년대중반부터인공종자생산기술이개발되었고
,
해삼자원을효율적으로이용하고자원을증강하기위하여 국가와지자체의노력으로2017
년에는2,045
톤으로빠르게증 가하였지만,
그수요에비해생산량은턱없이부족한상태이다(Anisuzzaman et al., 2018).
이를극복하기위해서는해삼양식 산업이규모의산업으로의전환이필요하며생산에서가공으로 연결되는물량의안정적인대량확보가선결과제이다.
현재생 산된종자는연안어촌계의자원증강을위한씨뿌림양식용으로 사용되고있으며,
서해안의대규모연안간석지를이용하여대 량생산이가능한축제식양식용으로는종자가격등의문제로고수온에서의 해삼(Apostichopus japonicus) 성장과 체지방 조성
정우철·Anisuzzaman Md·Feng Jin·최종국·한종철
1·최병대·강석중*
경상대학교 해양식품생명의학과, 1국립수산과학원 남동해수산연구소
Effect of High Water Temperature on the Growth and Lipid Compositions of the Sea Cucumber Apostichopus japonicus
U-Cheol Jeong, Anisuzzaman Md, Feng Jin, Jong-Kuk Choi, Jong-Cheol Han
1
, Byong-Dae Choi and Seok-Joong Kang*Department of Seafood and Aquaculture Science, Gyeongsang National University, Tongyeong 53064, Korea
1Southeast Sea Fisheries Research Institute, National Fisheries Research and Development Institute, Tongyeong 52440, Korea
The optimum growth temperature of sea cucumber is 18°C, above 20°C, the appetite declines and the digestive tract is atrophied. At 25°C or more, it is known that the digestive tract is degenerated as growth is stopped by minimizing metabolic activity. Because of these physiological and ecological characteristics, the growth period of sea cucumber Apostichopus japonicus is relatively short in the natural environment of Korea where four seasons are clear. There- fore, maintenance of water temperature in sea cucumber culture is very important. This experiment was conducted to investigate the growth and lipid compositions of sea cucumbers at high temperature. Results showed that the growth and survival rates of sea cucumber were not significantly different until 26°C (P>0.01). The fatty acid compositions of total lipid of sea cucumber tended to increase with saturated fatty acid content and decrease with n-3 fatty acid content at higher water temperature.
Key words: Sea cucumber, Apostichopus japonicus, Recirculating aquaculture system, High water temperature, Growth
*Corresponding author: Tel: +82. 55. 772. 9154 Fax: +82. 55. 648. 3089 E-mail address: [email protected]
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial Licens (http://creativecommons.org/licenses/by-nc/3.0/) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
Received 1 July 2019; Revised 22 July 2019; Accepted 24 July 2019
저자 직위: 정우철(연구교수), Anisuzzaman Md(연구원), Feng Jin(연구원), 최종국(연구원), 한종철(연구사), 최병대(교수), 강석중(교수)
https://doi.org/10.5657/KFAS.2019.0400
Korean J Fish Aquat Sci 52(4), 400-407, August 2019
고수온에서의 해삼 성장과 체지방 조성
401
인하여연결되지못하고있다
(Kang, 2012).
돌기해삼은극피동물문해삼강에속하는무척추동물로서기 존의양식생물과는달리양식생태학적으로복잡한유생생활과 하면
(夏眠)
을하는생태적특징때문에양식기술개발에많은 어려움이있다.
수정난에서아우리쿨라리아,
돌리올라리아,
펜 탁툴라,
착저유생및어린유생시기을거치는복잡한유생단 계와그단계별먹이생물의종류와급여방법은기초생물학적 기술개발에힘입어어느정도의알고리즘이정립된상태이다(Kang, 2012).
그러나,
수온생태에관해서는겨울철에는동면 하고여름철에는하면한다는자연상태에서일어나는일반생태 학적인관점에머물러있다.
지금까지수온과관련된생태학적 지식으로최적성장수온은10-18°C
이며(Li et al., 2002; Dong et al., 2006),
수온이낮은겨울철에는동면을하고,
수온이25°C
이상되는시기에는대사활동을최소화시켜성장을멈추거나,
소화관이퇴화되어하면하는것으로알려져있으며
(Liu et al.,
1996; Yang et al., 2006; Ji et al., 2008),
해삼의이런생태적인 특성때문에사계절이뚜렷한우리나라의자연환경에서는해 삼의성장기간이짧아해삼양성이불리한것으로인식되고있 다(Jeong et al., 2016a).
그러나현재인공종자생산현장에서는 양식장마다각기다른수온범위에서양성을하고있으며, 30°C
의고수온에서도섭이활동및생존하며그결과도다르게나타 나고있기때문에해삼인공종자생산의최적수온에대하여혼 란이생기고있다.
해삼종자생산에있어서수온관리는무엇보다도중요하고
,
특히수온변화에따라서동면과하면을겸하는해삼의생리적 특성을고려하면수온에대한이해는매우중요하다.
또한변온 성동물인해삼은수온변화에따른세포내체액항상성유지 를위해지방산의변화에도영향을미치는것으로알려져있다(Fredalina et al., 1999).
따라서본연구는해삼종자생산현장에서일어나고있는암묵지
(暗默智)
를객관화시키기위하여고수온에서어린해삼사육을디자인하였으며
,
수온에따른체지 방의조성변화도함께알아보고자하였다.
재료 및 방법
실험해삼
실험에사용한해삼
(Apostichopus japonicus)
은경남고성군 삼산면소재의해삼종자배양장에서양식중이던어미해삼으 로부터산란시켜서유생시기를거쳐어린해삼까지자체사육 한크기가비슷한해삼1,000
마리를분양받아사용하였다.
실 험실로운반된해삼은penicillin
계열의polymycin
을사용하여30 ppm
에서20
분간약욕처리후사용하였다.
실험해삼은실 험시작전3
주일동안실험수조에서예비사육을실시하였으 며,
이때실험에사용한해삼의평균습중량은4.60±0.08 g
을사용하여15
마리씩사용하였으며,
모든실험구는3
반복으 로하였다.
실험사료
실험에사용한사료는해삼종묘배양장에서사용중인분말 상품사료
(Qingdao hiford ecology technology Inc., China)
를 사용하였다.
사료의성분조성은수분10.0%,
조단백질23.0%,
조지방2.0%,
탄수화물10.0%
그리고조회분55.0%
였다.
사료공급량은해삼평균중량의
3-5%
를기준으로매일저녁8
시경
1
회/
일공급을하였으며,
사료를공급할때사료가수중에 퍼져서발생하는수질의혼탁을막기위해서유리관피펫을사 용하여수조바닥층에깔아주는방법으로공급하였다.
사육수 조내배설물및사료찌꺼기제거는매일오후6
시에사이펀으 로제거하였다.
사육관리
실험에 사용된 사육장치는 순환여과시스템을 이용하였다
.
전체시스템의수량은12,000 L
로순환펌프(PH-350M, Wilo
Inc., Korea)
를이용하여시스템전체순환율은30
회전/
일이었 으며,
수온조절은냉각기(DA-3000W, Dae-il Inc., Korea)
와티 타늄히터(2.0KW, Sewon Inc., Korea)
를 이용하여 사육수온 을18.0±0.5°C
로조절하였다.
사육에사용한수조는유효수량 은50 L
로직사각형아크릴재질로서크기는60 (length)×30
(width)×35 (height) cm
이고,
사육수조바닥은6%
의경사도 를가지고있으며배출구반대편의주입수에의하여해삼변이 배출구쪽으로모이도록하였다.
배출구에는PVC
파이프(
직 경16 m/m)
를설치하여무결망(
망목사이즈, 3 mm)
으로겉면 을둘러싸서사용하였다.
침전조는원추형FRP
원형수조(
직경150 cm×
높이130 cm,
유효수량1,000 L) 2
개를직열방식으로 사용하였으며바닥은2%
의경사로수집된중앙에집적된고형 물인뻘과잔류된사료는매일한번씩제거하였다.
생물학적여 과조는제1
여과조와제2
여과조2
개를설치하였다.
제1
여과조 는여과재를굴패각(
좌각과우각의비율1:1)
을사용하였으며,
수류의흐름은역여과방식으로하였고,
제2
여과조의여과재는 선라이트골판(65×60 cm) 800
여장를교차시켜서사용하였으 며,
총여과재의표면적(biofitler media surface)
은624 m
2였고,
수류의흐름은정여과방식으로하였다.
수위조정용탱크는여 과조에서여과된사육수,
헤드탱크에서남은여분의사육수와 보충된해수를받아들여서수위를자동으로조절하게하였다.
보충수는 증발,
누수,
고형오물의 청소목적으로 전체수량의7-10%
를보충하였다.
사육수조의 수온조절은히터(HB-100,
Periha Inc., China)
를이용하여각각18°C, 22°C, 26°C
그리고30°C
로하였으며,
사육조순환율은5-10
회전/
일이었다.
수온과pH
는pH meter (PH-20N, Istek Inc., Korea),
용존산소는DO
meter (YSI-58, YSI, USA)
그리고염분농도는EC meter (CM-
21P, Japan)
를사용하여1
회/
일측정하였다. TAN (total ammo-
nia nitrogen)
는ammonia meter (Palintest, Nessler, England)
로측정하였으며, TSS (total suspended solid)
는진공여과법으 로2
회/
주측정하였다.
성장도 측정
성장은
4
주간격으로12
주간측정하였으며,
측정전일은절식 시킨후,
각수조의해삼을수집하여전개체의습중량을측정 하여증체율(weight gain, %),
일간성장률(specific growth rate,
%/day)
그리고생존율(survival rate, %)
을산정하였으며,
계산 식은다음과같다.
Weight gain WG (g)=W
2-W
1Specific growth rate (SGR) (%/d)=[(lnW
2-lnW
1)/T]×100 Survival rate (SR) (%)=(N
2/N
1)×100
위식에서
W
1은초기생체량, W
2는최종생체량, T
는사육기 간, N
1은최초개체수, N
2는최종생존개체수이다.
일반성분 분석
일반성분은
AOAC (2000)
방법에따라수분은상압가열건 조법,
조단백질은Kjeldahl
법,
조회분은직접회화법으로측정 하였으며,
조지방은choloroform
과methanol
을2:1
비율로혼 합한용액을용매로한Bligh and Dyer (1959)
추출법에준하였 다.
수분은상압가열건조법으로10°C
의건조기(Dongwon, dry oven, Korea)
에서6
시간동안건조후측정하였으며,
조회분은 직접회화법으로60°C
회화로에서4
시간동안태운후정량하였 다.
탄수화물함량은시료의수분,
단백질,
지질및회분의함량 을뺀값으로하였다.
Total Lipid 추출
해삼시료약
50 g
에혼합추출용매(chloroform:methanol=
2:1, v/v)
를시료의2
배량넣고,
세포분쇄기(Homogenizer AM- 12, Nihonseiki Kaisha Co. Ltd., Tokyo, Japan)
로분쇄(15,000 rpm, 5
분)
한다음암소에서하루동안방치하였다.
이를여과 한다음잔사물은버리고,
여액(
추출용매와지방혼합물)
을분 액깔때기에넣어,
층분리가일어나면아래층(chloroform
층)
을제습제(Na
2SO
4)
를통해탈수시켜받은다음진공회전농축 기(Rotavapor R-114, BUCHI, Swiss)
로40°C
이하에서추출용 매를완전히증발시킨후총지질을얻었다.
지방산조성 분석
추출된총 지질
100 mg
을cap tube
에넣고0.5 N NaOH- methanol
용액을가하여질소충전한후100°C
에서8
분간검 화하였다.
방냉후BF3-methanol
를이용하여fatty acid methyl ester (FAME)
로methylation
한후isooctane 2 mL
에녹여이를capillary column (Omegawax-320, 30 m×0.25 mm i.d., Su- pelco Co., Bellefonte, PA, USA)
이장착된gas chromatogra- phy (Glarus 600, PerkinElmer, USA)
로지방산을분석하였다. Carrier gas
는헬륨을사용하였고, injector
온도는250°C, de- tector (FID)
온도는270°C
로각각설정하였다.
이때column
온 도는180°C
에서8
분간유지하고, 3°C/min
속도로230°C/min
까 지증가시킨후,
항온상태에서10
분간유지하였으며split rate
는1:50
으로설정하였다.
지방산의동정은표준품menhaden oil
을동일조건으로분석한후, ECL (equivalent chain length)
법으로동정하였다(Ackman, 1969).
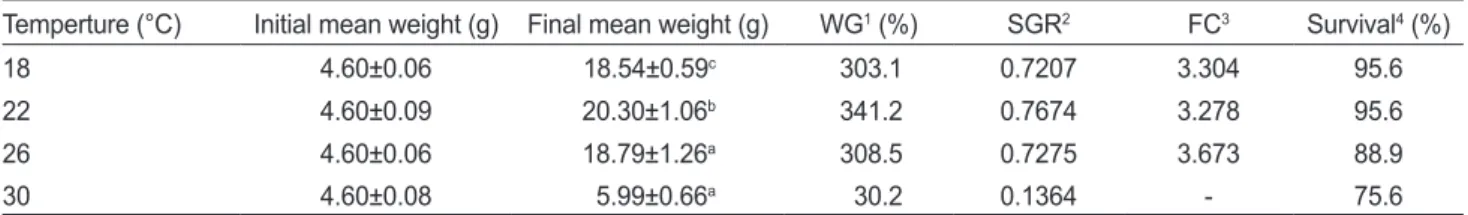
Fig. 1. Mean weight of sea cucumber Apostichopus japonicus grown at different water temperature for 12 weeks. Different letters indicate significant differences (P<0.05).
Mean body weight (g)
Water temperature (°C)
2520
15
10
5
0
18 22 26 30
Survival (%)
Water temperature (°C)
120100 80 60 40 20 0
18 22 26 30
고수온에서의 해삼 성장과 체지방 조성
403
통계처리
통계처리는
SPSS version 18.0 (SPSS Inc., Chicago, IL, USA) program
을 사용하여one-way ANOVA-test
를 실시한 후, Duncans multiple range test (Duncan, 1955)
로평균간의 유의성을검정하였다.
결 과
성장도
수온에따른해삼성장결과는
Table 1
과Fig. 1
에나타낸바 와같다.
실험초기평균습중량4.60 g
의해삼을수조당18°C, 22°C, 26°C
그리고30°C
로사육하였다.
사육수온이18°C
구 에서는4, 8
그리고12
주경과후각각6.96 g, 10.72 g
그리 고18.54 g
으로성장하였으며(P<0.05),
증중량은각각51.4%, 133.0%
그리고303.1%
로나타났다(P<0.05).
이때일간성장 률은4, 8
그리고12
주경과 후각각0.6431. 0.6561
그리고0.7207%/day
이었다.
사육 수온이22°C
구에서는4, 8
그리 고12
주경과후각각7.20 g, 12.10 g
그리고20.30 g
으로성 장하였으며(P<0.05),
증중량은 각각56.6%, 163.1%
그리고341.2%
로증가하였다(P<0.05).
이때일간성장률은4, 8
그리 고12
주경과후각각0.6957, 0.7502
그리고0.7674
이었다.
사 육수온이26°C
구에서는4, 8
그리고12
주경과후각각6.60 g, 10.81 g
그리고18.79 g
으로성장하였으며(P<0.05),
증중량은 각각43.5%, 135.0%
그리고308.5%
로나타났다(P<0.05).
이 때일간성장률은4, 8
그리고12
주경과후각각0.5601, 0.6625
그리고0.7275%/day
이었다.
사육수온이30°C
구에서는4, 8
그리고12
주경과후각각5.40 g, 5.80 g
그리고5.99 g
이었 으며,
증중량은각각17.4%, 26.2%
그리고30.2%
로나타났다(P<0.05).
이때일간성장률은4, 8
그리고12
주경과후각각0.2489, 0.1802
그리고0.1364%/day
이었다.
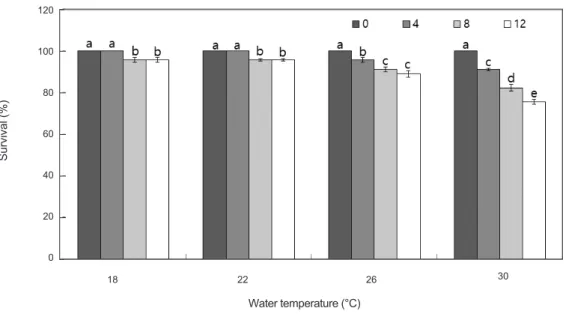
수온에따른해삼생존율은
Fig. 2
에나타낸바와같다.
사육 수온이18°C
구에서는4, 8
그리고12
주경과후각각100%, 95.6%
그리고95.6%
였으며,
사육 수온이22°C
구에서는각 각100%, 95.6%
그리고95.6%
로나타나18°C
에서의해삼과22°C
구에서의해삼의생존율은차이가없는것으로나타났다(P<0.05).
사육수온이26°C
구에서는4, 8
그리고12
주경과후각각
95.6%, 91.1%
그리고88.9%
이었으며(P<0.05),
사육수 온이30°C
구에서는각각91.1%, 82.2%
그리고75.6%
로나타 났다(P<0.05).
수온에따른생존율은사육수온이높아짐에따 라생존율은낮아지는것으로나타났다.
사육수질
사육기간 동안순환여과시스템 사육수질은염분농도
33.5- 33.8 psu,
용존산소6.38-6.62 mg/L, pH
는pH 8.01-8.13,
총 암모니아는0.120-0.208 mg/L
로 나타났으며,
총 부유물질은7.02-7.56 mg/L
이었다.
사육기간동안순환여과시스템내사 육수조의 수온은18±0.5°C, 22±0.5°C, 26±0.5°C
그리고30±0.5°C
로 유지되었으며,
염분농도33.5-33.8 psu,
용존산 소6.12-6.58 mg/L, pH
는pH 8.00-8.12,
총암모니아는0.132- 0.268 mg/L
로나타났으며,
총부유물질은7.38-8.06 mg/L
이 었다.
일반성분
수온에따른해삼의일반성분 분석 결과는
Table 2
에나타낸바와같다
.
사육수온이18°C
실험구해삼의경우수분함량91.8%,
조단백질3.73%,
지질0.58%
그리고조회분2.72%
이 었으며, 22°C
실험구경우수분함량은91.2%,
조단백질3.78%,
지질0.62%
그리고조회분2.98%
이었다.
사육수온이26°C
실험구해삼의경우수분함량91.7%,
조단백질3.92%,
지질0.51%
그리고조회분2.76%
였으며, 30°C
실험구경우수분 함량은91.3%,
조단백질3.88%,
지질0.53%
그리고조회분2.82%
였다.
사육수온에따른수분함량91.2-91.8%,
조단백질3.7-3.9%,
지질0.5-0.6%
그리고조회분2.7-3.0%
로나타났다. 지방산 조성
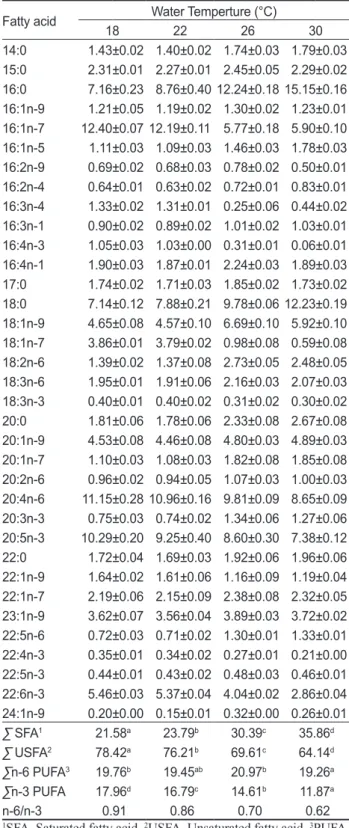
수온에따른해삼의총지질지방산조성은
Table 3
에나타낸바와같다
.
사육수온이18°C
실험구해삼의주요지방산조 성은16:1n-7, 20:4n-6, 20:5n-3, 16:0
그리고18:0
순으로각 각12.40%, 11.15%, 10.29%, 7.16%
그리고7.14%
로나타났 으며,
사육수온이22°C
구해삼주요지방산은16:1n-7, 20:4n- 6, 20:5n-3, 16:0
그리고18:0
순으로각각12.19%, 10.96%, 9.25%, 8.76%
그리고7.88%
로나타났다.
그리고사육수온 이26°C
실험구해삼주요지방산조성은16:0, 20:4n-6, 18:0,
Table 1. Growth performance of Apostichopus japonicus cultured at different water temperture for 12 weeks
Temperture (°C) Initial mean weight (g) Final mean weight (g) WG1 (%) SGR2 FC3 Survival4 (%)
18 4.60±0.06 18.54±0.59c 303.1 0.7207 3.304 95.6
22 4.60±0.09 20.30±1.06b 341.2 0.7674 3.278 95.6
26 4.60±0.06 18.79±1.26a 308.5 0.7275 3.673 88.9
30 4.60±0.08 5.99±0.66a 30.2 0.1364 - 75.6
1WG (Weight gain, %)=[(final sea cucumber weight-initial sea cucumber weight)/initial sea cucumber weight]×100. 2SGR (Specific growth rate, %/day)=[(loge final sea cucumber weight-loge initial sea cucumber weight)/days]×100. 3FC (Feed coefficient)=dry feed intake/wet weight gain. 4Survial rate (%)=(final sea cucumber /initial sea cucumber)×100.
정우철
ㆍ
Anisuzzaman Mdㆍ
Feng Jinㆍ
최종국ㆍ
한종철ㆍ
최병대ㆍ
강석중404
20:5n-3,
그리고18:1n-9
순으로각각12.24%, 9.81%, 9.78%, 8.60%
그리고6.69%
로나타났으며,
사육수온이30°C
실험구 해삼주요지방산은16:0, 18:0, 20:4n-6, 20:5n-3,
그리고18:1n- 9
순으로각각15.15%, 12.231%, 8.65%, 7.38%
그리고5.92%
로나타났다
.
특히,
포화지방산함량의차이가보였는데 사육 수온이18°C, 22°C, 26°C
그리고30°C
구에서각각21.58%, 23.79%, 30.39%
그리고35.86%
로사육수온이높아짐에따 라포화지방산의 함량도증가되는 것으로나타났으며, ∑n-3 PUFA
의경우도각각17.96, 16.79, 14.61%
그리고11.87%
로사육수온이증가함에따라
∑n-3 PUFA
함량은낮아지는것으로나타났다
(P<0.05).
고 찰
이번 실험을통하여해삼성장은
18°C, 22°C, 26°C
그리고30°C
로실험한결과대조구인18°C
구에비하여성장도는22°C
구에서높게나타났으며, 26°C
구에서는 유의적인차이가없는것으로났지만
,
극도의고수온구인30°C
구에서는낮게나 타났다.
이러한결과는지금까지수온과관련되어수행된해삼 의연구들이최적성장수온범위로알려진10-18°C (Li et al., 2002; Dong et al., 2006)
라는설정하에이루어졌기때문에비 교할수있는문헌을찾기가매우어렵다.
수온25°C
이상되 는시기에는대사활동을최소화시켜성장을멈추거나,
소화관 이퇴화되어하면(夏眠)
하는것으로알려져있으며(Liu et al., 1996; Yang et al., 2006; Ji et al., 2008),
해삼의이런생태적 인특성때문에사계절이뚜렷한우리나라의자연환경에서는 해삼의성장기간이짧아해삼양성이불리한것으로인식되고 있다(Jeong et al., 2016a).
수온변화에따른해삼의행동특성 에서수온28-30°C
에서의해삼행동특성은실험시작6
시간이 경과한이후에는 다른수온
(10-25°C)
보다더욱많은개체들이활발하게활동을하는것이관찰되었고
,
실험수조의밑바 닥과구석진곳에대거몰려있는모습이관찰되었다(Jeong et al., 2016a).
실험종료직후에는폐사체가나타났으며이들폐 사체의공통점은껍질이벗겨지고혹은녹아서표면이하얗게 Fig. 2. Survival of sea cucumber Apostichopus japonicus grown by different water temperature for 12 weeks. Different letters indicate significant differences (P<0.05).Mean body weight (g)
Water temperature (°C)
2015
10
5
0
18 22 26 30
Survival (%)
Water temperature (°C)
120100 80 60 40 20
0
18 22 26 30
Table 2. Proximate compositions of sea cucumber Apostichopus japonicus cultured at different water temperture (%)
Temperture (°C) Nutrient content
Moisture Protein Lipid Ash Carbohydrate
18 91.83±0.28c 3.73±0.03a 0.58±0.02a 2.72±0.01a 1.14±0.02a
22 91.23±0.40c 3.78±0.09a 0.62±0.06a 2.98±0.02a 1.39±0.02a
26 91.68±0.22c 3.92±0.07a 0.51±0.03a 2.76±0.01a 1.13±0.02a
30 91.29±0.20c 3.88±0.12a 0.53±0.01a 2.82±0.01a 1.48±0.02a
Nutrient content date were calculated on a dry matter basis. Data with different letters in the same column means
significant difference among different diet treatments (P<0.05).
고수온에서의 해삼 성장과 체지방 조성
405
변하였다는것이다
.
본실험결과와기존에알려진수온생태와의가장큰차이점은수온이
25°C
이상되는시기에는대사활동을최소화시켜성장을멈추거나
,
소화관이퇴화되어하면(夏 眠)
한다고알려져있지만본실험에서는하면없이성장하였으 며, Jeong et al. (2016b)
의연구에서도25°C
이상되는시기에서 도성장이보고되고있다.
이러한차이는해삼개체크기와자연 산과양식산의차이에기인된것으로생각할수있다.
본실험에 사용한해삼은당년생의어린해삼이면서고수온에천천히적 응시킨점이다.
전생활사를양식장환경에적응된어미해삼을 사용하면그종자는이미가축화되어서고수온에서도문제없이 성장할수있다는설이다.
그러나현장의암묵적지식과경험으로는같은방법으로얻은종자를사용했을때
26-30°C
의높은수온으로상승시키면모두폐사하므로가설설정이잘못되었다 고주장한다
.
그러나여기에대한연구자의의견은해삼의생리 적특성보다는수온상승에따른양식환경의철저한관리부재 에서기인된결과라고사료된다.
해삼사료의특성상공급한사 료는사육조바닥에침체하면서부패하기쉽다.
수온이상승함 으로써부패로인한수질오염은가속화될수밖에없기때문에 수온에따른사료관리가아주중요하다.
해삼사료의특성상액 상반죽사료를공급할때사료가수중에퍼져서수질혼탁을초 래하기때문에수질오염을최소화할수있는사료공급방법도 중요하다.
이러한문제점을해결하기위하여피스톤식유리관피펫을사용하여수조바닥층에깔아주는방법
(Anisuzzaman
et al., 2018)
을사용하였다.
본연구실에서개발한사료공급방 법을통하여전사육기간pH
는pH 8.00-8.12,
총암모니아는0.132-0.268 mg/L
그리고총부유물질은7.38-8.06 mg/L
으로 서해삼사육을위한적합한범위내에서유지가되어수질이해 삼성장에영향을준것은없는것으로사료된다.
Table 2
에나타낸총지질은051-0.62%
범위로서수온의변화에차이는없었으며
, Table 3
의해삼체지방의지방산조성에서는고수온으로갈수록포화지방산은증가되고있는반면 에불포화지방산은감소되는것으로나타났다
.
이러한수온에 따른결과는다른양식생물인연어(Kepshier et al., 1983)
와틸 라피아(Satoh et al., 1984)
연구에서도같은결과가보고되고 있다.
사육수온증가와관련하여 지방산계열별로 비교하면 오메가-9
지방산은변화가없었으며,
오메가-6
계열의20:4n-6
지방산은11.15%
에서8.65%
로 감소하였으며,
오메가-3
계 열의20:5n-3 (EPA)
도10.29%
에서7.38%
로 감소하였으며, 22:6n-3 (DHA)
는5.46%
에서2.86%
로감소하는것으로나타 났다. Table 3
의23:1n-9 (cis-14-tricosenoic acid)
의홀수지방 산은다른양식생물에서는볼수없는특이한지방산으로실험구에따라
3.5-3.9%
로서실험구간에유의적차이는나타나지않았다
.
이지방산은먹이로부터기인된것이아니라24:1n-9
가출현하는점으로보아alpha-oxidation
의결과물로사료된 다(Kaneniwa et al., 1986; Feng et al., 2016).
이상의결과를종합해보면이번실험을통하여수온
25°C
이상에서도성장이되Table 3. Total lipids fatty acid profiles of sea cucumber Aposticho- pus japonicus cultured at different water temperature (%) Fatty acid Water Temperture (°C)
18 22 26 30
14:0 1.43 ±0.02 1.40 ±0.02 1.74 ±0.03 1.79 ±0.03 15:0 2.31 ±0.01 2.27 ±0.01 2.45 ±0.05 2.29 ±0.02 16:0 7.16 ±0.23 8.76 ±0.40 12.24 ±0.18 15.15 ±0.16 16:1n-9 1.21 ±0.05 1.19 ±0.02 1.30 ±0.02 1.23 ±0.01 16:1n-7 12.40 ±0.07 12.19 ±0.11 5.77 ±0.18 5.90 ±0.10 16:1n-5 1.11 ±0.03 1.09 ±0.03 1.46 ±0.03 1.78 ±0.03 16:2n-9 0.69 ±0.02 0.68 ±0.03 0.78 ±0.02 0.50 ±0.01 16:2n-4 0.64 ±0.01 0.63 ±0.02 0.72 ±0.01 0.83 ±0.01 16:3n-4 1.33 ±0.02 1.31 ±0.01 0.25 ±0.06 0.44 ±0.02 16:3n-1 0.90 ±0.02 0.89 ±0.02 1.01 ±0.02 1.03 ±0.01 16:4n-3 1.05 ±0.03 1.03 ±0.00 0.31 ±0.01 0.06 ±0.01 16:4n-1 1.90 ±0.03 1.87 ±0.01 2.24 ±0.03 1.89 ±0.03 17:0 1.74 ±0.02 1.71 ±0.03 1.85 ±0.02 1.73 ±0.02 18:0 7.14 ±0.12 7.88 ±0.21 9.78 ±0.06 12.23 ±0.19 18:1n-9 4.65 ±0.08 4.57 ±0.10 6.69 ±0.10 5.92 ±0.10 18:1n-7 3.86 ±0.01 3.79 ±0.02 0.98 ±0.08 0.59 ±0.08 18:2n-6 1.39 ±0.02 1.37 ±0.08 2.73 ±0.05 2.48 ±0.05 18:3n-6 1.95 ±0.01 1.91 ±0.06 2.16 ±0.03 2.07 ±0.03 18:3n-3 0.40 ±0.01 0.40 ±0.02 0.31 ±0.02 0.30 ±0.02 20:0 1.81 ±0.06 1.78 ±0.06 2.33 ±0.08 2.67 ±0.08 20:1n-9 4.53 ±0.08 4.46 ±0.08 4.80 ±0.03 4.89 ±0.03 20:1n-7 1.10 ±0.03 1.08 ±0.03 1.82 ±0.08 1.85 ±0.08 20:2n-6 0.96 ±0.02 0.94 ±0.05 1.07 ±0.03 1.00 ±0.03 20:4n-6 11.15 ±0.28 10.96 ±0.16 9.81 ±0.09 8.65 ±0.09 20:3n-3 0.75 ±0.03 0.74 ±0.02 1.34 ±0.06 1.27 ±0.06 20:5n-3 10.29 ±0.20 9.25 ±0.40 8.60 ±0.30 7.38 ±0.12 22:0 1.72 ±0.04 1.69 ±0.03 1.92 ±0.06 1.96 ±0.06 22:1n-9 1.64 ±0.02 1.61 ±0.06 1.16 ±0.09 1.19 ±0.04 22:1n-7 2.19 ±0.06 2.15 ±0.09 2.38 ±0.08 2.32 ±0.05 23:1n-9 3.62 ±0.07 3.56 ±0.04 3.89 ±0.03 3.72 ±0.02 22:5n-6 0.72 ±0.03 0.71 ±0.02 1.30 ±0.01 1.33 ±0.01 22:4n-3 0.35 ±0.01 0.34 ±0.02 0.27 ±0.01 0.21 ±0.00 22:5n-3 0.44 ±0.01 0.43 ±0.02 0.48 ±0.03 0.46 ±0.01 22:6n-3 5.46 ±0.03 5.37 ±0.04 4.04 ±0.02 2.86 ±0.04 24:1n-9 0.20 ±0.00 0.15 ±0.01 0.32 ±0.00 0.26 ±0.01
∑ SFA1 21.58a 23.79b 30.39c 35.86d
∑ USFA2 78.42a 76.21b 69.61c 64.14d
∑n-6 PUFA3 19.76b 19.45ab 20.97b 19.26a
∑n-3 PUFA 17.96d 16.79c 14.61b 11.87a
n-6/n-3 0.91 0.86 0.70 0.62
1SFA, Saturated fatty acid. 2USFA, Unsaturated fatty acid. 3PUFA, Poly unsaturated fatty acid. The values are mean±S.D. (n=3). a-
dDifferent superscript letters within rows represent significant dif- ferences between treatments (P<0.05).
는것으로확인하였으며
,
수온이높아질수록공급되는수질변 화가빠르게나타나기때문에사육환경유지가무엇보다중요 하다고사료된다.
사 사
이연구는
2016
년도경상대학교연구년제연구교수연구지원비에의하여수행되었음
.
References
Anisuzzaman MD, Jeong UC, Jin F, Kabery K, Choi JK, Lee DI, Yu HS and Kang SJ. 2018. Effects of different algae in diet on growth and interleukin (IL)-10 production of juve- nile sea cucumber Apostichopus japonicus. Fish Aquat Sci 20: 1. 20-14. https://doi.org/10.1186/s41240-017-0069-5.
AOAC (Official method of Analysis). 2000. Association of of- ficial analytical chemists. 17th ed. Maryland, U.S.A., 59-60.
Bligh EG and Dyer WJ. 1959. A rapid method of total lipid ex- traction and purification. Can J Biochem Physiol 37, 911- 917. https://doi.org/10.1139/o59-099.
Dong YW, Dong SL, Tian XL, Wang F and Zhang MZ. 2006.
Effects of diel temperature fluctuations on growth, oxygen consumption and proximate body composition in the sea cucumber Apostichopus japonicus Selenka. Aquaculture 255, 514-521. http://dx.doi.org/10.1016/j.aquaculture.2005.
12.013.
Duncan DB. 1955. Multiple range and multiple F tests. Biomet- rics 11, 1-42. https://doi.org/10.2307/3001478.
Feng J, Anisuzzaman MD, Jeong UC, Choi JK, Yu HS, Kang SW and Kang SJ. 2016. Comparison of fatty acid composi- tion of wild and cultured Sea cucumber Apostichopus ja-
ponicus. Korean J Fish Aquat Sci, 49, 474-485. http://dx.doi.
org/10.5657/KFAS.2016.0474.
Fredalina BD, Ridzwan BH, Zainal Abidin AA, Kaswandi MA, Zaiton H, Zali I and Kittakoop P. 1999. Fatty acid com- position in local sea cucumber, Stichopus chloronotus, for wound healing. Gen Pharmacol 33, 337340. http://dx.doi.
org/10.1016/S0306-3623(98)00253-5.
Gao F, Yang HS and Xu Q. 2009. Seasonal variations of fatty acid composition in Apostichopus japonicus body wall. Mar Sci 33, 15-19. http://dx.doi.org/10.1007/s00343-011-0041- Jeong UC, Jin F, Choi JK, Han JC, Choi BD and Kang SJ. 2016a. 7.
A Laboratory-scale recirculating aquaculture system for Sea cucumber Apostichopus japonicas. Korean J Fish Aquat Sci 49, 343-350. .https://doi.org/10.5657/kfas.2016.0343.
Jeong UC, Han JC and Kang SJ. 2016b. Sea Cucumber Apos-
tichopus japonicas growth in Pond-bottom and Lantern-net
Cultures. Korean J Fish Aquat Sci 49, 337-342. https://doi.org/10.5657/kfas.2016.0337.
Ji TT, Dong YW and Dong SL. 2008. Growth and physiologi-
cal responses in the sea cucumber, Apostichopus japonicus Selenka: Aestivation and temperature. Aquaculture 283, 180- 187. http://dx.doi.org/10.1016/j.aquaculture.2008.07.006.
Jin F, Choi JK, Jeong UC, Anisuzzaman M, Rye CH, Choi BD and Kang SJ. 2016a. Effects of fermented fecal solid diets on growth of the sea cucumber Apostichopus japonicus. Ko- rean J Fish Aquat Sci 49, 161-167. https://doi.org/10.5657/
kfas.2016.0161.
Jin F, Anisuzzaman M, Jeong UC, Choi JK, Yu HS, Kang SW, Kang SJ. 2016b. Comparison of fatty acid composi- tion of wild and cultured sea cucumber Apostichopus ja-
ponicus. Korean J Fish Aquat Sci 49, 474-485. https://doi.
org/10.5657/kfas.2016.0474.
Kaneniwa M, Itabashi Y, Endo S and Takagi T. 1986. Fatty ac- ids in holothuroidea: occurrence of cis-14-tricosenoic acid.
Comparative Biochem Physiolo B 84, 451-455. http://
dx.doi.org/10.1016/0305-0491(86)90105-7.
Kang SJ. 2012. Sea cucumber aquaculture technology. In:
Stocking and transportation of sea cucumber. Aqua Info Press, Seoul, Korea, 310-318.
Kepshier BM, Tinsley IJ and Lowry RR. 1983. Effect of tem- perature on the chemical composition of pink salmon (On-
corhynchus gorbuscha) muscle. Aquaculture 32, 295-301.
https://doi.org/10.1016/0044-8486(83)90226-0.
Li B, Yang H, Zhang T, Zhou Y and Zhang C. 2002. Effect of temperature on respiration and excretion of sea cucumber
Apostichopus japonicus. Oceanol Limnol Sin 33, 182-187
Liu Y, Li F, Song B, Sun H, Zhang X and Gu B. 1996. Study onaestivating habit of sea cucumber Apostichopus Japonicus selenka I. Ecological characteristic of aestivation. J Fish Sci China 3, 41-49.
Satoh S, Takeuchi T and Watanabe T. 1984. Effects of starvation and environmental temperature on proximate and fatty acid compositions of Tilapia Nilotica. Nippon suisan gakkaishi 50, 79-84. https://doi.org/10.2331/suisan.50.79.
Slater MJ and Carton AG. 2007. Survivorship and growth of the sea cucumber Australostichopus (Stichopus) mollis (Hutton 1872) in polyculture trials with green-lipped mussel farms.
Aquaculture 272, 389-398. https://doi.org/10.1016/j.aqua- culture.2007.07.230.
Shahabuddin AM, Khan MND, Mikami K, Araki T and Yoshi- matsu T. 2017. Dietary supplementation of red alga Pyropia
spheroplasts on growth, feed utilization and body compo-
sition of sea cucumber, Apostichopus japonicus (Selenka).Aquacult Res 48, 5363-5372. https://doi.org/10.1111/
are.13350.
Wen B, Gao Q, Dong S, Hou Y, Yu H and Li W. 2016. Effects of different feed ingredients on growth, fatty acid profiles, lipid peroxidation and aminotransferases activities of sea cucumber Apostichopus japonicus (Selenka). Aquacul- ture 454, 176-183. http://dx.doi.org/10.1016/j.aquacul- ture.2015.12.027.
고수온에서의 해삼 성장과 체지방 조성
407
Yang HS, Zhou Y, Zhang T, Yuan XT, Li XX Liu Y and Zhang F. 2006. Metabolic characteristics of sea cucumber, Apos-
tichopus japonicus (Selenka) during aestivation. Experim
Mar Biol Ecolo 330, 505-510. http://dx.doi.org/10.1016/j.jembe.2005.09.010.
Yu Z, Zhou Y, Yang H, Ma Y and Hu C. 2014. Survival, growth, food availability and assimilation efficiency of the sea cu- cumber Apostichopus japonicus bottom-cultured under a fish farm in southern China. Aquaculture 426, 426-427.
https://doi.org/10.1016/j.aquaculture.2014.02.013.