215
대장균에서 비천연 아미노산의 위치특이적 삽입을 위한
Amber Suppressor tRNA 와 Aminoacyl-tRNA Synthetase 의 Amber Suppression 활성측정시스템 개발
김경태·박미영·박중찬*
한국외국대학교 자연과학대학 생명공학과
대장균에서비천연아미노산을단백질생합성시특정위치에삽입하는방법의하나로 amber suppressor tRNA와 여기에비천연아미노산을특이적으로결합할수있는변형된 aminoacyl-tRNA synthetase 쌍을이용한다. 이러한 기술의개발을위해필요한여러요소중하나는이러한시스템이대장균에서얼마나잘작동하는지를확인할수 있는in vivo보고시스템을설정하는것이다. 본논문에서는β-galactosidase 유전자의 N-말단에 amber 코돈을삽입 한보고유전자를제작하였으며, 이를대장균(DH10B)의 chromosomal DNA에삽입하여 DH10B(Tn:lacZam) 균주 를개발하였다. Genomic PCR과 Southern blot 분석을통하여lacZ amber 유전자가대장균의염색체에삽입된것 을 확인하였으며, DH10B(Tn:lacZam)은 amber suppression을 유도할 수 있는 벡터가형질 전환될경우만 β- galactosidase 활성을 나타냈다. DH10B(Tn:lacZam)에 효모균의 amber suppressor tRNATyr와 Tyrosyl-tRNA synthetase 쌍을동시에발현하는벡터를형질전환하였을때, amber suppression에의해서β-galactosidase 활성이 나타났다. 하지만이활성은대장균의 amber suppressor tRNAGln를발현하는 pSupE2를형질전환하였을때와비교 하여매우낮은β-galactosidase 활성을나타냈다. 이러한결과는 DH10B(Tn:lacZam) 균주가β-galactosidase 활성을 통하여정성및정량적으로in vivo amber suppression 활성을비교분석할수있는특성을가졌음을나타낸다. Key words □ amber suppression, aminoacyl-tRNA synthetase, suppressor tRNA, unnatural amino acid
자연계에일반적으로존재하는 20종의아미노산이 아닌새로 운생화학적특성을나타내는비천연아미노산을단백질의특정 위치에삽입하는기술은물리적, 화학적및생물학적특성이강
화된새로운 단백질을만드는좋은방법이 될수있을뿐만아 니라단백질의 기능연구에도활용될 수있다(3, 11, 18, 19). 앞
서 이러한 시도가 다양한 방법으로 in vitro에서 시도되었다.
Nonsense 코돈을 인식하는 suppressor tRNA에비천연 아미노산
을화학적으로결합한후, 이를in vitro번역시스템에넣어특정 위치에아미노산이 삽입되도록한다(2, 7, 12). 이방법은 매우 다양한비천연아미노산을여러위치에비교적용이하게삽입할 수있는장점이있으나, 세포밖에서단백질을합성한다는점에 서생산성이매우낮고, aminoacyl-tRNA의화학적 합성이 쉽지 않다는점에서한계를가지고있다.
반면, 비천연아미노산을 in vivo에서단백질의특정위치에삽 입하는방법은높은생산성과정확성을얻을수있을뿐만아니 라, 단백질의구조와기능연구를 in vivo와 in vitro에서모두할 수있다는 점에서 장점을 가지고있다. 이방법의 기본원리는
suppressor tRNA와여기에 비천연아미노산만을특이적으로 결
합하는 aminoacyl-tRNA synthetase (ARS)를유전자 변형의방법
을통하여얻어서 이를세포안에안정적으로 발현시켜줌으로 써단백질생합성시특정위치에 비천연아미노산이 삽입되도록 하는것이다(16, 17). 이를위해사용되는 suppressor tRNA는숙 주가가지고 있는내재적인 ARS에의해 aminoacylation이일어 나지않아야하며, 오직외부에서추가해주는 ARS에의해서만 aminoacylation이일어나야한다. 이를위하여 숙주가가지고있 는 tRNA를돌연변이시켜내재 ARS에의해서 aminoacylation이
일어나지 않도록 하거나(8), 숙주와는 다른 생물계의 tRNA와
ARS 쌍을 활용하는 것이다. 이러한예로써 효모균의 tRNATyr, tRNAGln, tRNAAsp, 그리고 archaebacteria인 Methanococcus jannaschii의 tRNATyr, 등이 대장균의 내재 ARS에 의해서
aminoacylation이되지 않는 이종 tRNA로알려져있으며, 이들
각각의 tRNA에 특정천연 아미노산을 결합하는효모 또는 M.
jannaschii의 ARS 또한 대장균의 내재 tRNA를 aminoacylation
하지않는다는점에서 suppressor tRNA와함께비천연아미노산 을삽입하기위한 ARS로활용되고있다(9, 12, 13, 14). 일반적
으로이종 suppressor tRNA와 ARS 쌍을선정하고, ARS에돌연 변이를유발하여라이브러리를만들고, 이로부터비천연아미노
산 만을 suppressor tRNA에 결합할수있는 변형된 ARS를선 별함으로써 in vivo비천연 아미노산을삽입하는시스템이개발 될 수있다(17). 따라서 이종 suppressor tRNA와 ARS 쌍의 선
별이 in vivo 시스템의개발을 위하여 필수적이고, 이들이 숙주
*To whom correspondence should be addressed.
Tel: 82-31-330-4355, Fax: 82-31-330-4566 E-mail: [email protected]
세포내에서숙주의 내재 tRNA와 ARS의영향을받지않고독 립적으로 amber suppression 활성을 나타낼 수있는가를확인하 는것이필요하다.
본논문에서는 대장균에서 amber suppression 활성을 빠르고
쉽게확인할수있는보고자 시스템을만들었다. Amber 코돈이
삽입된 β-galactosidase 유전자를대장균의 chromosome에안정적 으로삽입하였으며, 다양한조합의 suppressor tRNA/ARS 유전자 를형질전환하여균주가 amber suppression이일어날수있는조 건에서만β-galactosidase 활성이나타남을확인하였다
재료 및 방법 배지 및 균주 배양
본연구에 사용된 대장균은 50µg/ml ampicillin을첨가한 LB
배지 또는 변형된 M9 최소배지(1 L 배지 당 6 g Na2HPO4, 3 g KH2PO4, 1 g NH4Cl, 0.5 g NaCl, 1 g Yeast extract, 10 ml 20% 포도당용액, 1 M MgSO4 2 ml, 1 M CaCl2 0.1 ml)에서 37oC
또는 30oC에서배양하였으며, 각균주의특성은 Table 1에나타 나있다. β-Galactosidase의 정량 분석을위한 대장균의 배양은
분석하루 전에단일 colony를배지에 접종하여 18시간초기배
양을 한 후, 새로운 배지에 초기배양을 접종하였다. 30oC에서
24~72 시간진탕배양후, 세균을수확하여분석하였다.
플라스미드 제작
Amber 코돈을가지고있는 lacZ유전자를증폭하기위하여앞
서제작된 pGamb-1 (1)을주형으로사용하였다. pGamb-1은 lacZ
유전자의 N-말단 8개의 아미노산 잔기가 제거되고 대신 앞에
ATG 시작코돈과 1개의 amber 코돈이포함된 cassette 염기서열 을가지고있다. 이벡터의 lac promoter에서lacZ유전자의 1955
번 뉴클레오타이드에 위치한 제한요소 SacI자리를 포함하는
2.1 kb DNA 절편을 PCR로 증폭하였다. 사용된 primer 서열은
LacF (5’-CAATACGCAAACCGCCTCTCC-3’)와 LacR (5’- CAGAGGAGCTCGTTATCG-3’)이다. 생성된 DNA 절편을 SmaI
과 SacI 효소를 이용하여 pUT/mini-Tn5 Km 벡터(5)의 lac
promoter와 lacZ 유전자의 N-말단부위와 치환함으로써
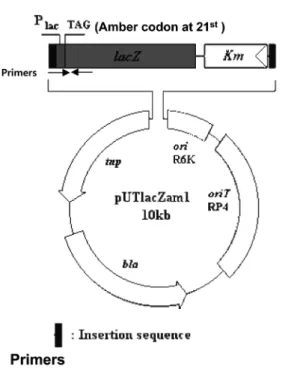
pUTlacZam1을제작하였다.
플라스미드 pSupE2의제작을 위하여 E. coli tRNAGln (glnV)의 염기서열(EcoGeneID: EG30029)을바탕으로 amber 코돈을 인식 하기 위한 anticodon 위치에 A35 돌연변이와 amber suppression
효율을 높이기 위해 A37 돌연변이가 추가 도입된 amber
suppressor tRNA를암호화하는유전자를 2개의 oligonucleotides로 제작하여만들었다(4). 제한효소 EcoRI 자리와 suppressor tRNA
의 5’-말단을 포함하는 59mer (5’-CCGGAATTCTGGGGTAT CGCCAAGCGGTAAGGCACCGGATTCTAAATCCGGCATTCCG A-3’)와 PstI 자리와 suppressor tRNA의 3’-말단을 포함하는
55mer (5’-AAAACTGCAGTGGCTGGGGTACGAGGATTCGAA CCTCGAACCTCGGAATGCCGGATTTAG-3’)의 oligonucleotide
를합성하였다. 이 2개의 oligonucleotides는 3’-말단에서로상보 적인뉴클레오타이드가 18개있다. 각각의 oligonucleotide를 500 pmole/ml의 농도로 TE buffer에 녹인 후, 두 oligonucleotides의 상보적인염기서열이혼성화되도록 65oC에서 15분간반응시키고 Klenow 효소(Promega, USA)를첨가하여 3’-말단을 채움으로써
suppressor tRNA를암호화한 2중가닥의 DNA를만들었다. 이를 EcoRI/PstI 효소를 이용하여 pBluescript SK에 클로닝하여
pSupE2를 제작하였다. 그리고 음성 대조군으로 사용하기 위해
supE2를역방향으로클로닝하여 pRSupE2라하였다.
효모균의 amber suppressor tRNATyr를발현하는벡터의제작도
pSupE2 제작과 유사하게 하였다. S. cerevisiae (Sc) cytoplasmic tRNATyr (SUP3, GeneID: 854140)의 anticodon을 amber 코돈을 인식하도록 치환한 tRNA 염기서열을 5’-말단부터 68mer (5’- ATAGCTCCGCGGCTCGAGCTCTCGGTAGCCAAGTTGGTTTA AGGCGCAAGACTCTAAATCTTGAGATC-3’)와 3’-말단부터 Table 1. Genotypes of bacterial strains and characteristics of plasmids used in this study
Strains / Plasmids Genotype / Characteristics Sources
Escherichia coli strains
DH10B F-, mcrA, ∆(mrr-hsdRMS-mcrBC), Φ80d lacZ∆M15, ∆lacX74, deoR,
recA1, endA1, araD139, ∆(ara, leu)7697, galU, galK, λ-rpsL, nupG Gibco BRL S17-1 λ pir Tpr, Smr, recA, thi, hsdRM+, RP4::2-Tc::Mu::Km Tn7, λ(pir) (5) DH10B(Tn:lacZam) DH10B containing chromosomal integration of lacZam-Kmr cassette
Plasmids
pGamb-1 Apr,one amber codon at the N-terminal region of lacZ (21st residue) (1) pUTlacZam1 Tnp, oriR6K, oriTRP4, Apr, Tn5, Kmr; delivery plasmid for chromosomal integration of
lacZam-Kmr cassette This study
pSupE2 supE(A37): E. coli amber suppressor tRNAGln(A37) in pBluescript-SK (Stratagene), Apr This study
pRSupE2 Reverse orientation of supE(A37) in pBluescript-SK, Apr This study
pSuptRNATyr (Sc) S. cerevisieae amber suppressor tRNATyr in pBluescript-SK, Apr This study pSuptRNATyr -TyrRS(Sc) S. cerevisieae amber suppressor tRNATyr and Tyrosyl-tRNA synthetase gene in
pBluescript-SK, Apr This study
55mer (5’-AATAATAAGCTTTGGTCTCCCGGGGGCGAGTCG AACGCCCGATCTCAAGATTTAG-3’)를 합성한 oligonucleotide
를제작하였다. 이들은각각 5’-말단에클로닝을위하여 SacII와
SpeI 자리를 가지고있으며, 앞서언급한같은방법으로이중가
닥의 DNA를만들어 SacII/SpeI 효소를이용하여 pBluescript SK
에클로닝하여 pSuptRNATyr(Sc)를제작하였다.
플라스미드 pSuptRNATyr-TyrRS(Sc)의제작을위하여효모균의
genomic DNA를 주형으로 하여 Sc tyrosyl-tRNA synthetase (TyrRS, GenBank ID: L12221.1)를암호화한 DNA 절편을 PCR
를수행하여 얻었다. 사용된 2개 primers의염기서열은 다음과 같다: YRSF (5’-ATAGTCTGCAGGAAGGAGCTACGTAGTCC TCTGCTGCC-3’), YRSR (5’-AACGTCCTCGAGTTACAATTT GGTTTC-3’). 사용된 primers는각각 PstI과 XhoI 자리를가지고 있으며, 약 1.3 kb PCR 산물을 pGEM-T easy vector (Promega, USA)에 클로닝한 후, 이를 다시 PstI/XhoI 효소로 잘라
pSuptRNATyr (Sc)에 클로닝 함으로써 pSuptRNATyr-TyrRS(Sc)를 만들었다. 본연구에사용된모든플라스미드의특성은 Table 1
에표시되어있다.
DH10B(Tn:
lacZam)
생성 및 확인돌연변이 유전자 lacZ amber를 E. coli DH10B 균주의
chromosome에 삽입하기 위하여 rifampicin (50µg/ml)이포함된
LB 배지에서 DH10B를 2~3일간배양하여자연돌연변이에의해
rifampicin에 저항성을 나타내는 클론을 얻었다. 이를 다시
rifampicin이 첨가된 LB 배지에서 하루 동안 배양하였다.
pUTlacZam1을형질전환한 E. coli S17-1λpir donor 균주를 역시
200µg/ml ampicillin이 포함된 LB에서하루 동안 배양하였다.
각각배양액 50µl를원심분리로수확하고, 둘을합친후, 50 µl
의새로운 LB에풀었다. 이를 LB 한천배지에 spotting하여 37oC
에서하루동안배양한후, 형성된 colony를긁어모아 LB 배지 에 희석하였다. 접합과정을 거친 희석된 대장균을 50µg/ml rifampicin, 10µg/ml kanamycin, 40µg/ml X-Gal을포함하는 LB
한천배지에서 37oC 하루 동안 배양하였다. 이과정에서 donor
균주는 rifampicin에민감하여생장을 못하며, rifampicin에저항 성을나타내고접합에의해서 pUTlacZam1를전달받은 DH10B
균주만 생장한다. 흰색 세균군락을 선별하여 10µg/ml kanamycin 또는 50µg/ml ampicillin이포함된 LB 한천배지에접 종하여 kanamycin 배지에서는 생장하나, ampicillin 배지에서는 생장하지못하는클론을선별하였다.
유전자 lacZ amber의 chromosome 삽입을 확인하기 위하여
DH10B와 선별된 양성 클론으로부터 TRIzol 용액(Invitrogen, USA)을 이용하여 genomic DNA를 분리하였으며, 분리된 DNA 1µg을 주형으로 lacZ amber와 glyceraldehydes-3-phosphate dehydrogenase (GAPDH, gapA, EcoGeneID:EG10367) 유전자에
대한 PCR을 수행하였다. 사용된 lacZ amber 유전자의 PCR primer는 5’-GAATACTCAAGCTTGGTCGAC-3
’
과 5’- TGCATCTGCCAGTTTGAGGG-3’
으로각각 lacZ 유전자 N-말단 에치환되어 삽입된 amber cassette 부위와 lacZ 유전자 N-말단부위와상보적인염기서열을가지고있어, PCR 반응시 278 bp의 절편을 생성한다. 내재대조군 유전자인 GAPDH PCR primer의 염기서열은 5’-GTAGGTATCAACGGTTTTGG-3
’
과 5’- CATCAGACCTTCGATGATG-3’
이며 대장균 GAPDH 유전자13~519 뉴클레오타이드를 증폭하여 507 bp 크기의절편을생성
한다.
Southern blot
분석DH10B(Tn:lacZam)로부터 TRIzol 용액(Invitrogen, USA)을이
용하여 genomic DNA를 분리하였다. 분리된 DNA 5µg을
EcoRV, BglII, SphI, PstI 효소로 37oC에서 20시간동안자른후,
1.0% agarose gel에서 전기영동하여 분리하였다. 젤을
denaturation 용액(1.5 M NaCl, 0.5 M NaOH)에서 40분간
denaturation 시킨후, nylon 막에전이를 하였다. 막에 DNA 절 편을 UV로 고정한 후, 혼성화 용액(5× SSC)에서 47oC, 4시간 동안 반응하였으며, PCR DIG-probe 합성키트(Roche Applied
Science, Germany)를이용하여 생산자 사용법에 따라 만들어진
DIG-표지된탐침(lacZ amber 유전자의 chromosome 삽입을확인 하기위하여앞서언급된 278 bp PCR 산물)을첨가한후, 같은 온도에서 14시간동안추가반응을시행하였다. 반응을마친막
을 65oC에서 15분간 2회 같은 용액으로 세척한 후, anti- Digoxigenin-alkaline phosphatase 항체(Roche Applied Science, Germany)와 NBT/BCIP 기질을 이용하여진한파란색의혼성화 된밴드를감지하였다.
플라스미드 형질전환 및 β-
galactosidase
활성분석Suppressor tRNA 및 TyrRS 발현벡터를 42oC heat shock 방법 에 의해서 DH10B(Tn:lacZam)에 형질전환한 후, 50µg/ml ampicillin이포함된 LB 한천배지에서 배양하였다.다음날 단일
colony를새로운 LB 배지에서배양한후 β-galactosidase 분석에 사용하였다. 1.0 ml의새균배양을 12,000 rpm에서 1분간원심분리
하여 수확하였다. 수확된 세균을 1.0 ml의 Z buffer (60 mM Na2HPO4, 40 mM NaH2PO4, 10 mM KCl, 1 mM MgSO4, 50 mM 2-mercaptoethanol)로세척하고, 200µl의 Z buffer에다시 현탁시 켰다. 여기에 4방울의 chloroform과 2방울의 0.1% SDS를첨가한
후 5초간 vortex 하여 β-galactosidase 효소를 추출하였다. 12,000×g에서 2분간 원심분리한후상층액을 새로운시험관에 옮겼다. 100µl의추출액과 ortho-nitrophenyl-β-galacoside (ONPG)
를기질로 사용하여 37oC에서 20분간 효소반응을 수행한후 1 M Na2CO3 0.5 ml를넣고 반응을정지시켰다. 시료를 A600, A550,
그리고 A420에서 각각의 흡광도를 측정하여 β-galactosidase의
Miller Unit (MU)을 결정하였다(6). DH10B가 LacI repressor를
과다 발현하지않는균주임으로 β-galactosidase 활성 분석을위 한 실험에서 Isopropyl-β-D-thio-galactoside (IPTG)와 같은 lac promoter의 inducer는사용하지않았으며, 예비실험에서배양배지 에 IPTG를 포함한 경우도 amber suppression에 의한 β- galactosidase 활성에큰영향을미치지않았다.
결과 및 고찰
In vivo
amber suppression
활성측정을 위한DH10B (Tn:
lacZam)
균주 생성대장균에서이종 amber suppressor tRNA와 ARS 쌍을이용한
비천연 아미노산의 삽입기술 개발을 위하여 in vivo amber suppression 활성을측정할 수있는균주개발을 시도하였다. 이 를위해 transposon에의해대장균 chromosome에형질유전자를 삽입할 수있는벡터 pUT/mini-Tn5 Km에 1개의 amber 코돈을 가지고 있는 lacZ amber 유전자를 클로닝하여 pUTlacZam1을 제작하였다(Fig. 1). 이플라스미드를대장균 DH10B 균주에 접 합방법을이용하여전달하였으며, 생성된 colony 약 400개를골
라 kanamycin 또는 ampicillin을포함하는 LB 한천배지에 각각 접종하여 kanamycin 배지에서는생장하나 ampicillin 배지에서는
생장하지 못하는클론을 3개(clones #1, #20, #301) 선별하였다
(Fig. 2A). 이들 클론은 접합에 의해서 DH10B 균주로 전달된 pUTlacZam1 중 transposon cassette 내에위치한 lacZ amber 유 전자와 Kmr가 transposon에의해 DH10B의 chromosome에삽입 되고 choromosome 안으로 삽입되지않는 β-lactamase를포함한 나머지유전자들은분해되어나타나는표현형질이다.
선별된 3개의 보고자 클론의염색체에 lacZ amber 유전자가 삽입되어있는것을확인하기위하여각각의보고자클론으로부
터 genomic DNA를 분리하였으며, 이를 주형으로 사용하여
amber 코돈을포함하는lacZ amber DNA 절편을 PCR로증폭하 여 한천겔에서 전기영동하여예상되는 약 0.3 kb 크기의 DNA
절편이 나오는가를 확인하였다(Fig. 2B). 그결과 선별된 3개의 클론에서 모두 0.3 kb band와내재 대조군 PCR인 0.5 kb 크기 의 glyceraldehyde-3-phosphate dehydrogenase (GAPDH) 유전자 절편이관찰되었으나, 형질전환이되지않은 DH10B의 genomic DNA PCR에서는 0.5 kb GAPDH 유전자절편만이나타났고, 형 질전환을위하여사용된 pUTlacZam1를주형으로한 PCR에서는 0.3 kb의lacZ amber 유전자 절편만이나타났다. 이결과는선 별된 3개의보고자클론 모두에서 lacZ amber 유전자가염색체
에삽입된형태로존재함을나타낸다.
선별된 보고자 클론들의 amber suppression 활성을비교하기 위하여 대장균의 내재 ARS인 glutaminyl-tRNA synthetase에의 해서 aminoacylation되는 E. coli amber suppressor tRNAGln (supE)을발현하는 pSupE2 벡터또는음성대조군으로빈벡터 인 pBluescript를 3개의선별된 보고자 클론에 각각 형질전환한 후, 세포추출물에 존재하는 β-galactosidase 활성을 분석하였다
(Fig. 2C). 농화배지인 LB와최소배지인 M9 배지에서 pSupE2가 형질전환된보고자클론들은 pBluescript가형질전환된클론보다 152~242배높은 β-galactosidase 활성을 나타냈으며, 3개의 보고 자클론들중 1번클론이가장높은β-galactosidase 활성증가를
보였다. 이러한 결과들은 또한 선별된 3개의 클론 모두의 β- galactosidase 활성이 amber suppression 을 통하여 나타내고있
음을보여주고있다.
선별된 3개의클론중에 amber suppression에의해가장높은 β-galactosidase 활성 증가를 나타내는 1번 클론을 선정하여
DH10B(Tn:lacZam)으로 명명하였으며, lacZ amber 유전자가대
장균염색체에몇개의 copy로삽입되어있는가를조사하기위
하여 Southern blot을 수행하였다(Fig. 2D). DH10B(Tn:lacZam)
로부터 genomic DNA를분리하여 4종류의제한효소로자른후,
한천겔에서 전기영동하였다. 이를나일론막에 옮겨 amber 코돈
부위를 포함한 lacZ유전자절편을 탐침으로사용하여 Southern
blot을 수행한결과 4종류의 제한효소를 처리한 모든 시료에서 단일밴드가나타났으며, 이로부터예측할때대장균의염색체에
삽입된lacZ amber 유전자는단일 copy인것으로예상된다.
DH10B(Tn:
lacZam)
에서 Scsuppressor tRNA
Tyr/TyrRS
쌍의amber suppression
활성 측정앞서 만들어진 in vivo amber suppression 보고자 클론인
DH10B(Tn:lacZam)가 이종 suppressor tRNA/ARS 쌍에 의해서 유도되는 amber suppression 활성을측정할 수있는가를확인하
기위하여대장균의내재 tRNA와 ARS의영향을받지않고독
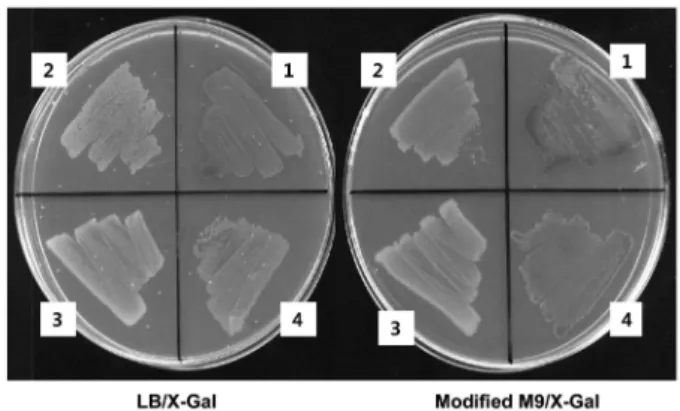
립적으로 aminoacyl-tRNA를생성할수있다고알려진 Sc amber suppressor tRNATyr/TyrRS 쌍의 amber suppression 활성을측정하 여 보았다(13). X-Gal이 포함된 LB 또는 M9 최소배지에서 pSuptRNATyr (Sc)가형질전환된 DH10B(Tn:lacZam)는흰색의세 균군락을 형성하였고, suppressor tRNATyr와 TyrRS를모두 발현
Fig. 1. Diagram of a transposable reporter plasmid pUTlacZam1. An amber stop codon was inserted at the 21st amino acid position. The arrows indicate the positions of PCR primers that were used to confirm chromosomal integration of the lacZ amber mutant gene. The primer sequences were written on the bottom.
하는 pSuptRNATyr-TyrRS (Sc)가형질전환된균주는 파란색세균 군락을 형성하였다(Fig. 3). 또한 양성 대조군으로 사용된
pSupE2 형질전환도 강한파란색 군락을생성하였으며, 음성대
조군으로사용된역방향의supE 유전자를가지고있는 pRSupE2
형질전환은 흰색의 세균군락을 형성하였다. 또한 이들의 β- galactosidase 활성을 정량적으로분석한 결과 amber suppression
을유도하지 못하는 pRSupE2 또는 pSuptRNATyr (Sc) 형질전환 에서는효소활성이전혀나타나지않았으며, pSupE2 형질전환은 강한 효소활성 (130-142 Miller unit)을, 그리고 pSuptRNATyr- TyrRS (Sc) 형질전환에서는 약한 효소활성(3.2-9 Miller unit)을 나타냈다(Table 2). 이러한 결과는 in vivo amber suppression 보
고자 클론인 DH10B(Tn:lacZam)가 매우 효과적으로 이종
suppressor tRNA/ARS 쌍의 amber suppression 활성을 정성 및 정량적으로측정할수있음을나타낼뿐만아니라 X-Gal이포함
된배지에서는세균군락의 색깔을통하여 amber suppression 활 성의유무와 강도를상대적으로 손쉽게비교할 수있음을 보여 주고있다.
DH10B(Tn:lacZam) 균주를 이용해 Sc suppressor tRNATyr과 TyrRS 쌍의 amber suppression 활성을측정해 보았을때비교적 낮은 값을 나타냈다. 이는 비록 이이종 suppressor tRNA/ARS
쌍이 대장균에서 내재 tRNA/ARS 시스템의 영향을 받지 않고
aminoacylation 활성을나타내지만그활성정도는그리높지않
은것으로 판단되며, 따라서이를이용한 in vivo비천연아미노
산삽입기술의개발은제한적일것으로예측된다.
Fig. 2. Isolation of DH10B(Tn:lacZam) reporter clones. (A) Growth properties of the selected reporter clones on different culture media. After transferring pUTlacZam1 into E. coli DH10B by a conjugation method, three positive clones were selected as described in Materials and Methods.
Their growth characteristics were tested on LB agar plates containing different antibiotics. Label 1, DH10B; 2, clone #1; 3, clone #20; 4, clone
#301; 5, E. coli S17-1 λpir containing pUTlacZam1. (B) Confirmation of chromosomal integration of the lacZ amber mutant gene by PCR.
Genomic DNAs isolated from DH10B (lane 2) or three positive clones (lanes 3~5) were amplified by PCR with primers specific for the cassette sequence as described in Materials and Methods. For control reaction, pUTlacZam1 was used for PCR template (lane 6). GAPDH PCR was used as an internal control. (C) Relative β-galactosidase activity produced by amber suppression in different DH10B(Tn:lacZam) clones. Different DH10B(Tn:lacZam) clones were transformed with either an empty pBluescript vector or pSupE2 and analysed for β-galactosidase activity in LB and modified M9 minimal media as described in Materials and Methods. The data were described as a relative fold induction of β-galactosidase activity between the empty pBluescript (basal background) and pSupE2 (amber suppression) transformants. (D) Southern hybridization of the lacZ amber mutant gene in the DH10B(Tn:lacZam) clone #1. Five µg of DH10B(Tn:lacZam) genomic DNA was digested with restriction enzymes as indicated and were subject to Southern hybridization analysis. E, EcoRV ; B, Bgl II ; S, SphI; P, PstI.
대장균에서단백질의특정위치에비천연아미노산을삽입하기 위한기술개발은일반적으로 ARS 아미노산결합부위를돌연변
이시켜서비천연 아미노산이결합할 수있도록 만듦으로써이 루어진다(3, 16, 17, 18). 하지만 이러한변형은 ARS가 가지고
있는구조적 한계로인하여 원래의천연아미노산과는전혀다 른모양의비천연 아미노산을인식하게하는데는한계가 있다.
예를들어, TyrRS를돌연변이시켜비천연아미노산을인식하게
할경우, tyrosine과유사한 구조의비천연 아미노산들로제한이
되며, 구조적으로 크게차이가나는 Arg 또는 Cys과유사한모 양을한비천연아미노산을인식하게하는데는어려움이있다.
따라서다양한형태의비천연아미노산을삽입하기위해서는다 양한 이종 suppressor tRNA/ARS 쌍이 활용되어야 하며, 본논 문의 DH10B(Tn:lacZam) 균주는대장균에서이러한선별을간편
하고빠르게수행할수있을것으로기대된다. 감사의 말
본 논문의 연구를 위하여 pUT/mini-Tn5 Km 벡터를 제공해
주신 한국외국어대학교환경학과 이규호교수께감사드린다. 본 연구는경기도지역협력연구센터(GRRC) 지원사업으로지원되었 다(단백질 내에 비천연 아미노산 삽입기술 개발을 위한
orthogonal tRNA/ARS 쌍의최적화).
참고문헌
1.김경태, 박중찬. 2001. In vivo amber suppression의 측정을 위한 reporter system의개발. 한국외국어대학교기초과학연 구 11, 93-100.
2. Bain, J.D., C.G. Glabe, T.A. Dix, A.R. Chamberlin, and E.S. Diala.
1989. Biosynthetic site-specific incorporation of a non-natural amino acid into a polypeptide. J. Am. Chem. Soc. 111, 8013-8014.
3. Bose, M., D. Groff, J. Xie, E. Brustad, and P.G. Schultz. 2006. The incorporation of a photoisomerizable amino acid into proteins in E. coli. J. Am. Chem. Soc. 128, 388-389.
4. Bradley, D., J.V. Park, and L. Soll. 1981. TRNA2Gln Su+2 mutants that increase amber suppression. J. Bacteriol. 145, 704- 5. De Lorenzo, V., M. Herrero, U. Jakubzik, and K.N. Timmis. 1990.712.
Mini-Tn5 transposon derivatives for insertion mutagenesis, pro- moter probing, and chromosomal insertion of cloned DNA in gram-negative eubacteria. J. Bacteriol. 172, 6568-6572.
6. De Lorenzo, V., I. Cases, M. Herrero, and K.N. Timmis. 1993.
Early and late responses of TOL promoters to pathway inducers:
identification of postexponential promoters in Pseudomonas putida with lacZ-tet bicistronic reporters. J. Bacteriol. 175, 6902- 6907.
7. Hohsaka, T., D. Kajihara, Y. Ashizuka, H. Murakami, and M.
Sisido. 1999. Efficient incorporation of nonnatural amino acids with large aromatic groups into streptavidin in in vitro protein syn- thesizing systems. J. Am. Chem. Soc. 121, 34-40.
8. Kowal, A.K., C. Kohrer, and U.L. RajBhandary. 2001. Twenty- first aminoacyl-tRNA synthetase suppressor tRNA pairs for pos- sible use in site-specific incorporation of amino acid analogues into proteins in eukaryotes and in eubacteria. Proc. Natl. Acad.
Sci. USA 98, 2268-2273.
9. Liu, D.R., T.J. Magliery, M. Pastrnak, and P.G. Schultz. 1997.
Engineering a tRNA and aminoacyl-tRNA synthetase for the site- specific incorporation of unnatural amino acids into proteins in vivo. Proc. Natl. Acad. Sci. USA 94, 10092-10097.
10. Liu, D.R. and P.G. Schultz. 1999. Progress toward the evolution of an organism with an expended genetic code. Proc. Natl. Acad. Sci.
USA 96, 4780-4785.
11. Mendel, D., V.W. Cornish, and P.G. Schultz. 1995. Site-directed mutagenesis with an expanded genetic code. Annu. Rev. Biophys.
Biomol. Struct. 24, 435-462.
12. Noren, C.J., S.J. Anthony-Cahill, M.C. Griffith, and P.G. Schultz.
1989. A general method for site specific incorporation of unnatural amino acids into proteins. Science 244, 182-188.
13. Ohno, S., T. Yokogawa, I. Fujii, H. Asahara, H. Inokuchi, and K.
Nishikawa. 1998. Co-expression of yeast amber suppressor tRNATyr and tyrosyl-tRNA synthetase in Escherichia coli: possi- bility to expand the genetic code. J. Biochem. 124, 1065-1068.
14. Pastrnak, M., T.J. Magliery, and P.G. Schultz. 2000. A new orthog- onal suppressor tRNA/aminoacyl-tRNA synthetase pair for evolv- ing an organism with an expanded genetic code. Helv. Chim. Acta 83, 2277-2286.
15. Steer, B.A. and P. Schimmel. 1999. Major anticodon-binding region missing from an archaebacterial tRNA synthetase. J. Biol.
Chem. 274, 35601-35606.
16. Wang, L., A. Brock, B. Herberich, and P.G. Schultz. 2001. Expand- Fig. 3. Evaluation of amber suppression through blue/white screening.
Reporter strain DH10B(Tn:lacZam) was transformed with different plasmids as indicated. Single colony of each transformation was grown on LB or modified M9 agar plates containing X-Gal. Label 1, pSupE2; 2, pRSupE2; 3, pSuptRNATyr (Sc); 4, pSuptRNATyr-TyrRS (Sc).
Table 2. Comparison of β-galactosidase activity derived by different amber suppression conditions
Plasmid β-galactosidase activity (Miller units)
LB Modified M9
None 0.4±0.1 0.4±0.00
pRSupE2 000.5±0.0 0.4±0.00
pSupE2 142.8±8.4 129.9±11.8
pSuptRNATyr(Sc) 0.4±0.0 0.4±0.00 pSuptRNATyr -TyrRS(Sc) 9.0±1.2 3.2±1.20
ing the genetic code of Escherichia coli. Science 292, 498-500.
17.Wang, L., J. Xie, and P.G. Schultz. 2006. Expanding the genetic code. Annu. Rev. Biophys. Biomol. Struct. 35, 225-249.
18. Xie, J., L. Wang, N. Wu, A. Brock, G. Spraggon, and P.G. Schultz.
2004. The site-specific incorporation of p-iodo-L-phenylalanine into proteins for structure determination. Nat. Biotechnol. 22,
1297-1301.
19. Zhang, Z., J. Gildersleeve, Y.Y. Yang, R. Xu, J.A. Loo, S. Uryu, C.H. Wong, and P.G. Schultz. 2004. A new strategy for the synthe- sis of glycoproteins. Science 303, 371-373.
(Received May 27, 2009/Accepted June 23, 2009)