—247— ISSN: 1225-8598
서 론
송사리속(Oryzias) 어류는 송사리과(Adrianichthyidae)에 속하는 소형 담수어로 24종이 아시아 전역에 널리 분포한 다(Prenti, 2008). 우리나라의 송사리속 어류는 전 연안으로 흐르는 하천 중∙하류와 서∙남해 도서 지방의 담수역 및 일본에 분포한다고 알려진 송사리(O. latipes)와 서해안으로 흐르는 하천과 섬진강 및 서해안 도서 지방의 담수역과 중 국 대륙, 타이완에 분포한다고 알려진 대륙송사리가 분류되 어 있다(김과 박, 2002).
골격형질은 분류학적 결정이나 계통을 이해하는데 많이 사용되는 형질로써(김 등, 2002), 국외에 서식하는 송사리과 어류의 경우 골격형질을 이용한 형태분석(Yabumoto and Uyeno, 1984; Werneburg and Hertwig, 2009), 종 또는 속간의 계통 비교(Parenti, 2008), 척추골 형태발달의 계통비교(Iwa-
matsu et al., 2009) 등의 많은 연구가 이루어진바 있다. 골격 형질 중 턱과 이빨 등을 포함하는 섭이기관은 어류의 먹이 생물과 섭식행동에 밀접한 연관이 있는 것으로 알려져 있 다(Gosline, 1987). 또한 섭이기관은 생태적 연관성뿐만 아 니라 어류의 성적이형(Egami, 1956; Takeuchi, 1968)이나 계 통분석(Parenti, 1987) 등의 형태적 비교 연구에 이용되기도 하며, Mbomba (1983)는 악골과 턱의 형태적 차이를 근거로 탄자니아 호수에 서식하는 시클리드과 어류의 분류학적 재 검토를 수행한 바 있다. 특히, 형태적으로 유사한 시클리드 과 어류의 턱과 이빨은 특수하게 분화된 형태를 보이는데, 이러한 유사종들의 섭이기관의 형태적 차이는 먹이생물과 관련된 종의 생태학적, 분류학적인 유용한 정보를 제공하기 도 한다(Witte and van Oijen, 1990).
우리나라에 서식하는 송사리와 대륙송사리는 미병부의 반점과 뒷지느러미의 색깔 차이로 두 종이 서로 잘 구분되 지만(Park et al., 2006), 그 외의 전반적인 외부 형태에서는 두 종이 매우 유사한 특성을 보인다. 또한 먹이에 있어서도 두 종은 모두 동물성플랑크톤을 섭식하며, 이들의 서식처
한국산 송사리속 Oryzias 2종 섭이기관의 구조적 특징
김현태∙김재구∙박종영*
전북대학교 자연과학대학 생명과학과, 전북대학교 생물다양성연구소
The Structures of Feeding Organs in Two Korean Ricefishes (Pisces, Adrianichthyidae), Oryzias latipes and O. sinensisby Hyun-Tae Kim, Jae-Goo Kim and Jong-Young Park*(Faculty of Biological Science and Institue for Biodiversity, College of Natural Science, Chonbuk National University, Jeonju, Jeollabuk-do 561-756, Korea)
ABSTRACT Structure on the feeding organs in two Korean ricefishes, Oryzias latipes and O. sinensis, was examined in the jaws and the teeth. In the jaw, O. latipes has a dent on the dorsal edge of distal arm of premaxilla (vs. straight in O. sinensis) and a alveolar arm of premaxilla running in straight line with distal arm of premaxilla (vs. ventra line in O. sinensis). O. latipes has also a dent on the posterior edge of dentary which is connected to coronoid process of dentary (vs. straight in O. sinensis). Large teeth developing only in the male, which is sexual dimorphism, developed into outside direction of the man- dible in O. latipes (vs. dorsal direction in O. sinensis). In the teeth forms, O. latipes has two types (coni- cal type and arrowhead type), whereas O. sinensis has only one type (conical type). Through this study, we confirmed that two Korean ricefishes show distinctive characteristics in the structure of the feeding organs.
Key words : Oryzias latipes, Oryzias sinensis, feeding organs, sexual dimorphism
*Corresponding author: Jong-Young Park Tel: 82-63-270-3344, Fax: 82-63-270-3362, E-mail: [email protected]
Accepted: December 24, 2012
http://www.fishkorea.or.kr
역시 수심이 얕고 물이 거의 흐르지 않는 저수지나 습지 또 는 하천의 표층에 떼를 지어 서식하는 등 생태적으로도 매 우 유사한 특성을 보인다(김과 박, 2002). 하지만 아직까지 생태적 유사종인 송사리와 대륙송사리의 섭이기관에 대한 연구는 이루어진 바 없기 때문에 이러한 연구를 수행하는 것은 서식처의 특성이 매우 유사한 이 종들의 생태적, 분류 학적 차이에 대한 정보를 제공해줄 것으로 판단된다. 따라 서 본 연구에서는 한국산 송사리속 두 종의 악골 및 이빨 을 비교하여 형태적 특성 및 차이에 관한 기초자료를 얻고 자 한다.
재료 및 방법
1. 표본의 채집
본 연구 재료인 송사리(26.4~34.5 mm SL)와 대륙송사 리(18.4~28.2 mm SL)는 2010년에서 2011년까지 우리나라 의 서해와 남해 그리고 동해로 흐르는 여러 하천 중 8개 지점의 농수로에서 족대(망목 1×1 mm)를 이용하여 채집하 였다 (Table 1, Fig. 1). 채집된 개체는 각각 10% Formalin에 고정하여 실험에 사용하였다.
2. 실험방법
실험에 이용된 개체들은 박과 최(1998)의 방법에 따라 성적이형이 나타나는 크기에서 선별하였고, 골격 명칭은 Parenti (1981, 1987)와 이 등(1998)에 따랐다.
고정된 표본은 Potthoff (1984)의 투명골격염색법에 따라 실험한 후 글리세린에 고정하기 전 주상악골과 전상악골, 하악골을 두부에서 분리하였다. 그리고 분리된 뼈들을 다시 5% KOH용액에 24시간 담가서 뼈를 제외한 조직을 완전히 제거한 후 주사전자현미경(JEOL JSM-6400, Japan) 관찰을 위해 0.1 M cacodylate buffer (7.4)로 조정된 2.5% glutaral- dehyde로 4�C에서 4시간 전고정 후 1% OsO4로 90분간 후
고정하였다. 동결건조법으로 건조한 후 OsO4로 코팅하여 미세구조를 관찰하였다.
결 과
1. 악골(Jaw bones)의 형태
1) 전상악골(Premaxilla)
전상악골은 공통적으로 앞쪽(tip of mouth)부터 상행돌기 (ascending process of premaxilla), 중앙부(distal arm of pre- maxilla, dapm) 그리고 후반부(alveolar arm of premaxilla, alv) 로 나눌 수 있었으며, 상행돌기는 발달하지 않았다(Fig. 2).
두 종간의 형태적 차이에서 송사리는 중앙부의 등 쪽 가 장자리에 움푹 들어가는 홈이 존재하였으며 후반부의 위치 는 중앙부와 일직선상으로 존재하였다(Fig. 2A, B). 그러나 대륙송사리는 중앙부의 등 쪽 가장자리가 일직선이었으며 후반부의 위치는 중앙부보다 배 쪽으로 존재하였다(Fig. 2 C, D).
2) 주상악골(Maxilla)
주상악골에서는 형태적 차이를 보이지 않았으며 공통적으 로 이빨이 발달하지 않았다. 이들의 형태는 앞쪽부터 아래 쪽으로 향하는 배 쪽 부분(ventral arm of maxilla, vam)과 일 직선상으로 배열되어 있는 뒷부분(posterior ramus of maxilla, prm)으로 구성되어 있다(Fig. 3).
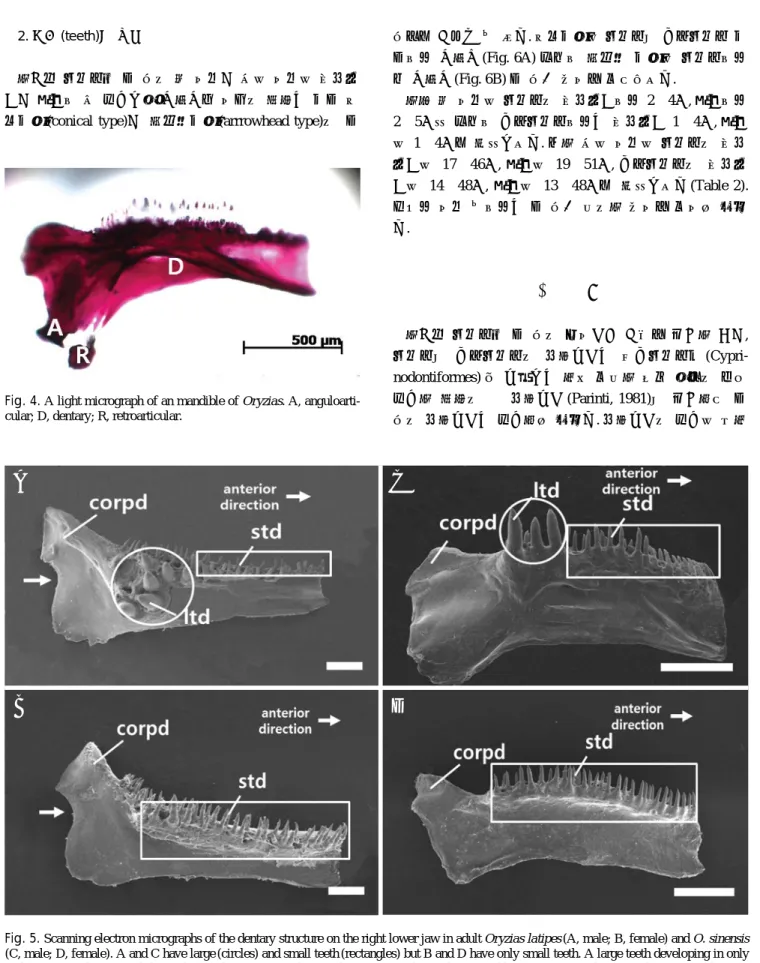
3) 하악골(Mandible)
하악골은 공통적으로 치골(dentary), 관절골(anguloarticu-
Fig. 1. A map showing the collecting sites of Oryzias sinensis (Cir- cles) and O. latipes (Triangles) in Korea. The localities are described in Table 1.
38�
36�
34�
124� 126� 128� 130�
0 100km
Table 1. The collecting sites of Oryzias sinensis and O. latipes in Korea
Collecting site Species
Stations 1 Unyeon, Namdong, Incheon-si
Stations 2 Songchoen, Dukjin, Jeonju-si, Jeollabuk-do
Stations 3 Taechang, Taein, Jeongeup-si, Jeollabuk-do O. sinensis Stations 4 Negyo, Hampyung, Hampyung-gun,
Jeollanam-do
Stations 5 Sinjido, Sinji, Wando-gun, Jeollanam-do Stations 6 Haksung, Heunghae, Pohang-si,
Gyeongsangbuk-do
O. latipes Stations 7 Wonhwang, Byeonggok, Youngduk-gun,
Gyeongsangbuk-do
Stations 8 Keumjeon, Okgye, Gangreun-si, Gangwon-do
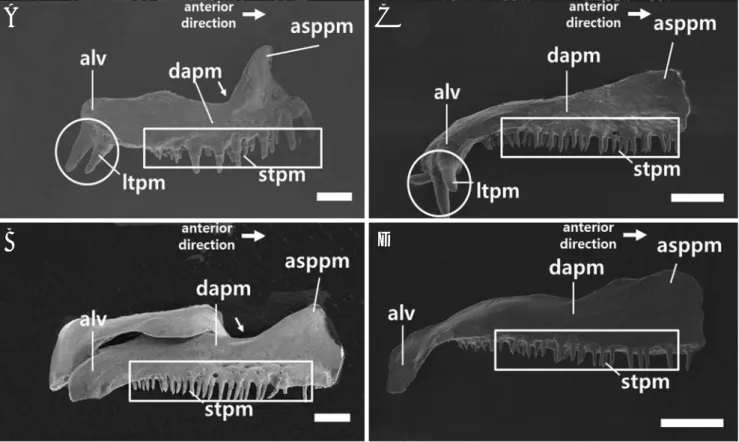
lar), 각골 (retroarticular)로 구성되어 있었으며 (Fig. 4) 그 중 치골의 형태에서 두 종간 차이를 보였다. 송사리에서는 구상돌기(coronoid process of dentary) 아래로 이어지는 치 골 뒷가장자리에 움푹 들어가는 홈(dent)이 존재하였으며, 수컷에서만 나타나는 큰 이빨이 뼈의 측면방향 (outside
direction)으로 발달하는 성적이형이 관찰되었다 (Fig. 5A, B). 그러나 대륙송사리에서는 구상돌기 아래로 이어지는 치 골 뒷가장자리가 일직선이었으며 큰 이빨은 등 쪽 방향
(dorsal direction)으로 발달하였다(Fig. 5C, D).
Fig. 2. Scanning electron micrographs of the premaxillary structure of the right upper jaw in adult Oryzias latipes (A, male; B, female) and O.
sinensis (C, male; D, female). The arrows are dent curved inward on dorsal edge in premaxilla. Abbreviations: alv, alveolar arm of premaxilla;
appm, ascending process of premaxilla; dapm, distal arm of premaxilla; ltpm, large teeth of premaxilla (circles); stpm, small teeth of premaxilla (rectangles). Bars indicate 200μm, respectively.
A C
B D
Fig. 3. Scanning electron micrographs of the maxillary structure of the right upper jaw in males of Oryzias. A, O. latipes; B, O. sinensis. Abbrevia- tions: prm, posterior ramus of maxilla; vam, ventral arm of maxilla. Bars indicate 200μm, respectively.
A B
2. 이빨(teeth)의 형태
한국산 송사리속 두 종의 큰 이빨과 작은 이빨은 전상악 골과 치골에 잘 발달되어 나타나며 이들의 형태는 모두 원 뿔모양(conical type)과 화살촉모양(arrrowhead type)의 두
종류로 구분될 수 있다. 원뿔모양은 송사리와 대륙송사리 모 두에서 나타난(Fig. 6A) 반면에 화살촉 모양은 송사리에서 만 나타나(Fig. 6B) 두 종간 차이를 보여주었다.
한편 큰 이빨은 송사리의 전상악골에서 2~4개, 치골에서 2~5개인 반면에 대륙송사리에서는 전상악골 1~4개, 치골 은 1~4개로 확인되었다. 또한 작은 이빨은 송사리의 전상 악골은 17~46개, 치골은 19~51개, 대륙송사리의 전상악 골은 14~48개, 치골은 13~48개로 확인되었다(Table 2).
따라서 이빨 수에서는 두 종간 유의한 차이를 보이지 않았 다.
고 찰
한국산 송사리속 두 종의 섭이기관 구조를 비교한 결과, 송사리와 대륙송사리의 상행돌기는 열대송사리목 (Cypri- nodontiformes) 중 돌출되는 턱을 보유한 일부 어류의 매우 발달한 형태의 긴 상행돌기 (Parinti, 1981)와 비교하여 두 종의 상행돌기는 발달하지 않았다. 상행돌기의 발달은 윗턱 Fig. 4. A light micrograph of an mandible of Oryzias. A, anguloarti-
cular; D, dentary; R, retroarticular.
Fig. 5. Scanning electron micrographs of the dentary structure on the right lower jaw in adult Oryzias latipes (A, male; B, female) and O. sinensis (C, male; D, female). A and C have large (circles) and small teeth (rectangles) but B and D have only small teeth. A large teeth developing in only male, appear to be a sexual dimorphism. The arrows are dent on the posterior edge of dentary. Abbreviations: corpd, coronoid process of dentary;
ltd, large teeth of dentary; std, small teeth of dentary. Bars indicate 200μm, respectively.
A C
B D
의 돌출과 매우 밀접한 관련이 있기 때문에 발달하지 않은 상행돌기를 나타낸 한국산 송사리속 어류 두 종은 윗턱을 돌출 시키지 못할 것으로 생각되며 이러한 결과는 돌출되 지 않는 턱을 가진 동갈치목 (Alexander, 1967)에서 보여지 는 일반적인 특징으로 판단된다.
Barel (1983)은 섭이양상(feeding apparatus)을 먹이를 입 안에서 단단히 씹지 않고 흡입하는 양상(sucking)과 턱 힘 을 이용하여 입안에서 씹는 양상(biting) 등 2가지로 설명 하였는데, 그 중 sucking은 전상악골의 돌출을 이용하여 먹 이를 포획한다고 하였다. 또한 Mok and Munro (1991)는 O.
javanicus 턱의 비돌출 현상은 활동적인 먹이생물을 sucking 하는 것 보다 물속에서 분해된 유기물과 같은 비활동적인 먹이를 섭식하는데 적합하게 적응된 결과라고 보고하였다.
따라서 한국산 송사리속 두 종은 돌출되는 턱을 이용한 sucking보다는 biting의 양상에 가깝다고 생각되며 농경지 같은 유기물이 많은 서식환경에서 활동성이 약한 유기물과 같은 먹이를 섭식하는데 적응된 결과로 생각된다.
그 밖에 전상악골의 후반부 위치와 윗가장자리 모양, 상행 돌기 발달양상 그리고 치골의 뒷가장자리 모양과 큰 이빨의 발달방향에서 두 종간에 차이를 보였다. Kwun et al. (2010)
는 Coilia brachygnathus와 C. nasus 두 종에서 나타나는 전 상악골의 형태적 차이를 이용하여 종간에 분류학적 형질로 사용하였다. 따라서 본 연구에서 나타난 두 종간의 차이는 확실한 종간 분류학적 형질로 이용될 수 있을 것으로 사료 된다.
전상악골과 치골에 발달한 이빨의 모양은 송사리에서 2 types (원뿔모양, Conical type; 화살촉모양, Arrowhead type), 대륙송사리는 1 type (원뿔모양)만이 관찰되었다. 원뿔모양은 adrianichthys을 제외한 송사리과 어류에서 나타나는 이빨 의 형태이지만 화살촉모양은 보고된 바 없다. 이러한 결과 를 바탕으로 송사리과에서 이빨의 모양은 분류학적 형질로 사용되기 때문에 송사리에서 보여지는 화살촉모양의 이빨 은 이 종만이 가지는 분류학적 특징으로 판단된다(Parenti, 2008).
한국산 송사리속 두 종에서 암수간 전상악골과 치골의 큰 이빨 발달유무에서 차이를 보였다. Egami and Ishii (1956)는 일본산 송사리에서 나타나는 큰 이빨 유무의 암수간 차이 를 2차 성징에 따른 성 호르몬의 차이 때문이라고 보고한 바 있는 데, 아직까지 큰 이빨이 어떠한 기능을 하는 지에 대해서는 알려진 바 없다. 한편 Parenti (2008)는 암수간 큰 Fig. 6. Scanning electron micrographs of small teeth in Oryzias. A, conical type; B, arrowhead type. Bars indicate 30μm, respectively.
A B
Table 2. Counts of the teeth in Oryzias
Locality Oryzias sinensis Oryzias latipes
Incheon Jeonju Jeongeup Hampyeong Sinjido Pohang Youngduk Gangreung
Individual numbers 12 19 20 13 13 20 10 17
Large teeth of premaxilla 3 2 3 2 3 3 3 3
(2~4) (1~4) (2~3) (1~4) (3~4) (2~4) (2~3) (2~4)
Small teeth of premaxilla 33 31 25 32 34 32 31 31
(24~41) (25~48) (14~32) (23~43) (28~42) (26~42) (26~38) (17~46)
Large teeth of dentary 3 2 2 3 3 4 3 3
(2~3) (1~4) (1~4) (2~3) (2~5) (2~4) (2~3) (2~4)
Small teeth of dentary 37 31 28 29 43 34 34 36
(26~48) (21~46) (13~40) (15~39) (33~63) (21~47) (22~45) (19~51)
Mean±SD, (range)
이빨의 유무가 송사리속 어류들 간에 차이를 보인다고 하 였는데, 송사리는 암수 모두에서 나타나며 대륙송사리는 수 컷에서만 나타난다고 하였다. 하지만 본 연구에서는 이들 종 모두 수컷에서만 나타나 위의 연구와 차이를 보였다.
이상과 같이 한국산 송사리속 어류 두 종의 섭이기관 구 조는 전상악골과 치골, 이빨에서 형태적 차이를 보이고 있 는데, 이러한 결과는 먹이생물의 종류와 섭식행동에 적응한 종의 특이성으로 판단되며 두 종의 분류학적 차이를 확인 하는 중요한 자료로 사료된다.
요 약
한국산 송사리속 2종에 대한 섭이기관의 구조적 차이를 알아보기 위하여 악골 및 이빨 구조를 조사하였다. 악골에 서 송사리 Oryzias latipes는 전상악골 중앙부의 등 쪽 가장 자리에 움푹 들어가는 홈이 존재하였으나 대륙송사리 O.
sinensis는 일직선이었다. 후반부의 위치에서 송사리는 중앙 부와 일직선상에 존재하였으나 대륙송사리는 배 쪽에 존재 하였다. 또한 송사리는 하악골의 구상돌기 아래로 이어지는 치골 뒷가장자리에 움푹 들어가는 홈이 존재한 반면에 대륙 송사리에서는 뒷가장자리가 일직선 모양을 하였다. 송사리 는 성적이형으로서 수컷에서만 나타나는 큰 이빨이 하악골 의 측면 방향으로 발달하였고, 대륙송사리는 등 쪽 방향으 로 발달하였다. 한편 송사리의 이빨형태는 원뿔모양과 화살 촉모양 2 types이 나타난 반면에 대륙송사리에서는 원뿔 모 양 1 type만 나타나는 특징을 보였다. 이상과 같이 한국에 분포하는 2종의 송사리는 섭이기관에서 뚜렷한 종간의 특 징을 잘 보여 주었다.
인 용 문 헌
김익수∙박종영. 2002. 한국의 민물고기. 교학사, 465pp.
김익수∙안정현∙김병직. 2002. 한국산 새코미꾸리 Koreocobitis 어류의 골학적 연구. 한국어류학회, 14: 19-28.
박두신∙최신석. 송사리와 대륙송사리의 산란, 난발생 및 자어의 성장. 1998. 한국하천호수학회, 32: 109-118.
이장욱∙김용억∙백철인∙김종빈∙김진구∙황선재. 1998. 인도 네시아 아라푸라해의 하스돔속 Pomadasys (하스돔과, 농 어목) 어류 3종의 섭이구조. 한국어류학회, 10: 32-39.
Alexander, R.M. 1967. The functions and mechanism of the protru- sible upper jaws of some acanthopterygian fish. J. Zool.
London, 151: 43-64.
Barel, C.D.N. 1983. Towards a constructional morphology of cichlid fishes (Teleostei, Perciformes). Neth. J. Zool., 33: 357-424.
Egami, N. 1956. Notes on sexual difference in size of teeth of the fish, Oryzias latipes. Japan J. Zool., 12: 65-69.
Egami, N. and S. Ishii. 1956. Sexual differences in the shape of some bones in the fish, Oryzias latipes. J. Fac. Sci. Tokyo Univ., 7: 563-571.
Gosline, W.A. 1987. Jaw structures and movements in higher teleo- stean fishes. Japan. Japanese Ichthyol., 34: 21-32.
Iwamatsu, T., M. Sato and K. Nakane. 2009. Development of the first vertebra in Oryzias latipes and its morphology in beloni- formes and cyprinodontiformes. Bulletin of Aichi Univ.
Educat., 58: 69-79.
Kwun, H.J., Y.H. Kim, J.B. Kim, C.H. Jeong and J.K. Kim. 2010.
One unusual species, Coilia sp. (Engraulidae, Pisces) from the Yellow Sea. Animal Cells and Systems, 14: 137-145.
Mbomba, N.B. 1983. Comparative morphology of the feeding appa- ratus in cichlidian algal feeders of lake tanganyika. Afr. Stud.
Monogr., 3: 1-23.
Mok, E.K.M and A.D Munro. 1991. Observation on the food and feeding adaptations of four species of small pelagic teleosts in streams of the sungei buloh mangal, singapore. Raffles Bull. Zool., 39: 235-257.
Parenti, L.R. 1981. A phylogenetic and biogeographic analysis of cyprinodontiform fishes. Bull. American Mus. Nat. Hist., 168: 335-557.
Parenti, L.R. 1987. Phylogenetic aspects of tooth and jaw structure of the medaka, Oryzias latipes, and other beloniform fishes.
J. Zool., Lond., 211: 561-572.
Parenti, L.R. 2008. A phylogenetic analysis and taxonomic revision of ricefishes, Oryzias and relatives (Beloniformes, Adriani- chthyidae). Zool. J. Linn. Soc., 154: 494-610.
Park, M.K., I.S. Kim and J.Y. Park. 2006. Morphological characters and geographical distributions of Korean ricefish. Korean J.
Ichthyol., 18: 163-169.
Potthoff, T. 1984. Clearing and staining techniques. In: Moser, H.G., W.J. Richards, D.M. Cohen, M.P. Fahay, A.R. Kendall, Jr.
and S.L. Richardson. (eds.), Ontogeny and systematics of fishes. Am Soc. Ichthyol. Herpetol. Spec. Publ. 1., pp. 35- 37.
Takeuchi, K. 1968. Inhibition of large distal tooth formation in male medaka, Oryzias latipes, by estradiol. Experientia., 24: 1061- 1062.
Werneburg, I. and S.T. Hertwig. 2009. Head morphology of the rice- fish, Oryzias latipes (Teleostei: Beloniformes). J. Morphol., 270: 1095-1106.
Witte, F. and M.J.P. van Oijen. 1990. Taxonomy, ecology and fich- ery of Lake Victoria haplochromine trophic groups. Zool.
Verh. Leiden, 262: 1-47.
Yabumoto, Y. and T. Uyeno. 1984. Osteology of the ricefish, Oryzias latipes. Bull. Kitakyushu Mus. Nat. Hist., 5: 143-161.