21(4) : 268-272 (2015)
http://dx.doi.org/10.20307/nps.2015.21.4.268
268
Anti-inflammatory Effect of Dactyloquinone B and Cyclospongiaquinone-1 Mixture in RAW264.7 Macrophage and ICR Mice
Dong-Sung Lee
1,#, In Hyun Hwang
2,#, Nam-Kyung Im
3, Gil-Saeng Jeong
3,*, and MinKyun Na
4,*
1
Department of Biomedical Chemistry, Konkuk University, Chungju 380-701, Korea
2
Department of Chemistry, University of Iowa, Iowa City, IA 52242, United States
3
College of Pharmacy, Keimyung University, Daegu 704-701, Korea
4
College of Pharmacy, Chungnam National University; Daejeon 305-764, Korea
Abstract – Sesquiterpene-quinone is a class of secondary metabolites frequently encountered from marine sponge.
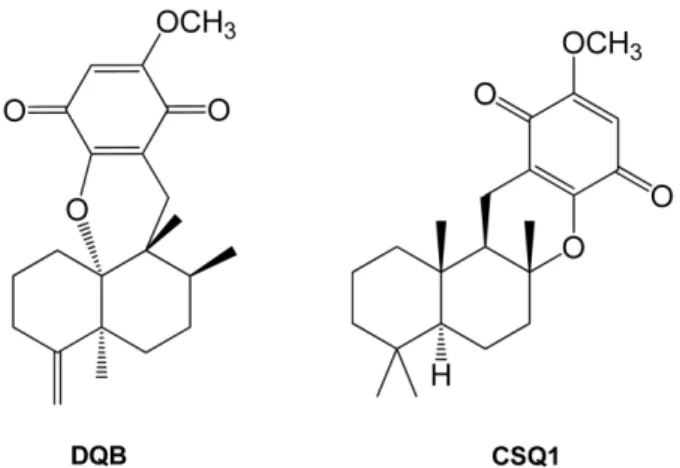
The present study was designed to examine the anti-inflammatory action of sponge-derived dactyloquinone B (DQB) and cyclospongiaquinone-1 (CSQ1) mixture using lipopolysaccharide (LPS)-induced inflammatory responses.
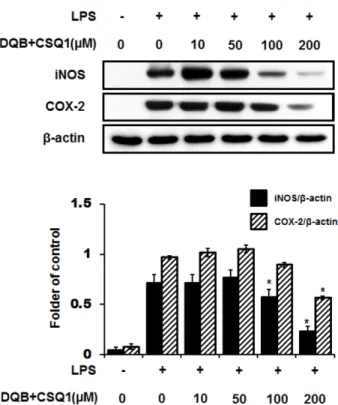
We measured the production of nitric oxide (NO), tumor necrosis factor-alpha (TNF-α), interleukin-1β (IL-1β), and interleukin-6 (IL-6) and expression of inducible nitric oxide synthase (iNOS) and cyclooxygenase-2 (COX-2) protein. TNF-α, IL-1β, and IL-6 production, which increased by treatment with LPS, were significantly inhibited by DQB and CSQ1 mixture. It also decreased the production of NO production, and iNOS and COX-2 expression. Furthermore, it reduced 12-O-tetradecanoylphorbol 13-acetate (TPA)-induced ear edema of ICR mice.
These results demonstrate that sesquiterpene-quinone, DQB and CSQ1 mixture, might serve as a chemical pipeline for the development of anti-inflammatory agent.
Keywords – Dactyloquinone B, Cyclospongiaquinone-1, Anti-inflammatory effect, Ear edema model
Introduction
Marine sponge has been served as a prolific source of diverse secondary metabolites.
1In particular, sesquiterpene- quinone is a class of compounds frequently encountered as bioactive marine natural products.
2A wide range of bioactivities have been described from this class, which included antimicrobial, anti-HIV, anti-inflammatory, and cytotoxic activities, and thereby a lot of synthetic attempts have been made for these compounds.
2Ilimaquinone is one of the most well-known sesquiterpene-quinones comprised of sesquiterpene and benzoquinone subunits,
3,4and its mechanism of action associated with Golgi membrane was reported.
5As a part of the chemical investigation of marine sponge, a series of cyclospongia- quinones were isolated from Stelospongia conulata.
6Cyclospongiaquinones contain a characteristic ether linkage between the sesquiterpene and quinone units, forming an additional ring system relative to that of most other sesquiterpene-quinones such as ilimaquinone.
6Recently, a short synthetic strategy for cyclospongiaquinone-1 was developed using commercial diterpene alcohol.
7Another set of example showing the same type of ether connection is dactyloquinones obtained from Dactylospongia elegans, but the skeleton of their sesquiterpene unit is rearranged as was the case for ilimaquinone.
8-10Despite the unique structural feature of these series of compounds, to our knowledge, previous reports of their bioactivity have been limited to moderate cytotoxicity of dactyloquinone B, neodactyloquinone, and cyclospongiaquinone-2.
10-12Inflammatory response is a defense mechanism of the body to various stimuli, in which macrophages play an important role in regulating the reaction via production of pro-inflammatory mediators and cytokines, such as nitric oxide (NO), tumor necrosis factor- α (TNF-α), and interleukin-1 β (IL-1β).
13Prolonged or deregulated inflam- matory responses can lead to various diseases, including arthritis, hepatitis, septic shock syndrome, neurodegenerative disorders, and sepsis.
14Accordingly, many studies have focused on searching for the treatment of inflammatory-
*Author for correspondence
Professor Gil-Saeng Jeong, College of Pharmacy, Keimyung Uni- versity, 1095 Dalgubeol-daero, Daegu 704-701, Republic of Korea Tel: +82-53-580-6649; E-mail: [email protected]
Professor MinKyun Na, College of Pharmacy, Chungnam National University, 99 Daehak-ro,Yuseong, Daejeon 305-764, Republic of Korea
Tel: +82-42-821-5925; E-mail: [email protected]
#