Vol. 11, No. 2, June, 2004
인류의 신장 혈관간세포에서 유착분자 발현과 Chemokine 발현에 미치는 Th1 Cytokine과
Th2 Cytokine의 상호 작용
한양대학교 의과대학 내과학교실*, 류마티스병원*, 류마티즘 연구소**, 비뇨기과학교실***, 에모리대학교 의과대학 내과학교실†
나경선*․강태영*․이진숙**․박용욱*
†․이혜순*․엄완식*
김태환*․전재범*․배상철*․이춘용***․유대현*
= Abstract =
Interaction between Th1 Cytokine and Th2 Cytokine on the Expression of Chemokine and Adhesion Molecule in Human Mesangial Cells
Kyoung-Sun Na, M.D.*, Tae-Young Kang, M.D.*, Jin-Sook Lee, B.S.**, Yong-Wook Park, M.D.*†, Hye-Soon Lee, M.D.,*, Wan-Sik Uhm M.D.*,
Tae-Hwan Kim, M.D.*, Jae-Bum Jun, M.D.*, Sang-Cheol Bae, M.D.*, Tchun Yong Lee, M.D.***, Dae-Hyun Yoo, M.D.*
Division of Rheumatology, Department of Internal Medicine*, Institute of Rheumatology**, Department of Urology***, Hanyang University College of Medicine, Seoul, Korea,
Lowance Center for Human Immunology, Department of Medicine, Emory University School of Medicine, Atlanta, USA†
Purpose: Predominance of IFN-γ over Th2 cytokines is evident in proliferative lupus nephritis.
Th2 cytokines such as IL-10, or IL-4 have been known to suppress Th1 cytokine driven pro-inflammatory responses. However a combination of cytokines can exert variable effects during the evolution of autoimmune diseases depending on the various factors, such as the stage of diseases or local versus systemic expression. To determine whether Th2 cytokines play a pro- or anti-inflammatory role in the local glomerular inflammation induced by IFN-γ, we studied the effect of IL-10 and IL-4 on the expression of chemokines and adhesion molecules in human mesangial cells stimulated with IFN-γ.
ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ
<접수일:2003년 8월 20일, 심사통과일:2004년 3월 15일>
※통신저자:유 대 현
서울시 성동구 행당동 산 17
한양대학교 의과대학 내과학교실, 류마티스병원
Tel:02) 2290-9202, Fax:02) 2298-8231, E-mail:[email protected]
서 론
사이토카인은 면역 세포의 분화와 면역 조절에 중 요한 역할을 하며, 자가면역 질환의 발생과 밀접한 관련이 있는 것으로 알려져 있다1). 사이토카인은 이 를 분비하는 보조 T 림프구(T helper cell)에 따라, T helper 1 (Th1)과 T helper 2 (Th2) 사이토카인으로 나눌 수 있는데, Th1 세포는 interleukin-2 (IL-2), interferon-γ (IFN-γ)를 분비하고 세포독성 작용, 지 연형 과민반응, 대식세포 활성화 등의 세포 매개성 면역반응에 관여한다. 이에 반해 Th2 세포는 interle- ukin-4 (IL-4), interleukin-5 (IL-5), interleukin-6 (IL-6), interleukin-10 (IL-10), interleukin-13 (IL-13)을 분비하 고, 항체 생성을 유도하는 체액 매개성 면역반응에 관여한다2).
전신성 홍반성 루푸스(이하 루푸스)의 발병 기전 에서 Th1과 Th2 사이토카인 간의 불균형은 많은 논 란의 대상이 되었다. 루푸스는 다클론성 B세포의 활 성과 병원성 자가항체의 과잉생성을 특징으로 하기 때문에 Th2 면역반응이 지배적인 자가면역질환으로 여겨져 왔다3,4). 그러나 루푸스 동물모델(lupus prone mice)에서 증식성 사구체신염의 유발과 진행에 IFN- γ가 결정적인 역할을 하며, 증식성 루푸스 신염 환 자의 말초혈액과 신조직에서도 Th1 면역반응이 우
세하다는 최근의 보고들을 통해 세포 매개성 면역반 응을 촉진하는 IFN-γ가 루푸스의 병인, 특히 증식성 사구체신염의 발생과 진행에 보다 중요한 역할을 함 이 알려졌다5-11). 그러나 IFN-γ의 중추적인 역할에 대한 증거들과 마찬가지로, IL-4, IL-10 또한 자가면 역 사구체신염의 진행에 기여한다는 루푸스 동물 실 험의 결과들은 루푸스는 Th1 또는 Th2 중 어느 한 쪽의 면역반응에 전적으로 의존하지 않고 Th1, Th2 사이토카인이 복합적으로 작용하는 자가면역질환임 을 시사한다12-14).
루푸스 신염의 발생은 병원성 자가항체의 생산, 면역복합체의 침착과 보체의 활성화로부터 기인하는 반면, 신 손상의 진행은 대식세포와 T세포의 신장조 직 내의 침윤이라는 일반적인 경로를 따르는데, 이 러한 과정에서 chemotactic factor와 유착분자가 중요 한 역할을 하게 된다15). 대표적인 chemotactic factor 인 monocyte chemotactic protein-1 (MCP-1)은 IL-1, TNF-α, 또는 IFN-γ에 의해 혈관간세포, 신세뇨관 상피세포와 같은 신장내의 실질세포에서 생성되며, intercellular adhesion molecule-1 (ICAM-1), vascular cell adhesion molecule-1 (VCAM-1) 등의 유착분자의 발현은 루푸스 환자와 동물모델에서 혈관간세포와 신세뇨관 상피세포에서 증가되어 있다16-18). 사구체 내에서 분비된 사이토카인과 사구체 내에서 일어난 세포간 상호 작용을 통한 자극은 혈관간세포로 하여 Methods: Human mesangial cells were obtained from 3 patients undergoing nephrectomy.
MCP-1, RANTES, ICAM-1, and CD40 expressions were examined in response to various cytokine stimulation; IFN-γ, IL-10, IL-4, IFN-γ+IL-10, IFN-γ+IL-4, and IFN-γ+IL-10+IL-4, respectively. Expression of MCP-1 mRNA was analyzed by RT-PCR and the levels of MCP-1 and RANTES in the culture supernatants were measured using an enzyme immunoassay.
ICAM-1 and CD40 surface expressions were analyzed by flow cytometry.
Results: IFN-γ increased basal mRNA expression of MCP-1, and IL-10 strongly enhanced the level of MCP-1 mRNA and protein expressions induced by IFN-γ. IL-4 did not affect significantly on the MCP-1 expression induced by IFN-γ. Although IFN-γ increased the concentration of RANTES, IL-10 and IL-4 did not affect IFN-γ induced RNTES expression.
Increased expression of ICAM-1 and CD40 induced by IFN-γ were not down-regulated by IL-10 or IL-4.
Conclusion: Taken together, IL-10 appears to augment local inflammatory reaction though the up-regulation of MCP-1 from mesangial cells in the presence of IFN-γ rather than inhibit inflammatory response in the pathogenesis of proliferative glomerulonephritis.
ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ Key Words: Cytokines, Chemokines, Adhesion molecules, Mesnagial cell
금 MCP-1, regulated upon activation, normal T cell expressed and secreted (RANTES), interleukin-8 등의 chemokine과 ICAM-1, VCAM-1 등의 유착분자의 발 현을 유도한다16-20). 그러므로, 혈관간세포는 대식세 포, T세포와의 상호작용을 통해 사구체내로의 염증 세포의 이동과 침윤을 촉진시켜 사구체 염증과 손상 을 증폭시키는 데 중요한 역할을 한다고 알려져 있다.
신염 환자의 사구체 내에는 IFN-γ와 함께 IL-10과 IL-4 같은 Th2 사이토카인의 발현도 동시에 관찰되며11), IL-10과 IL-4는 Th1 면역반응의 억제와 함께 단핵구 세포와 대식세포의 사이토카인, chemokine의 분비 억제 등의 항염증 기능을 하는 것으로 알려져 있
다21-23). 그러나 Th2 사이토카인의 전신적인 작용 이
외에 신장 조직 내에서 IFN-γ에 의해 유도된 혈관 간세포의 chemokine과 유착분자의 발현에 Th2 사이 토카인이 어떤 영향을 주는지는 연구된 바 없다.
혈관간세포의 chemokine과 유착분자의 발현에 대 한 IFN-γ와 IL-10, IL-4 간의 상호작용을 규명하는 것은 증식성 루푸스 신염의 병인에서 국소적인 사구 체내의 염증반응에 대한 사이토카인의 역할에 대한 이해를 높이고, 이에 대한 적절한 치료 목표를 설정 하는 데 도움을 줄 것으로 기대된다. 이에 연구자는 인간 혈관간세포를 이용하여 IL-10과 IL-4가 IFN-γ 에 의해 유도된 MCP-1, RANTES, ICAM-1과 CD40 의 발현에 미치는 영향을 알아보고자 하였다.
대상 및 방법 1. 혈관간세포의 배양
신장 종양 환자의 신절제술로 얻어진 신조직으로 부터 정상 신피질을 분리하고, 이를 가위와 syringe plunger로 분쇄한 후에 220μm, 75μm의 stainless steel mesh에 차례로 통과시켜 75μm mesh 위에 모 여진 사구체를 RPMI 1,640 배양액 (Gibco Ltd, Paksley, UK)으로 세척하였다. 수확된 사구체는 37oC 에서 1 mg/mL의 type IV collagenase (Sigma, St.
Louis, MO)로 처리하면서 5분마다 현미경 시야에서 효소의 효과를 관찰한 다음, fetal bovine serum (우혈 청; Gibco Ltd, Paksley, UK)을 첨가하여 collagenase 의 작용을 중지시키고, 75 cm2의 조직배양 플라스크 로 옮겨, 20% 열 불활성형 우혈청, 2 mM L-gluta-
mine, penicillin (100 IU/mL), streptomycin (100μg/
mL), insulin (5μg/mL; Sigma, St. Louis, MO)이 함유 된 RPMI 1,640 배양액으로, 37oC, 5% CO2 배양기에 서 배양하였다. 배양된 세포가 단층을 형성하면, 우 혈청이 없는 RPMI로 2회 세척한 후 0.05% trypsin- EDTA용액으로 처리하여 계대 배양하였다.
배양된 세포는 역상현미경을 통한 세포 형태의 관 찰과 anti-CD31, anti-cytokeratin, anti-vimentin, anti- smooth muscle actin 단클론 항체(Dako corp., Carpin- teria, CA)와 avidin-biotin peroxidase complex 염색을 통한 면역조직화학법으로 혈관간세포임을 확인하였 다. 배양된 세포는 vimentin과 smooth muscle actin에 양성, desmin, CD31, cytokeratin에 음성을 보여 내피 세포와 상피세포가 배제된 혈관간세포임을 알 수 있 었고, 형태학적으로 혈관간세포의 특징인 끝이 길고 각이 진 별 모양의 세포를 관찰할 수 있었다(그림 1).
본 실험에서는 3명의 환자에게서 분리한 인간 혈관 간세포주를 3∼6회 계대 배양한 다음 사용하였다.
2. 사이토카인 자극
사이토카인 자극에 따른 효과를 비교하기 위하여 배양 플라스크에서 trypsin-EDTA로 처리하여 수확한 혈관간세포를 각 well당 1-2×105의 세포를 6 well plate에 분주하였다. 이를 사이토카인을 처리하지 않 은 대조군, IFN-γ (250 U/mL), IL-10 (10 ng/mL), IL-4 (10 ng/mL), IFN-γ+IL-10, IFN-γ+IL-4와 IFN-γ+
IL-10+IL-4으로 처리한 총 7개 군으로 분리하여 배 양하였다. rhIFN-γ, rhIL-10, rhIL-4는 R&D Systems (Minneapolis, MN)에서 구입하였다.
3. MCP-1에 대한 역전사 중합효소반응법(reverse transcription-polymerase chain reaction; RT-PCR)
MCP-1의 mRNA 발현을 관찰하기 위하여 혈관간 세포를 우혈청이 없는 단순 배양액에 24시간에서 48 시간 동안 배양시킨 후에 신선한 serum free 배양액 으로 교환하고, 사이토카인 처리 4시간 후에 세포를 채취하여 RNA를 분리하였다. RNAzol (Tel-Test Inc., Friendswood, TX)을 이용하여 추출한 3μg의 total RNA에 oligodT primer, M-MuLV reverse transcriptase, RNasin (Promega, Madison, WI), 25 mM dNTP (Roche Diagnostics, Ottweiler, Germany)를 첨가하여 42oC에서
60분간, 95oC에서 5분간 반응시켜 cDNA를 합성하였 다. cDNA의 중합효소반응은 2μL의 10X 반응완충 액, 1.6μL의 4 dNTP (10 mM), 1 unit의 Taq DNA polymerase (Roche Diagnostics, Ottweiler, Germany), 0.5μM/L의 primer가 포함된 총 20μL의 반응액을 GeneAmp PCR system 9,600 (PerkinElmer Co., Wellesley, MA)을 사용하여 시행하였으며, mRNA의 정량을 위하여 β-actin을 대조군으로 사용하였다. PCR 에 사용한 각각의 primer의 염기서열은 다음과 같다.
MCP-1 forward primer 5’
-CCAATTCTCAAACTGAAGCTCGCAC-3’
backward primer 5’
-GTTAGCTGCCAGATTCTTGGGTTGTG-3’
β-actin forward primer 5’
-CTCCTTAATGTCACGCACGATTTC-3’
backward primer 5’
-GTGGGGCGCCCCAGGCACCA-3’
PCR 반응의 조건은 denaturation을 위하여 94oC에
서 45초간, annealing을 위하여 60oC에서 45초간, ex- tension을 위하여 72oC에서 2분간의 과정을 35회 반 복하고, 최종 extension으로 72oC에서 5분간 반응시켰 다. 중합효소반응에 의해 얻어진 증폭 산물은 ethi- dium bromide가 포함된 1.5% agarose gel에 전기영동 을 시킨 후 Gel-Doc 2000 (Bio-Rad Laboratories, Her- cules, CA)으로 영상을 얻고, Quantity-One (Bio-Rad Laboratories, Hercules, CA) 프로그램을 이용하여, 증폭 산물을 농도계측법으로 정량하였다. 이 값을 β-actin과 의 비율로 환산하여 각 세포군 간의 mRNA 발현정 도를 비교하였다.
4. MCP-1과 RANTES의 단백질 발현에 대한 효소 면역 검사법(Enzyme Linked Immunosorbent assay; ELISA)
MCP-1과 RANTES의 단백질 발현은 24시간 배양 후에 얻어진 배양 상층액의 농도를 ELISA kit (Amersham Biosciences, Buckinghamshire, UK)를 사용 Fig. 1. Characterization of human mesangial cells was performed by immunohistochemistry. Cultured cells were
not stained with anti-cytokeratin (A), anti-CD31 (B), but positively stained with anti-vimentin (C), anti-smooth muscle actin (D).
A B
C D
하여 측정하였다. Anti-human MCP-1, anti-human RANTES가 도포된 96 well plate에 100μL의 bio- tinylated antibody reagent와 50μL의 STANDARD 용 액과 검체 배양액을 한 쌍씩 well에 첨가하고 실온 에서 2시간 동안 반응시켰다. 다시 100μL의 strept- avidin-HRP conjugate reagent를 처리한 후 실온에서 30분 동안 반응시켰다. 100μL의 TMB substrate로 발 색반응을 유도하였고, 30분 후에 stop solution으로 중 지시켰다. 자동화된 마이크로 평판 판독기 (VERSA- maxⓇ; Molecular Devices, Palo Alto, CA)를 이용하여 파장이 450 nm, 550 nm일 때의 흡광도를 읽고, 파장 의 보정을 위하여 450 nm의 값에서 550 nm의 값을 감한 값을 취하였다.
5. ICAM-1과 CD40 발현에 대한 유세포분 (flow cytometry)
ICAM-1과 CD40의 세포막 발현은 anti-CD54 (Phar- Mingen, San Diego, CA), anti-CD40 (R&D Systems, Minneapolis, MN) 단클론 항체를 이용한 유세포분석 법을 이용하여 측정하였다. 혈관간세포를 6 well plate에 1×105개씩 분주하고, 각각의 사이토카인을 처리한 후 ICAM-1의 발현을 보기 위해 24시간, CD40는 48시간 동안 20% 우혈청 배양액에서 배양 하였다. Trypsin-EDTA로 세포를 6 well plate에서 분 리하여 phosphate buffer saline에 재부유시킨 다음 anti-CD54, anti-CD40 단클론 항체를 첨가하고 4oC에
Fig. 2. MCP-1 mRNA expressions in human mesangial cells with various cytokine stimulations assayed by RT- PCR. (A) Representative results of gel electrophoresis; lane 1: media alone, lane 2: IFN-γ, lane 3: IL-10, lane 4: IL-4, lane 5: IFN-γ+IL-10, lane 6: IFN-γ+IL-4, lane 7: IFN-γ+IL-10+IL-4 (B) Den- sitometric analysis of the PCR product expressed as the ratio of MCP-1:β-actin. The mean±SEM of three experiments are presented. *p<0.05 versus media alone, **p<0.05 versus IFN-γ treated cells.
`
1 2 3 4 5 6 7
IFN-γ (250 U/mL) - + - - + + +
IL-10 (10 ng/mL) - - + - + - +
IL-4 (10 mg/mL) - - - + - + +
0 1 2 3 4 5 6 7
A
β-actin MCP-1
2
1 3 4 5 6 7
*
**
Ratio of MCP-1/-actinβ
B
* *
서 30분간 반응시켰으며 PBS로 3회 세척 후 fluo- rescein isothiocyanate (FITC)-conjugated goat anti- mouse IgG (Biosource International, Camarillo, CA)로 다시 4oC에서 30분간 반응하였다. 대조군으로 anti- CD54, anti-CD40 단클론 항체 없이 FITC-conjugated goat anti-mouse IgG만으로 반응시킨 세포를 사용하 였다. 염색된 세포는 즉시 검사하거나 1% para- formaldehyde로 고정시킨 후 FACScaliburⓇ (Bekton Dickinson, San Jose, CA)를 사용하여 세포막 발현을 측정하였다. 자료 분석은 CellQuest version 3.1 (Becton Dickinson, San Jose, CA)을 이용하였다.
6. 통계 분석
모든 수치는 평균±평균의 표준오차(mean±stan- dard error of mean, SEM)로 표시하였다. 각 세포군 간의 차이를 비교하기 위하여 윈도용 통계프로그램 인 SPSS 9.0의 paired Student t-test를 사용하였으며, 통계적 유의수준은 p<0.05로 하였다.
결 과
1. 혈관간세포의 MCP-1 mRNA의 발현
IFN-γ는 혈관간세포의 MCP-1 mRNA 발현을 대 조군에 비해 3.7배 증가시켰다. IL-10과 IL-4에 의한 발현의 증가는 미약하였다. IL-10을 IFN-γ와 함께 처리하였을 때 IFN-γ를 단독으로 처리한 세포군보 다 mRNA의 1.6배 증가함을 관찰할 수 있었다. IL-4 는 IFN-γ에 의해 유도된 mRNA의 양과 IL-10과 IFN-γ를 함께 처리한 세포군의 mRNA의 양에 큰 영향을 주지 않았다(그림 2).
2. 사이토카인 자극에 의한 혈관간세포의 MCP-1 분비능
세포 배양액에 IFN-γ를 첨가하면, MCP-1의 발현 이 대조군에 비해 4.8배 증가하였으며, IL-10과 IL-4 의 자극에 의해서도 MCP-1의 발현이 각각 1.2배, 2.6배 증가하였다. IL-10은 IFN-γ에 의해 증가된 MCP-1의 발현을 IFN-γ 단독으로 처리한 농도의 1.6배 증가시켰다. IL-4 또한 소량 증가시켰으나,
Fig. 3. MCP-1 protein expressions in human mesangial cells with various cytokine stimulations assayed by ELISA. The mean±SEM of three indepen- dent experiments are presented. *p<0.05 ver- sus media alone, **p<0.05 versus IFN-γ treated cells.
IFN-γ
- + - - + + +
(250 U/mL) IL-10
- - + - + - +
(10 ng/mL)
IL-4 - - - + - + +
(10 ng/mL)
Fig. 4. RANTES protein expressions in human mesan- gial cells with various cytokine stimulations assayed by ELISA. The data are shown as mean±SEM of three independent experiments.
*p<0.05 versus media alone.
IFN-γ
- + - - + + +
(250 U/mL) IL-10
- - + - + - +
(10 ng/mL)
IL-4 - - - + - + +
(10 ng/mL)
IFN-γ 단독투여와 의미 있는 차이를 보이지 않았다 (그림 3).
3. 혈관간세포의 RANTES의 단백질의 발현
IFN-γ에 의한 RANTES의 발현은 대조군보다 2.4 배 증가되었다. 그러나, IL-10 또는 IL-4는 대조군과 비교하여 의미 있는 차이를 보이지 않았고, IFN-γ에 의해 증가된 RANTES의 발현에 영향을 주지 않았다 (그림. 4).
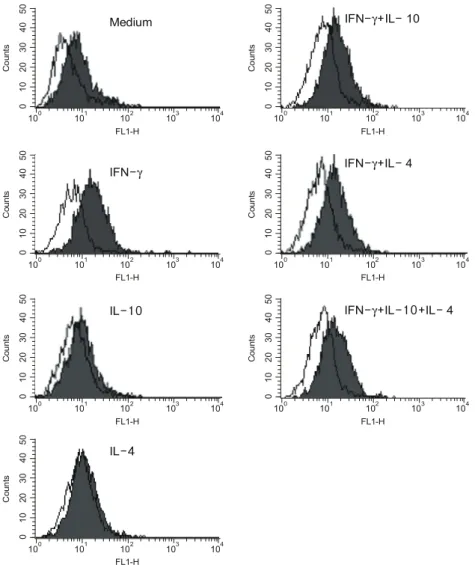
4. 혈관간세포의 ICAM-1의 발현 양상
사이토카인을 처리하지 않은 대조군에서 ICAM-1 의 발현은 관찰되지 않았다. ICAM-1은 IFN-γ, IL-10, IL-4 중 IFN-γ에 의해서만 발현이 유도되었 다(배양액 단독: 6.7±0.46, IFN-γ: 58.6±17.3, mean
±SEM of mean channel fluorescence). IFN-γ에 IL-10 또는 IL-4를 추가한 세포군의 ICAM-1의 발현은 IFN-γ를 단독으로 처리한 세포군과 비교하여 의미
Fig. 5. IFN-γ induced ICAM-1 expressions of human mesangial cells were not influenced by Th2 cytokines.
Cultured human mesangial cells were incubated with IFN-γ (250 U/mL), IL-10 (10 ng/mL), IL-4 (10 ng/mL), IFN-γ+IL-10, IFN-γ+IL-4, and IFN-γ+IL-10+IL-4 for 24 hrs and ICAM-1 expressions were analyzed by flow cytometry. Negative controls are shown as the open histogram. These histograms represent typical example of three independent experiments.
Counts
101
FL1-H
10203040500
100 102 103 104
Counts
101
FL1-H
10203040500
100 102 103 104
Counts
101
FL1-H
10203040500
100 102 103 104
Counts
101
FL1-H
10203040500
100 102 103 104
Counts
101
FL1-H
10203040500
100 102 103 104
Counts
101
FL1-H
10203040500
100 102 103 104
Counts
101
FL1-H
10203040500
100 102 103 104
Medium IFN- + -10γ IL
IFN-γ
IL-10
IL-4
IFN- + -γ IL 4
IFN- +γ IL 10 IL- + -4
있는 차이를 보이지 않았다(IFN-γ+IL-10: 53.9±4.2, IFN-γ+IL-4: 54.6±9.7). IL-10과 IL-4를 함께 처리한 세포군은 mean channel fluorescence가 70.4±20.9로서 IFN-γ에 비해 유의한 차이를 관찰할 수 없었다(그 림 5).
5. 혈관간세포의 CD40의 발현
그림 6에서와 같이 사이토카인을 처리하지 않은
대조군에서도 CD40의 기저 발현이 소량 관찰되었 다. 사이토카인 중 IFN-γ만이 CD40의 발현을 의미 있게 증가시켰다(배양액 단독: 7.4±2.4, IFN-γ: 16.6
±4.2). 그러나 IL-10 또는 IL-4는 IFN-γ에 의해 증 가된 CD40의 발현에 영향을 주지 않았다(IFN-γ+
IL-10: 16.4±2.2, IFN-γ+IL-4: 14.1±2.4, IFN-γ+
IL-4+IL-10: 14.9±2.9).
Fig. 6. IFN-γ induced CD40 expression by human mesangial cells was not influenced by Th2 cytokines. Cultured human mesangial cells were incubated with IFN-γ (250 U/mL), IL-10 (10 ng/mL), IL-4 (10 ng/mL), IFN- γ+IL-10, IFN-γ+IL-4, and IFN-γ+IL-10+IL-4 for 24 hrs and CD40 expressions were analyzed by flow cytometry. Negative controls are shown as the open histogram. These histograms represent typical example of three independent experiments.
Counts
101
FL1-H
10203040500
100 102 103 104
Counts
101
FL1-H
10203040500
100 102 103 104
Counts
101
FL1-H
10203040500
100 102 103 104
Counts
101
FL1-H
10203040500
100 102 103 104
Counts
101
FL1-H
10203040500
100 102 103 104
Counts
101
FL1-H
10203040500
100 102 103 104
Counts
101
FL1-H
10203040500
100 102 103 104
Mediu m
IFN- + -γ IL
IFN-
IL-
IL-
IFN- + -γ IL
IFN- +γ IL10 IL + -
Medium IFN- + -γ IL 10
IFN-γ
IL-10
IL-4
IFN- + -γ IL 4
IFN- + -10+ -γ IL IL 4
고 찰
T세포가 사이토카인의 분비 양상과 기능에 따라 Th1과 Th2로 구분이 되면서 오랫동안 루푸스는 Th2 면역반응이 지배적인 대표적인 자가면역질환으로 인 정되어 왔다. 루푸스 환자의 혈액에서 IL-10이 비정 상적으로 증가되어 있고, IL-10의 혈 중 농도는 질병 의 활성도와 상관관계를 보이듯이, IL-10은 루푸스의 질병활성도의 중요한 조절자로 인식되어 왔다24-25). 또한 IL-10에 대한 항체는 NZB/NZW mice의 질병의 발생을 지연시키고, 항 IL-10 항체가 루푸스를 호전 시킨다는 보고는 루푸스의 병인에서 IL-10의 중요성 을 시사한다12,26). 반면에 루푸스에서 Th1 면역반응의 중요성을 강조한 연구 결과들도 많다. NZB/NZW F1 mice에 IFN-γ를 투여하면 질환의 진행이 가속화되 고, IFN-γ에 대한 항체를 투여했을 때 루푸스 발병 이 지연된다는 보고를 시작으로 NZB/NZW F1 mice 에 IFN-γ에 대한 가용성 수용체를 투여하고, MRL- Faslpr mice에서 IFN-γ 또는 IFN-γ 수용체의 유전자 를 제거했을 때 관찰되는 자가면역성 신염의 발생률 과 사망률의 현저한 감소에 대한 일련의 동물실험의 결과들은 루푸스신염의 병인에서 IFN-γ의 중추적인 역할을 입증하였다5-8). 또한 class IV 루푸스신염 환 자의 말초혈액의 단핵구와 신조직에서도 정상인과 class V 루푸스신염 환자에 비해 IFN-γ의 발현의 증 가를 관찰한 최근의 보고는 특히 증식성 루푸스 신 염에서 IFN-γ에 의한 면역반응이 지배적임을 뒷받 침한다10-12).
이러한 상반된 보고들은 루푸스는 Th1, Th2 중 어 느 한쪽으로 치우치지 않고 두 개의 면역반응이 공 존하는 질환이라는 것을 시사한다. Th2 사이토카인 이 B 세포의 활성화와 자가항체의 생성을 촉진하는 전신적인 효과를 통해 루푸스신염의 진행에 기여하 는 것과 더불어, 사구체내의 국소적인 염증반응의 조절에도 직접적으로 관여하는지는 밝혀져 있지 않 다. 저자들은 Th1, Th2 사이토카인이 함께 증가되어 있는 루푸스 신염의 사이토카인 환경을 가정하고 혈 관간세포를 대상으로 한 체외 실험을 통해 Th1, Th2 사이토카인 간의 상호작용을 관찰하고자 하였다. 이 를 위해 IFN-γ에 의해 유도된 혈관간세포의 che-
mokine, 유착분자와 CD40의 발현에 대한 IL-10, IL-4 의 영향을 관찰하고자 하였고 이는 증식성 루푸스 신염의 신손상의 과정에서 Th2 사이토카인이 차지 하는 사구체내의 국소적인 역할에 대한 이해를 높이 는데 도움이 될 수 있을 것이라고 생각하였다.
MCP-1은 단핵구세포와 T 세포를 유인하는 chemo- kine으로 MRL-FASlpr mice와 루푸스 신염의 사구체 와 세뇨관에서 발현이 증가되어 있다27,28). MCP-1의 유전자를 제거하면 사구체내로의 염증 세포 침윤의 감소와 함께 신장조직의 염증 소견과 사망률이 감소 하는 결과로 보아 MCP-1은 루푸스 신염에서 신장내 로의 세포 침윤을 일으켜 사구체신염을 진행시키는 가장 중요한 물질의 하나로 인식되고 있다29). 특히 소변 내의 MCP-1의 수치가 신조직 내로의 백혈구 침윤 정도와 밀접한 관계가 있고, MRL-Faslpr mice의 신조직에서 검출되는 MCP-1의 대부분이 침윤 세포 보다 신장 실질세포에서 발견된다는 사실은 혈관간 세포와 같은 신장 실질세포가 MCP-1을 생성하는 주 된 장소라는 것을 의미한다30,31). 혈관간세포는 IL-1, TNF-α, IFN-γ, 면역복합체에 반응하여 MCP-1을 분 비하는 것으로 널리 알려져 있지만, 아직까지 혈관 간세포의 MCP-1의 생성에 대한 IFN-γ와 Th2 사이 토카인 간의 상호작용은 보고된 적이 없었다16,32). 인 류의 혈관간세포를 대상으로 시행한 본 연구에서는 IL-10이 IFN-γ에 의해 유도된 MCP-1의 발현을 억제 하기보다 오히려 더욱 증가시킴을 확인하였다. 그러 나 본 체외 실험에서 사용한 IL-10의 용량이 생리적 으로 적절한 용량인지의 여부는 명확하지 않으며, 향후 사이토카인의 용량에 따른 효과를 확인해 볼 필요가 있다고 사료된다.
RANTES도 MCP-1과 함께 단핵세포, T세포를 신 장 내로 유인하는 chemokine으로, MRL-Faslpr mice의 신조직에서 발현이 증가되어 있어 루푸스에 의한 신 손상에 관여할 것으로 생각되었다33). 그러나 MRL- Faslpr mice에서 RANTES 유전자의 제거가 신손상에 영향을 주지 않는다는 최근의 보고가 있어 루푸스신 염에 있어서 RANTES의 역할에 대한 규명을 위해서 는 더 많은 연구가 필요하리라 생각된다34).
ICAM-1은 혈관내막세포의 β2-integrin과의 결합을 통해 백혈구가 염증 부위로 이동하는데 필수적인 유 착분자로 루푸스신염과 MRL-Faslpr mice의 혈관간세
포와 세뇨관상피세포에서 발현이 증가되어 있다35,36). ICAM-1에 대한 항체를 투여하거나, 유전자를 제거 했을 때 혈관간세포와 대식세포 간의 결합이 현저히 감소되고, 사구체신염의 염증소견이 개선된다는 보 고는 ICAM-1은 백혈구의 이동뿐만 아니라, 단핵구 세포와 신장 실질세포 간의 접촉에도 관여하는 것으 로 생각되고 있다35,36). 단핵구세포에서는 IFN-γ에 의한 ICAM-1의 발현을 IL-10이 억제하는 반면, rat의 혈관간세포에서는 IL-10이 ICAM-1의 발현을 유도하 고, IFN-γ에 의한 ICAM-1의 발현을 더욱 증가시킨 다는 보고가 있다23,37). 그러나 rat의 결과와 다르게, 본 실험에서는 IL-10이 혈관간세포의 ICAM-1 발현 을 증가시키지 못하였을 뿐 아니라 IFN-γ에 의한 ICAM-1 발현에 미치는 영향을 관찰할 수 없었다.
이는 아마 인간 세포와 rat 세포와의 종의 차이에서 기인한 것으로 추정된다.
CD40-CD40 리간드는 T 세포와 B 세포 간의 공동 자극 신호로서의 기능뿐만 아니라 CD40를 발현하는 수상돌기세포, 단핵구, 혈관간세포, 내피세포, 섬유아 세포, 상피세포와 T 세포 간의 직접적인 접촉을 매 개함으로써 MCP-1, E-selectin, ICAM-1, VCAM-1의 발현을 증가시켜 염증 부위로 백혈구의 이동을 촉진
시킨다38-40). Class III와 class IV 루푸스신염의 사구
체 조직에서 CD40의 발현이 class V 신염에 비해 현 저히 증가되어 있고, IFN-γ에 의한 혈관간세포의 CD40 발현의 유도는 증식성 루푸스신염에서 CD40- CD40 리간드를 통한 T 세포와 혈관간세포의 세포간 상호작용의 중요성을 보여준다41). 대식세포와 micro- glia세포에서는 IL-4가 IFN-γ에 의한 CD40 발현을 억제한다는 보고가 있으나, 본 연구에서는 IL-10과 IL-4는 IFN-γ에 의한 CD40 발현에 영향을 주지 않 았다42).
혈관간세포는 사구체의 모세혈관 고리를 지지해주 고, 수축 작용을 제공하는 구조적인 기능뿐만 아니 라, 사구체내의 면역반응을 조절하는 신장실질세포 로서 염증 자극에 대하여 MHC class II 분자, Fc 수 용체, nitric oxide (NO) 등을 생성하여 대식세포와 유사한 기능을 수행한다43). 혈관간세포의 활성화와 증식은 루푸스 신염 외에도 IgA 신염, 막증식성 사 구체신염 등의 사구체 염증을 일으키는 다양한 질환 에서 관찰된다. 본 연구의 결과 IL-10은 단핵구와 대
식세포에서 관찰되었던 일반적인 항염증 기능과는 달 리, 인간 혈관간세포에서는 IFN-γ에 의한 RANTES, ICAM-1, CD40의 발현에는 영향을 주지 않고, MCP-1 의 발현을 더욱 증가시켰다. 대상 세포에 따른 IL-10 의 상이한 효과는 IL-10이 대식세포에서는 IFN-γ에 의한 MHC class II 분자의 발현을 감소시키는 반면, 혈관간세포에서는 더욱 증가시킴을 관찰한 이전의 보고에서도 찾아볼 수 있다44). 이러한 결과는 IL-10 이 IFN-γ와 함께 적어도 혈관간세포가 참여하는 사 구체내의 염증반응을 촉진하는 방향으로 작용할 것 이라는 추정을 가능케 한다. 본 연구에서는 혈관간 세포의 MHC class II의 발현을 동시에 관찰하지 못 하였고, 루푸스 신염 환자의 세포로 실험하지 못하 는 기술적인 한계가 있으나 IL-10이 사구체내에서 국소 염증의 촉진에 기여할 가능성을 제시하였다.
Th1과 Th2 면역반응이 복합적으로 작용하는 루푸 스와 달리 Th1 면역반응에 전적으로 의존하는 반월 체 사구체신염과 항 사구체기저막 신염의 동물모델 에서도, IL-10의 역할은 논쟁의 여지가 많다. 항 사 구체기저막 신염의 동물모델에서 IL-10의 투여가 사 구체내의 대식세포의 침윤을 증가시킨다는 결과가 있는가 하면, 반월체 사구체신염과 항 사구체기저막 신염의 유사한 동물 모델에서 IL-10과 IL-4의 투여가 신염의 발생을 예방하거나, 호전시킨다는 정반대의 결과를 보고하였다45-47). 이러한 IL-10에 의한 서로 다른 효과를 설명할 구체적인 근거는 제시되지 않았 지만, 작용하는 세포, 질환의 진행 단계와 사이토카 인 환경 등의 여러 가지 요인들에 의해 영향을 받는 것으로 추정된다44).
MCP-1에 대한 IL-10과 IFN-γ 간의 상승작용에 대 한 기전은 아직까지 자세히 밝혀진 바가 없다. 단핵 구의 IFN-γ 유도 유전자의 발현에 대한 IL-10의 억 제효과의 기전으로 supressor of cytokine signaling-3 (SOCS-)3의 발현에 의한 signal transducer and activa- tors of transcription-1 (STAT-1)의 인산화의 억제가 가능한 기전으로 제시되고 있으나48), 혈관간세포에 서 두 사이토카인 간의 상호작용의 기전을 밝히기 위해서는 STAT-SOCS뿐만 아니라 이와 무관한 다양 한 세포내 사이토카인 신호전달 물질의 발현에 대한 연구가 필요할 것으로 생각된다.
결 론
본 연구는 루푸스의 동물모델이나, 루푸스 환자에 서 추출한 혈관간세포가 아닌, 정상적인 인간혈관간 세포를 대상으로 하였고, 다른 세포와 사이토카인의 영향을 배제한 in vitro 실험이기 때문에 본 연구의 결과를 루푸스 신염에 직접적으로 적용할 수는 없으 나, Th1, Th2 사이토카인이 공존하는 증식성 루푸스 신염에서 IL-10이 IFN-γ에 의해 유도된 사구체내 염 증 현상을 MCP-1의 발현 증가를 통해 오히려 촉진 할 가능성을 제시하고 있다고 생각된다.
REFERENCES
1) Falcone M, Sarvetnick N. Cytokines that regulate autoimmune responses. Curr Opin Immunol 1999;
11:670-6.
2) Mosmann TR, Cherwinski H, Bond MW, Giedlin MA, Coffman RL. Two types of murine helper T cell clones. I. Definition according to profiles of lymphokine activities and secreted proteins. J Immunol 1986;136:2348-57.
3) Goldman M, Druet P, Gleichmann E. TH2 cells in systemic autoimmunity: insights from allogeneic diseases and chemically induced autoimmunity.
Immunol Today 1991;12:223-7.
4. Klinman DM, Steinberg AD. Inquiry into murine and human lupus. Immunol Rev 1995;144:157-93.
5. Jacob CO, van der Meide PH, McDevitt HO. In vivo treatment of (NZB×NZW) F1 lupus-like nephritis with monoclonal antibody to interferon-γ.
J Exp Med 1987;166:798-803.
6) Ozmen L, Roman D, Fountoulakis M, Schmid G, Ryffel B, Garotta G. Experimental therapy of systemic lupus erythematosus: the treatment of NZB/W mice with mouse soluble interferon-γ receptor inhibits the onset of glomerulonephritis.
Eur J Immunol 1995;25:6-12.
7) Haas C, Ryffel B, Hir ML. IFN-γ is essential for the development of autoimmune glomerulonephritis in MRL/lpr mice. J Immunol 1997;158:5484-91.
8) Haas C, Ryffel B, Hir ML. IFN-γ receptor deletion prevents autoantibody production and glo- merulonephritis in lupus-prone (NZB×NZW) F1 mice. J Immunol 1998;160:3713-8.
9) Akahoshi M, Nakashima H, Tanaka Y, Kohsaka T,
Nagano S, Ohgami E. Th1/Th2 balance of periph- eral T helper cells in systemic lupus erythema- tosus. Arthritis Rheum 1999;42:1644-8.
10) Masutani K, Akahoshi M, Tsuruya K, Tokumoto M, Ninomiya T, Hirakata H, et al. Predominance of Th1 immune response in diffuse proliferative lupus nephritis. Arthritis Rheum 2001;44: 2097-106.
11) Uhm WS, Na K, Song GW, Jung SS, Lee T, Park MH, et al. Cytokine balance in kidney tissue from lupus nephritis patients. Rheumatology 2003;
42:935-8.
12) Ishida H, Muchamuel T, Sakaguchi S, Andrade S, Menon S, Howard M. Continuous administration of anti-interleukin 10 antibodies delays onset of autoimmunity in NZB/W mice. J Exp Med 1994;
179:305-10.
13) Peng SL, Moslehi J, Craft J. Roles of interferon-γ and interleukin-4 in murine lupus. J Clin Invest 1997;99:1936-46.
14) O’Shea JJ, Ma A, Lipsky P. Cyokine and auto- immunity. Nature Rev Immunol 2002;2:37-45.
15) Kuroiwa T, Lee EG. Cellular interactions in the pathogenesis of lupus nephritis: the role of T cells and macrophages in the amplification of the inflammatory process in the kidney. Lupus 1998;
7:597-603.
16) Rovin BH, Yoshiumura T, Tan L. Cytokine-in- duced production of monocyte chemoattractant protein-1 by cultured human mesangial cells. J Immunol 1992;148:2148-53.
17) McMurray RW. Adhesion molecules in autoim- mune disease. Semin Arthritis Rheum 1996;25:
215-33.
18) Wuthrich RP, Jevnikar AM, Takei F, Glimcher LH, Kelley VE. Intercellular adhesion molecule-1 (ICAM-1) expression is upregulated in auto- immune murine lupus nephritis. Am J Pathol 1990;136:441-50.
19) Brennan DC, Jevnikar AM, Takei F, Reubin-Kelley VE. Mesangial cell accessory functions; mediation by ICAM-1. Kidney Int 1990;38:1039-46.
20) van Kooten C, Gerritsma JS, Paape ME, van Es LA, Banchereau J, Daha MR. Possible role for CD40-CD40L in the regulation of interstitial infiltration in the kidney. Kidney Int 1997;51:
711-21.
21) De Wall MR, Abrams J, Bennett B, Figdor C, de Vries J. IL-10 inhibits cytokine synthesis by human monocytes: an autoregulatory role of IL-10
produced by monocytes. J Exp Med 1991;174:
1209-20.
22) Larner AC, Petricoin EF, Nakagawa Y, Finbloom DS. IL-4 attenuates the transcriptional activation of both IFN-α and IFN-γ induced cellular gene expression in monocytes and monocytic cell lines.
J Immunol 1993;150:1944-50.
23) Song S, Ling-Hu H, Roebuck KA, Rabbi MF, Donnelly RP, Finnegan A. Interleukin-10 inhibits interferon-γ-induced intercellular adhesion molecule-1 gene transcription in human monocytes. Blood 1997;89:4461-9.
24) Llorente L, Richaud-Patin Y, Wijdenes J, Alcocer- Varela J, Maillot MC, Durand-Gasselin I, et al.
Spontaneous production of interleukin-10 by B lymphocytes and monocytes in systemic lupus erythematosus. Eur Cytokine Netw 1993;4:421-7.
25) Park YB, Lee SK, Kim DS, Lee J, Lee CH, Song CH. Elevated interleukin-10 levels correlated with disease activity in systemic lupus erythematosus.
Clin Exp Rheumatol 1998;16:283-8.
26) Llorente L, Richaud-Patin Y, Garcia-Padilla C, Claret E, Jakez-Ocampo J, Cardiel MH, et al.
Clinical and biologic effects of anti-interleukin-10 monoclonal antibody administration in systemic lupus erythematosus. Arthritis Rheum 2000;43:
1790-800.
27) Zoja C, Liu XH, Donadelli R, Abbate M, Testa D, Remuzzi G, et al. Renal expression of mon- ocyte chemoattractant protein-1 in lupus auto- immune mice. J Am Soc Nephrol 1997;8:720-9.
28) Prodjosudjadi W, Gerritsma JS, van Es LA, Daha MR, Bruijn JA. Monocyte chemoattractant pro- tein-1 in normal and diseased human kidneys: an immunohistochemical analysis. Clin Nephrol 1995;
44:148-55.
29) Tesch GH, Maifert S, Schwarting A, Rollins BJ, Kelley VR. Monocyte chemoattractant protein 1-dependent leukocytic infiltrates are responsible for autoimmune disease in MRL-Faslpr mice. J Exp Med 1999;190:1813-24.
30) Noris M, Bernasconi S, Casiraghi F, Sozzani S, Gotti E, Mantovani A, et al. Monocyte chemo- attractant protein-1 is excreted in excessive amounts in the urine of patients with lupus ne- phritis. Lab Invest 1995;73:804-9.
31) Wada T, Yokoyama H, Su SB, Mukaida N, Iwano M, Matsushima K, et al. Monitoring urinary levels of monocyte chemotactic and activating factor
reflects disease activity of lupus nephritis. Kidney Int 1996;49:761-7.
32) Schwarz M, Radeke HH, Resch K, Uciechowski P.
Lymphocyte-derived cytokines induce sequential expression of monocyte- and T cell-specific che- mokines in human mesangial cells. Kidney Int 1997;52:1521-31.
33) Moore, KJ, Wada, T, Barbee, SD, Kelley VR.
Gene transfer of RANTES elicits autoimmune renal injury in MRL-Faslpr mice. Kidney Int 1998;53:1631-41.
34) Tsukahara T, Makino Y, Fujii T, Ogawa M, Saisho H, Danoff TM, et al. Role of RANTES in the development of autoimmune tissue injuries in MRL-Fas lpr mice. Clin Immunol 2002;103:89-97.
35) Wuthrich RP, Jevnikar AM, Takei F, Glimcher LH, Kelley VE. Intercellular adhesion molecule-1 (ICAM-1) expression is upregulated in autoim- mune murine lupus nephritis. Am J Pathol 1990;
136:441-50.
36) Bruijn JA, Bergijk EC, de Heer E, Kootstra CJ.
Molecular adhesion mechanisms and recruitment of inflammatory cells in renal diseases. Life Sci Adv 1994;13:9-15.
37) Chadban SJ, Tesch GH, Foti R, Lan HY, Atkins RC, Nikolic-Paterson DJ. Interleukin-10 differen- tially modulate MHC class II expression by mesangial cells and macrophage in vitro and in vivo. Immunology 1998;94:72-8.
38) Alderson MR, Armitage RJ, Tough TW, Strock- bine L, Fanslow WC, Spriggs MK. CD40 expres- sion by human monocytes: regulation by cytokines and activation of monocytes by the ligand for CD40. J Exp Med 1993;178:669-74.
39) Caux C, Massacrier C, Vanbervliet B, Dubois B, Van Kooten C, Banchereau J, et al. Activation of human dendritic cells through CD40 cross-linking.
J Exp Med 1994;180:1263-72.
40) Karmann K, Hughes CC, Schechner J, Fanslow WC, Pober JS. CD40 on human endothelial cells:
inducibility by cytokines and functional regulation of adhesion molecule expression. Proc Natl Acad Sci USA 1995;92:4342-6.
41) Yellin MJ, D'Agati V, Parkinson G, Han AS, Szema A, Baun D, et al. Immunohistologic anal- ysis of renal CD40 and CD40L expression in lupus nephritis and other glomerulonephritides.
Arthritis Rheum 1997;40:124-34.
42) Nguyen VT, Benveniste EN. IL-4-activated STAT-6
inhibits IFN-γ induced CD40 gene expression in macrophages/microglia. J Immunol 2000;165:6235-43.
43) Tetsuka T, Daphna-Iken D, Srivastava SK, Baier LD, DuMaine J, Morrison AR. Cross-talk between cyclooxygenase and nitric oxide pathways: pros- taglandin E2 negatively modulates induction of nitric oxide synthase by interleukin 1. Proc Natl Acad Sci USA 1994;91:12168-72.
44) Chadban SJ, Tesch GH, Foti R, Lan HY, Atkins RC, Nikolic-Paterson DJ. Interleukin-10 differen- tially modulates MHC class II expression by mesangial cells and macrophages in vitro and in vivo. Immunology 1998;94:72-8.
45) Chadban SJ, Tesch GH, Lan HY, Atkins RC, Nikolic-Paterson DJ. Effect of interleukin-10 treat- ment on crescentic glomerulo-nephritis in rats.
Kidney Int 1997;51:1809-17.
46) Kitching AR, Tipping PG, Huang XR. Interleukin-4 and interleukin-10 attenuate established crescentic glomerulonephritis in mice. Kidney Int 1997;52:
52-9.
47) Huang XR, Kitching AR, Tipping PG. Interleu- kin-10 inhibits macrophage induced glomerular injury: studies in passive anti-glomerular basement membrane glomerulonephritis. J Am Soc Nephrol 2000;11:262-9.
48) Ito S, Ansari P, Sakatsume M, Dickensheets H, Vazquez N, Donnelly RP, et al. Interleukin-10 inhibits expression of both interferon-α and inter- feron-γ induced genes by suppressing tyrosine phosphorylation of STAT1. Blood 1999;93:1456-63.