Expression Properties and Skin Permeability of Human Basic Fibroblast Growth Factor with or without PTD Fused to N- or C-terminus in Escherichia coli
In-Sun Park
1, Chung-Hyeon Choe
2, Bo-Ra Kwon
1, Young-Ji Choi
1, Tae-Ho Kwon
2, Kang-Yeol Yu
1, Juhyung Lee
3,4and Young-Moo Choo
1*
1Jeonju AgroBio-Materials Institute, Jeonju 54810, Korea
2Natural Bio-Materials Inc., Wanju 55322, Korea
3Department of Preventive Medicine, Chonbuk National University Medical School, Jeonju 54907, Korea
4Research Institute of Clinical Medicine of Chonbuk National University and Chonbuk National University Hospital, Jeonju 54907, Korea Received November 17, 2017 /Revised March 20, 2018 /Accepted March 22, 2018
Human fibroblast growth factor (FGF) has the potential to be a commercially important therapeutic or cosmeceutical agent due to its ability to generate tissue and heal wounds. Granting permeability into skin tissues increases the therapeutic effects of FGF. Thus, several researchers have attempted the fusion of FGF conjugates with protein transduction domains (PTDs) to investigate the transduction ability and therapeutic effects of FGF. Less is known, however, about whether the location of PTD fused to the N- or C-terminus of FGF proteins has a significant impact on the folding and stability in Escherichia coli, and eventually, on transduction. Here, we report cloning of human basic fibroblast growth factor (FGF2) as a control and FGF2 with PTD fused to the N- or C-terminal ends of FGF proteins by an overlap extension PCR. We performed expression, verified expression properties of re- combinant FGF2 without or with PTD fused to the N-terminus and the C-terminus, and investigated transduction ability into tissue by treating the dorsal skin of mice subjects. As a result, FGF2 and FGF2-PTD (fused to C-terminus) fusion protein were expressed as soluble forms suitable for straight- forward purification, unlike insoluble PTD-FGF2 (fused to N-terminus), but only FGF2-PTD fusion protein could transduce into the dorsal skin tissue of the mice subjects. Our results suggest that FGF2 with PTD fused to the C-terminus is more efficient than other options in terms of expression, purifica- tion, and delivery into skin tissue, as it does not require labor-intensive, costly, and time-consuming methods.
Key words : FGF2 conjugates, purification, solubility, transduction
*Corresponding author
*Tel : +82-63-711-1026, Fax : +82-63-711-1004
*E-mail : [email protected]
This is an Open-Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/3.0) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
Journal of Life Science 2018 Vol. 28. No. 3. 275~283 DOI : https://doi.org/10.5352/JLS.2018.28.3.275
Introduction
A few fibroblast growth factors (FGFs), such as FGF1, FGF2, FGF7, FGF10, FGF12, and FGF20, have radioprotective effects against radiation-induced injury, and are therefore potentially therapeutic in clinical applications [1, 15, 26-30].
What is more, these proteins may have effective cosmeceut- ical applications. Although these proteins may prove to be excellent agents for the prevention and treatment of skin damage for cosmeceutical purposes, one obstacle to over- come is efficient delivery of these proteins into tissues and
cells for beneficial clinical effects. A few studies have tried to use protein transduction domains (PTDs) to facilitate effi- cient delivery of FGF proteins into tissues and cells. Protein transduction domains (PTDs) may play an important role in intracellular delivery of PTD conjugates such as proteins, peptides, and antisense in in vivo animal models and in vitro various cells [22, 23, 36]. A recent study reported that the fusion of PTD-FGF1 and protein transduction domain-epi- dermal growth factor (PTD-EGF) showed efficient trans- duction of these proteins into tissues and had ultraviolet (UV) radiation-protective effects in UV-induced skin damage in mice models. Consequently, these findings suggested that the conjugated proteins might be useful as cosmeceutical agents for skin damage and UV-related injury [5].
A limitation is that it is difficult to obtain sufficient ther-
apeutic proteins for clinical or industrial applications due
to low availability in natural sources [24]. For example, while
FGF2 has the potential to become a commercially important
therapeutic agent due to its tissue generation and wound
healing effects [7], purification of native FGF2 from animal tissues is very difficult and expensive due to limited supply [14]. To cope with this situation, recombinant FGF2 has been directly expressed by using Escherichia coli as host cells [12, 14, 16, 19, 41]. Expression of desired recombinant proteins in E. coli is one useful aspect of biotechnology in satisfying the matter at hand. The practice is well established, and has become the most common molecular tool for protein expression. The expression and purification of large amounts of proteins is advantageous insofar as it allows researchers to study biochemical characterization, to use in industrial processes, and to develop commercial products [34], includ- ing therapeutic and cosmetic agents. One drawback, how- ever, is that high-level expression proteins in E. coli occasion- ally result in the formation of inclusion bodies [6, 8, 40], which are known as insoluble and inactive forms.
Although some research has reported that FGF2 is well expressed as soluble proteins with no inclusion body for- mation [4, 18, 39], many studies have performed FGF2 ex- pression and have attempted various refolding processes from inclusion bodies in bacterial cell lysates [2-4, 17]. The refolding process does not readily yield properly folded pro- teins from denatured proteins due to the disadvantages of aggregation and/or the misfolding of target proteins, which frequently occur in the purification process [25]. To over- come this problem, many tagging systems with FGF2 have been developed, such as b‘a’ domains of human protein di- sulfide isomerase (hPDIb‘a’) [25], hexahistidine [19], maltose binding protein (MBP) [21], glutathione S-transferase (GST) [37], thioredoxin (Trx) [14], and HaloTag [39]. All of these have improved expression and solubility problems in E. coli.
Although tagging systems have alleviated defects in ex- pression, solubility, and purification in E. coli, it is essential to take into account a number of criteria for optimal ex- pression conditions in this line of products. Moreover, it has not been reported whether the location of PTD fused to the N- or C-terminus of cargo proteins had a profound effect on folding and stability in E. coli. Herein, we performed ex- pression and purification of FGF2 without or with PTD fused to the N- and C-terminus, and determined expression properties in E. coli. Accordingly, using the recombinant pro- teins, we show transduction ability in the skin of mice subjects.
Materials and Methods
Gene synthesis and cloning of FGF2 or PTD-FGF2 Sequences of human basic fibroblast growth factor (FGF2) and protein transduction domains (PTDs) (intracellular de- livery peptides Pep-1 and Antp) were obtained from Gen Bank (accession numbers NM_002006.4, HE605043.1, and S63455.1). Mature FGF2 (495-935 base pair (bp)) and PTD genes were provided by a custom gene synthesis service from Bioneer (Daejeon, South Korea). To obtain mature FGF2 with PTD fused to the N- or C-terminus, overlap extension polymerase chain reaction (OE-PCR) was performed with the following gene specific primers (the underlined section shows the restriction enzyme site):
FGF2-Fw (Nde I): 5’- GGAATTCCATATGCCCGCCTTGC CCGAGGAT -3’; FGF2-Rv (Xba I): 5’- GCTCTAGAGCTCA GCTCTTAGCAGACATTGG -3’; Pep-1-Fw (Nde I): 5’- GGAATTCCATATGAA AGA AACC TGGTGGGA A -3’;
Pep-1-Rv (Xba I): 5’- GCTCTAGAGCTCACACTTTACGTTT TTTTTT -3’; Pep-1-FGF2-Fw: 5’- AAAAAA AAACGTAAAG TGCCCGCCTTGCCCGAGGAT -3’; Pep-1-FGF2-Rv: 5’- ATC CTCGGGCAAGGCGGGCACTTTACGTTTTTTTTT -3’; FGF2- Pep-1-Fw: 5’- CCAATGTCTGCTAAGAGCAAAGAAACCT GGTGGGAA -3’; FGF2-Pep-1-Rv: 5’- TTCCCACCAGGTTTC TTTGCTCTTAGCAGACATTGG -3’; Antp-Fw (Nde I): 5’- GGAATTCCATATGCGCCAGATTAAGATCTGG -3’; Antp- Rv (Xba I): 5’- GCTCTAGAGCTCACTTCTTCCACTTCATG CG -3’; Antp-FGF2-Fw: 5’- CGCATGAAGTGGAAGAAGCC CGCCTTGCCCGAGGAT -3’; Antp-FGF2-Rv: 5’- ATCCTCG GGCAAGGCGGGCTTCTTCCACTTCATGCG -3’; FGF2-Antp- Fw: 5’- CCAATGTCTGCTAAGAGCCGCCAGATTAAGAT CTGG -3’; FGF2-Antp-Rv: 5’- CCAGATCTTAATCTGGCGG CTCTTAGCAGACATTGG -3’.
Polymerase chain reaction (PCR) reactions were carried out using the Advantage GC 2 PCR Kit (Clontech, Mountain View, CA, USA). Polymerase chain reaction (PCR) products were purified with a QIAquick Gel Extraction Kit (Qiagen, Valencia, CA, USA), and subsequently cloned with pGEM-T Vector Systems (Promega, Madison, WI, USA). Plasmid was extracted by a QIAprep Spin Miniprep Kit (Qiagen) and se- quenced by an Applied Biosystems (ABI) 3130 Genetic Analyzer (Jeonju AgroBio-Materials Institute, South Korea).
Both plasmids (including pCold II vector) were digested
with appropriate restriction enzymes (Nde I and Xba I) sepa-
rately for 1 hr at 37℃. Digested products were purified using
the QIAquick Gel Extraction Kit (Qiagen), ligated into pCold
II (Clontech), and transformed using One Shot TOP10 Competent cells for E. coli (Invitrogen, Carlsbad, CA, USA).
Plasmids were extracted using the QIAprep Spin Miniprep Kit (Qiagen), and were sequenced by the ABI 3130 Genetic Analyzer for confirmation.
Recombinant protein expression and purification Expression vectors were transformed into BL21 (DE3) ex- pression hosts (Novagen, Madison, WI, USA). A single col- ony on a Luria broth (LB) agar plate was cultured into 5 ml of LB medium with 100 μg/ml ampicillin at 37℃ over- night. The culture was added to 500 ml of LB medium with 100 μg/ml ampicillin, and was grown until the host cells reached optical density at 600 nm (OD600) of 0.6-0.8.
Recombinant proteins were induced by adding 1 mM Isopropyl β-D-1-thiogalactopyranoside (IPTG) at 15℃ for 24 hr. Cells were harvested by centrifugation at 4℃, 12,000 g for 10 min, resuspended in native lysis buffer (50 mM mono- sodium phosphate (NaH
2PO
4), 300 mM sodium chloride (NaCl), 10 mM imidazole, pH 8.0) or denaturing lysis buffer (100 mM NaH
2PO
4, 10 mM Tris-Cl, 8 M urea, pH 8.0), and disrupted by sonication (10 cycles of 15 s sonication with 30 s pause) on ice. Cell debris was removed by cen- trifugation at 4℃, 12,000 g for 20 min. Supernatants were combined with nickel-charged resin (Ni-NTA Superflow, Qiagen) and incubated for 2 hr by shaking at 200 rpm on ice. The suspension was loaded into a disposable poly- propylene column (Thermo Fisher Scientific, Rockford, IL, USA) and washed four times with wash buffer (soluble con- dition: 50 mM NaH
2PO
4, 300 mM NaCl, 20 mM imidazole, pH 8.0; insoluble condition: 100 mM NaH
2PO
4, 10 mM Tris-Cl, 8 M urea, pH 6.3). Subsequently, the proteins were eluted with elution buffer (soluble condition: 50 mM NaH
2PO
4, 300 mM NaCl, 250 mM imidazole, pH 8.0; in- soluble condition: 100 mM NaH
2PO
4, 10 mM Tris-Cl, 8 M urea, pH 4.5). The eluted proteins were subjected to 14%
sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE). After electrophoresis, gels were fixed and stained with 0.1% Coomassie Brilliant Blue R-250 dye. For Western blotting analysis, the gel was transferred into a pol- yvinylidene fluoride (PVDF) membrane at 30 V overnight on ice. After transferring, the PVDF membrane was blocked in Tris-buffered saline with Tween 20 (TBST)-1% skim milk for 1 hr, and was subsequently incubated with anti-human FGF-basic (FGF2) monoclonal antibodies (1:1,000) (Pepro Tech, Rocky Hill, NJ, USA) in TBST-1% skim milk at 4℃
overnight. After incubation, the membrane was washed twice with TBST for 5 min in a rotary shaker, and was in- cubated with anti-mouse alkaline phosphatase (IgG-AP)- conjugate secondary antibodies (1:30,000) (Sigma-Aldrich, St.
Louis, MO, USA) in TBST-1% skim milk at room temper- ature (RT) for 1 hr. The membrane was washed twice with TBST for 10 min in a rotary shaker, incubated with BCIP/
NBT substrate solution (Sigma-Aldrich) for 5 min, and de- veloped in a dark room.
Refolding of inclusion bodies
The protein concentration of purified inclusion bodies was measured by the Pierce BCA Protein Assay Kit (Thermo Fisher Scientific). The protein (1 mg/ml) was diluted 10 times (final conc. = 0.1 mg/ml) in refolding buffer (50 mM Tris-HCl, 120 mM guanidin hydrochloride, 0.045 mM re- duced glutathione, 0.45 mM oxidized glutathione, pH 8.5).
After vigorous mixing for 2 min, the sample underwent the reshuffling of disulfide bonds and oxidation with gentle stir- ring at 4℃ overnight. Subsequently, the last dialysis was re- peated against PBS buffer overnight. Finally, the sample was centrifuged at 4℃, 12,000 g for 20 min to remove aggregated proteins, and properties were determined using the Pierce BCA Protein Assay Kit (Thermo Fisher Scientific).
Immunofluorescence staining of mouse skin tissue Male SKH-1 hairless mice were purchased from ORIENT BIO Inc. (Iksan, South Korea). The mice were admitted to the experimental animal facility at the Jeonju AgroBio- Materials Institute in an environment of constant temper- ature (22±2℃) and humidity (55±5%), under a photoperiod of 12:12 hr (light:dark), for at least one week prior to experimentation. All procedures were in compliance with the guidelines of the Korean Council on Animal Care, and research protocols were approved by the Animal Committee of the Jeonju AgroBio-Materials Institute (JAMI IACUC 2016002). To confirm skin permeability of recombinant FGF2 or PTD-FGF2, the mice were divided into four groups (group 1, PBS-treated as blank; group 2, FGF2-treated as control;
group 3, Pep-1-FGF2-treated; group 4, FGF2-Pep-1-treated).
In the protein-treatment groups, recombinant proteins (250
μg/ml) were applied on the dorsal skin of the mice for 1
hr at RT. The mice skin absorption sites were dissected for
histological examination. Tissues were frozen in an optimal
cutting temperature (OCT) compound (Sakura FineTek
Europe B.V., Netherlands), and 7 mm sections were made.
A B
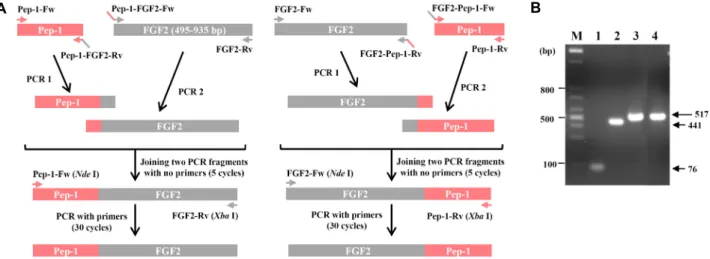
Fig. 1. cDNA synthesis of FGF2 and PTD-FGF2 fusion proteins. (A) Schematic diagram of OE-PCR. (B) Agarose gel analysis of PCR product of PTD, FGF2, and PTD-conjugated FGF2. M, DNA ladder markers; Lane 1, Pep-1; Lane 2, FGF2; Lane 3, Pep-1-FGF2; Lane 4, FGF2-Pep-1. The arrow on the right indicates the size of fragments.
The sections were thaw-mounted on slides at room temper- ature. The sections on slides were fixed in ethanol for 15 min, permeabilized with 0.25% Triton X-100 for 10 min, and blocked with 1% Bovine serum albumin (BSA) for 30 min.
The sections were exposed to anti-human FGF-basic mono- clonal antibodies (1:100) at 4
oC overnight. The sections were washed in phosphate-buffered saline (PBS) and incubated in anti-mouse IgG-FITC (Santa Cruz Biotechnology, Inc., Santa Cruz, CA, USA) (1:100) for 1 hr at RT in dark con- ditions. Following washes in PBS, the sections were in- cubated in blue-fluorescent DNA stain DAPI (4',6-dia- midino-2-phenylindole; Thermo Fisher Scientific) for 15 min.
Following additional washing with PBS and wet mounting, visualization and localization of recombinant proteins in the skin tissue were performed by laser scanning confocal mi- croscopy (LSM 510 META, Carl Zeiss, Germany).
Results and Discussion
Gene cloning and generation of expression vectors To explore efficient delivery methods of human basic fi- broblast growth factor (FGF2), previously known as FGFb (fibroblast growth factor 2 (basic)) [20], we generated FGF2 and FGF2 expression vectors with Pep-1 fused to the N- or C-terminus (amphipathic peptide of 21 amino acids; KET WWETWWTEWSQPKKKRKV), which has been reported to efficiently deliver a variety of active peptides and proteins into cells [13]. Overlap extension PCR (OE-PCR) was per- formed as shown in Fig. 1A, with expected sizes analyzed by agarose gel (Fig. 1B), representing Pep-1 (76 bp), FGF2
(441 bp), and Pep-1-conjugated FGF2 (517 bp), respectively.
Thereafter, we cloned the cDNA sequences encoding FGF2 and FGF2 with Pep-1 fused to the N- or C-terminus into pCold II expression vectors, which contained His-tag coding sequences prior to multicloning sites (MCSs) and the cold shock protein A (cspA) promoter for driving high expression and improving solubility proteins by cold shock in E. coli [32].
Expression of recombinant FGF2 and PTD-FGF2 fusion proteins
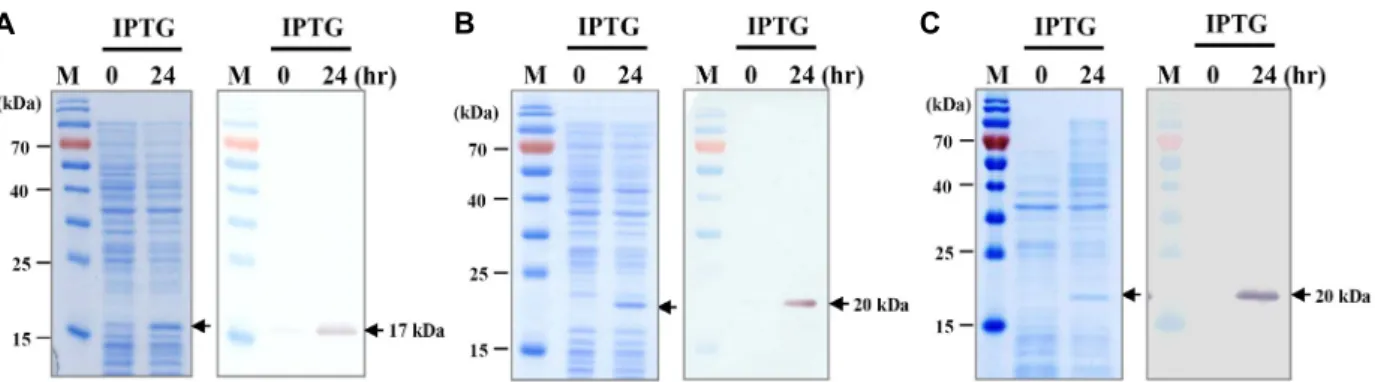
To express recombinant FGF2 and Pep-1-conjugated FGF 2, pCold II-FGF2, pCold II-Pep1-FGF2, and pCold II-FGF2- Pep1 were constructed, and expression of recombinant pro- teins was attempted in the bacterial expression system. To examine expression patterns and levels of FGF2 or Pep-1- conjugated FGF2, protein synthesis in E. coli cells with or without IPTG was analyzed via SDS-PAGE and Western blotting. The FGF2 single protein was presented as a single band of 17 kDa, as shown in Fig. 2A, while FGF2 fusion proteins showed 20 kDa molecular weight in E. coli (Fig.
2B, Fig. 2C). In order to investigate the solubility of recombi- nant proteins before large-scale expression, recombinant proteins were expressed at intervals of 0, 6, 12, and 24 hr.
Recombinant FGF2 (Fig. 3A) and FGF2 with Pep-1 fused to
C-terminus (Fig. 3C) were observed in soluble conditions,
whereas FGF2 with Pep-1 fused to N-terminus (Fig. 3B) was
expressed in insoluble conditions, which formed inclusion
bodies and may be precipitated. Fibroblast growth factors
(FGFs), such as FGF1, 2, 3, 6, 7, 8, 10, 16, 17, 20, and 22,
A B C
Fig. 2. Expression of FGF2 and PTD-FGF2 fusion proteins in E. coli. SDS-PAGE (left) and Western blotting analysis (right) of recombi- nant (A) FGF2, (B) Pep-1-FGF2, and (C) FGF2-Pep-1. Recombinant FGF2 proteins were identified using an anti-human FGF-ba- sic (FGF2) monoclonal antibody (1:1,000). M, protein molecular weight marker. The arrow on the right indicates the molecular weight of recombinant proteins.
were divided into three different groups (soluble, insoluble, and low-level expression) based on expression and solubility qualities in E. coli. As mentioned in previous studies, FGF2 was well expressed as soluble proteins [39]. Six histidine res- idues at the N-terminal end of FGF2 seemed not to interfere with its innate activity [19], indicating that the His-tag had no effect on the solubility and biological activity of FGF2.
However, it has been reported that human FGF2 (previously known as bFGF) fusion proteins with auxiliary colla- gen-binding decapeptide at the N-terminal end were ex- pressed as inclusion bodies, which may be due to the in- crease in hydrophobic interactions associated with extra de- capeptide fused to the N-terminus [2]. In our expression re- sults, recombinant His-FGF2 and His-FGF2-Pep1 were well expressed as soluble proteins, whereas His-Pep1-FGF2 fused to N-terminus was expressed as inclusion bodies, suggesting that extra amphipathic N-terminal residues between six his- tidine residues and FGF2 amino acids may have significant effects on aggregation of recombinant proteins in E. coli.
These findings urged us to determine whether extra N-ter- minal residues of FGF2 gave rise to insoluble conditions.
Thus we fused another known PTD, Antennapedia protein (Antp; RQIKIWFQNRRMKWKK) [9], to the N- and C-termi- nus of FGF2, respectively. In the same way, pCold II-Antp- FGF2 and pCold II-FGF2-Antp were constructed, and ex- pression of recombinant proteins was attempted at intervals of 0, 6, 12, and 24 hr. To determine the expression properties of Antp-FGF2 or FGF2-Antp, the expressed proteins were analyzed via SDS-PAGE. Interestingly, unlike the FGF2 with Antp fused to the C-terminus (Fig. 3E), the Antp-FGF2 with Antp fused to the N-terminus was also expressed in E. coli as insoluble (Fig. 3D). This result was consistent with the
results of FGF2 with Pep-1 fused to N-terminus. Thus the findings indicated that extra N-terminal residues of synthetic peptides, such as PTDs, may cause solubility problems in recombinant FGF2 proteins.
Purification of recombinant FGF2 and PTD-FGF2 fusion proteins
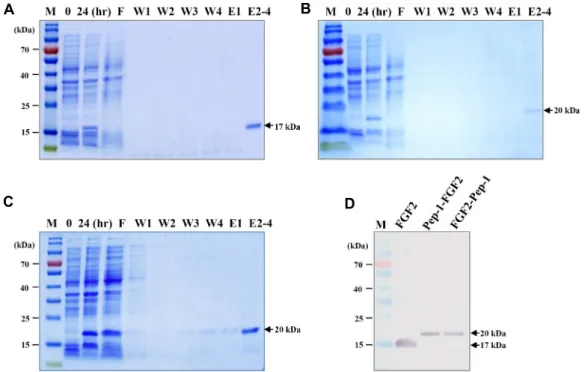
After expression in bacterial cells for 24 hr, while super- natants with soluble proteins were directly harvested follow- ing cell lysis by centrifugation, pellets with inclusion bodies were solubilized in denaturing lysis buffer containing strong denaturants, such as 6 M guanidine·HCl, and the super- natant was harvested by centrifugation. Recombinant pro- teins were purified in native conditions or denaturing con- ditions by using the 6×His-tag system at the N-terminal end.
Recombinant proteins in elution buffer were given single bands as 17-20 kDa molecular weight via SDS-PAGE, as shown in Fig. 4A, Fig. 4B, Fig. 4C, and Western blot analysis using monoclonal human FGF2 antibodies, as shown in Fig.
4D. The eluted samples were dialyzed by PBS for desalting
and concentration. However, the denatured Pep1-FGF2 fu-
sion proteins went through a renaturation (refolding) phase
by dialysis using glutathione (GSH) and glutathione disul-
fide (GSSG) for disulfide bond reshuffling and oxidation,
and subsequently exchanged buffer with PBS, according to
previous studies [2]. Despite our efforts, the Pep1-FGF2 was
lost during the refolding process, and there was some re-
version to insoluble proteins by reaggregation. Different
studies of recombinant human basic fibroblast growth factor
in E. coli have observed almost 5-60% loss [25, 33]. Some
loss might be unavoidable, and these types of fusion prod-
ucts might seem to have higher inclusion bodies due to
A B C
D E
Fig. 3. Investigation of solubility of FGF2 and PTD-FGF2 fusion proteins from E. coli. (A) SDS-PAGE of recombinant FGF2, (B) Pep-1-FGF2, (C) FGF2-Pep-1, (D) Antp-FGF2, and (E) FGF2-Antp expressed at 0, 6, 12, and 24 hr. M, protein molecular weight marker; S, soluble lysate fraction; I, Insoluble lysate fraction (inclusion body). The arrow on the right indicates the molecular weight of recombinant proteins.
A B
C D
Fig. 4. Purification of FGF2 and PTD-FGF2 fusion proteins. 6×His-tagged recombinant proteins were expressed for 24 hr and then purified on Ni-NTA Superflow under native or denaturing conditions. (A) SDS-PAGE of recombinant FGF2, (B) Pep-1-FGF2, (C) FGF2-Pep-1 proteins. (D) Western blotting analysis of purified recombinant proteins using an anti-human FGF-basic (FGF2) monoclonal antibody (1:1,000). The arrow on the right indicates the molecular weight of recombinant proteins. M, protein molecular weight marker; F, flow-through fraction; W, wash fractions; E, elute fractions.
high-level expression and extra N-terminal residues by syn- thetic peptides in E. coli. Although various refolding meth- ods have been performed in many studies [2-4, 17], refolding
is actually a very challenging task [25, 38]. Thus we per-
formed large-scale expression and repeated the same process
more than twice to obtain the proper concentration of
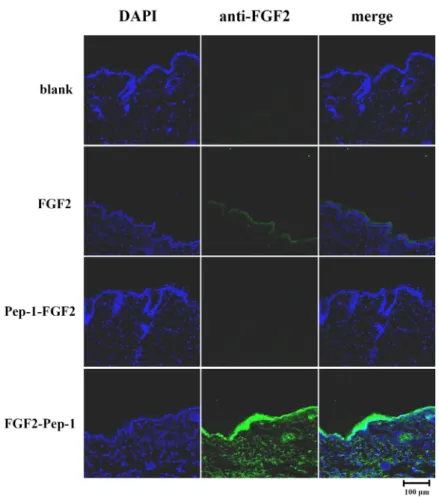
Fig. 5. Immunofluorescence staining for trans- duction ability of FGF2 and PTD-FGF2 fusion proteins. Tissue sections of mice dorsal skins were stained with an an- ti-human FGF-basic (FGF2) monoclonal antibody (1:100) (green) and with DAPI (blue). Merged confocal images are shown in the third column. Scale bar corre- sponds to 100 μm.
Pep-1-FGF2, which is the one and only inclusion body in
E. coli observed herein.Transduction ability of recombinant FGF2 and PTD- FGF2 fusion proteins
Protein transduction domains (PTDs), such as Trp-rich motif-SV40 NLS Pep-1, HIV transcription factor TAT, the
Drosophila homeotic transcription factor Antp, and herpessimplex virus transcription factor VP22 [9, 10, 13, 34], may be useful tools to deliver recombinant therapeutic or cos- metic proteins across various mammalian cell types. Actually, proteins and peptides that are efficiently fused to PTDs have been transduced across cell membranes, as well as cytosol and the nucleus of cells [23, 31, 35].
To assess skin permeability of FGF2 or FGF2 with Pep-1 fused to the N- or C-terminus, purified recombinant proteins were treated on shaved areas of the dorsal skin of mice subjects. Immunofluorescence staining was used to de- termine whether the recombinant proteins were delivered into skin tissues. Immunofluorescence staining is employed to visualize antibodies that bind to antigens using a fluo- rophore in tissue. FGF2-Pep-1 in particular was observed in
the top layer of the skin (epidermis) and in the layer of skin beneath the epidermis (dermis), while FGF2 control proteins were not detected in dermis of animal subjects, indicating that FGF2-Pep-1 (fused to C-terminus) was efficiently deliv- ered into the skin tissues of mice subjects (Fig. 5).
In contrast, Pep-1-FGF2 (fused to N-terminus) proteins were not observed in epidermis or in dermis of the skin of subjects (Fig. 5), suggesting that the permeability of FGF2 by extra N-terminus of amphipathic peptides might be inef- fective because it is ultimately hard to overcome an activity barrier. Our findings show that FGF2 with Pep-1 fused to the C-terminus is more efficiently expressed, purified, and delivered into skin tissues than FGF2 with Pep-1 fused to the N-terminus, without labor-intensive, costly, and time- consuming methods.
Acknowledgement
This research was supported by the Korea Institute for
the Advancement of Technology (KIAT) of the Ministry of
Trade, Industry, & Energy (MOTIE) through the Encourage-
ment Program for the Industries of Economic Cooperation
Region (grant number R0004670).
References
1. Alvarez, E., Fey, E. G., Valax, P., Yim, Z., Peterson, J. D., Mesri, M., Jeffers, M., Dindinger, M., Twomlow, N., Ghatpande, A., LaRochelle, W. J., Sonis, S. T. and Lichen- stein, H. S. 2003. Preclinical characterization of CG53135 (FGF-20) in radiation and concomitant chemotherapy/ radi- ation-induced oral mucositis. Clin. Cancer Res. 9, 3454-3461.
2. Andrades, J. A., Santamaría, J. A., Wu, L. T., Hall, F. L., Nimni, M. E. and Becerra, J. 2001. Production of a recombi- nant human basic fibroblast growth factor with a collagen binding domain. Protoplasma 218, 95-103.
3. Andrades, J. A., Wu, L. T., Hall, F. L., Nimni, E. and Becerra, J. 2001. Engineering, expression, and renaturation of a colla- gen-targeted human bFGF fusion protein. Growth Factors 18, 261-275.
4. Alibolandi, M. and Mirzahoseini, H. 2011. Purification and refolding of overexpressed human basic fibroblast growth factor in Escherichia coli. Biotechnol. Res. Int. 973741, 1-6.
5. An, J. J., Eum, W. S., Kwon, H. S., Koh, J. S., Lee, S. Y., Baek, J. H., Cho, Y. J., Kim, D. W., Han, K. H., Park, J., Jang, S. H. and Choi, S. Y. 2013. Protective effects of skin permeable epidermal and fibroblast growth factor against ultraviolet-induced skin damage and human skin wrinkles.
J. Cosmet. Dermatol. 12, 287-295.
6. Bowden, G. A., Paredes, A. M. and Georgiou, G. 1991.
Structure and morphology of inclusion bodies in Escherichia coli. Bio/Technology 9, 725-730.
7. Bikfalvi, A., Klein, S., Pintucci, G. and Rifkin, D. B. 1997.
Biological roles of fibroblast growth factor-2. Endocr. Rev.
18, 26-45.
8. Carrió, M. M., Corchero, J. L. and Villaverde, A. 1998.
Dynamics of in vivo protein aggregation: building inclusion bodies in recombinant bacteria. FEMS Microbiol. Lett. 169, 9-15.
9. Derossi, D., Joliot, A. H., Chassaing, G. and Prochiantz, A.
1994. The third helix of the antennapedia homeodomain translocates through biological membranes. J. Biol. Chem.
269, 10444-10450.
10. Elliott, G. and O'Hare, P. 1997. Intercellular trafficking and protein delivery by a herpesvirus structural protein. Cell.
88, 223-233.
11. Frankel, A. D. and Pabo, C. O. 1988. Cellular uptake of the tat protein from human immunodeficiency virus. Cell 55, 1189-1193.
12. Garke, G., Deckwer, W. D. and Anspach, F. B. 2000. Prepar- ative two-step purification of recombinant human basic fi- broblast growth factor from high-cell-density cultivation of Escherichia coli. J. Chromatogr. B: Biomed. Sci. Appl. 737, 25- 38.
13. Gros, E., Deshayes, S., Morris, M. C., Aldrian-Herrada, G., Depollier, J., Heitz, F. and Divita, G. 2006. A non-covalent peptide-based strategy for protein and peptide nucleic acid
transduction. Biochim. Biophys. Acta. 1758, 384-393.
14. Gasparian, M. E., Elistratov, P. A., Drize, N. I., Nifontova, I. N., Dolgikh, D. A. and Kirpichnikov, M. P. 2009. Overex- pression in Escherichia coli and purification of human fibro- blast growth factor (FGF-2). Biochemistry (Mosc) 74, 221-225.
15. Hagiwara, A., Nakayama, F., Motomura, K., Asada, M., Suzuki, M., Imamura, T. and Akashi, M. 2009. Comparison of expression profiles of several fibroblast growth factor re- ceptors in the mouse Jejunum: Suggestive evidence for a differential radioprotective effect among major FGF family members and the potency of FGF1. Radiat. Res. 172, 58-65.
16. Iwane, M., Kurokawa, T., Sasada, R., Seno, M., Nakagawa, S. and Igarashi, K. 1987. Expression of cDNA encoding hu- man basic fibroblast growth factor in E. coli. Biochem. Biophys.
Res. Commun. 146, 470-477.
17. Knoerzer, W., Binder, H, P., Schneider, K., Grussc, P., McCarthyd, J. E. G. and Risaue, W. 1989. Expression of syn- thetic genes encoding bovine and human basic fibroblast growth factors (bFGF). Gene 75, 21-30.
18. Kuriyama, M., Nakatu, M., Nakao, M., Igarashi, K. and Kitano, K. 1992. Controlled expression of human basic fibro- blast growth factor mutein CS23 in Escherichia coli under a bacteriophage T7 promoter. J. Ferment. Bioeng. 74, 67-72.
19. Kroiher, M., Raffioni, S. and Steele, R. E. 1995. Single step purification of biologically active recombinant rat basic fi- broblast growth factor by immobilized metal affinity chro- matography. Biochim. Biophys. Acta. 1250, 29-34.
20. Kim, H. S. 1998. Assignment1 of the human basic fibroblast growth factor gene FGF2 to chromosome 4 band q26 by ra- diation hybrid mapping. Cytogenet. Cell Genet. 83, 73.
21. Lemaitre, G., Laaroubi, K., Soulet, L., Barritault, D. and Miskulin, M. 1995. Production and purification of active FGF2 via recombinant fusion protein. Biochimie 77, 162-166.
22. Lindgren, M., Hallbrink, M., Prochiantz, A. and Langel, Ü.
2000. Cell-penetrating peptides. Trends Pharmacol. Sci. 21, 99- 103.
23. Lindsay, M. A. 2002. Peptide-mediated cell delivery: appli- cation in protein target validation. Curr. Opin. Pharmacol. 2, 587-594.
24. Misawa, S. and Kumagai, I. 1999. Refolding of therapeutic protein produced in Escherichia coli as inclusion bodies.
Biopolymers 51, 297-307.
25. Mirzahoseini, H., Mehraein, F., Omidinia, E. and Razavi, M.
R. 2004. Differential expression of human basic fibroblast growth factor in Escherichia coli: potential role of promoter.
World. J. Microbiol. Biotechnol. 20, 161-165.
26. Maclachlan, T., Narayanan, B., Gerlach, V. L., Smithson, G., Gerwien, R. W., Folkerts, O., Fey, E. G., Watkins, B., Seed, T. and Alvarez, E. 2005. Human fibroblast growth factor 20 (FGF-20; CG53135-05): a novel cytoprotectant with radio- protective potential. Int. J. Radiat. Biol. 81, 567-579.
27. Nakayama, F., Muller, K., Hagiwara, A., Ridi, R., Akashi, M. and Meineke, V. 2008. Involvement of intracellular ex- pression of FGF12 in radiation-induced apoptosis in mast cells. J. Radiat. Res. 49, 491-501.
28. Nakayama, F., Hagiwara, A., Kimura, M., Akashi, M. and
초록:대장균 발현시스템에서 단백질 전달 도메인 PTD가 인간 섬유아세포 성장인자(FGF2)의 N- 또는 C-말단에 결합 되었을 때 미치는 재조합 단백질 복합체의 발현 특성과 피부 투과능력
박인선
1․최충현
2․권보라
1․최영지
1․권태호
2․유강열
1․이주형
3․추영무
1*
(1(재)전주농생명소재연구원, 2(주)엔비엠, 3전북대학교 예방의학교실)
인간 섬유아세포 성장인자는 조직 생성 및 상처 치료 효과로 인해 상업적으로 중요한 치료제 또는 화장품소재 로서 가능성이 높다. 피부 조직에 침투성을 부여하여 치료효과를 높이기 위해서 단백질 전달 도메인인 PTD를 FGF에 융합을 시도하고 있으며 피부로의 투과능력과 그로인한 치료 효과에 대한 연구도 진행되고 있다. 그러나, PTD가 FGF 단백질의 N- 또는 C-말단에 결합 되었을 때 PTD의 위치가 대장균 발현시스템에서 재조합단백질 접 힘 및 안정성, 그리고 결국 피부로의 도입능력에 상당한 영향을 미치는지에 대해서는 알려져 있지 않다. 여기에서 우리는 대조군으로 PTD가 융합되지 않은 인간 염기성 섬유아세포 성장인자(FGF2)와 PTD가 FGF2의 N-말단 또는 C-말단에 융합된 FGF2 복합체를 중합효소연쇄반응(OE-PCR)을 통해 클로닝 하였다. 그 다음 이들 재조합 FGF2의 단백질 발현 및 특성을 확인하고 마우스 등 피부를 이용하여 조직 내로의 도입 능력을 조사 하였다. 결과적으로, 불용성 PTD-FGF2 (N 말단 융합)와는 달리 대조군 FGF2와 FGF2-PTD 융합 단백질(C- 말단 융합)은 가용성 형태 로 발현되어 재조합단백질 획득이 용이하였고, 마우스 피부 도입능력은 FGF2-PTD 융합단백질에서만 나타내 보 였다. 우리의 결과는 C-말단에 융합된 FGF2-PTD 융합단백질이 발현, 정제, 피부 투과능력의 측면에서 다른 두 옵션들보다 노동, 비용, 시간면에서 보다 더 효율적일 수 있음을 시사한다.
Imamura, T. 2009. Evaluation of radiation-induced hair fol- licle apoptosis in mice and the preventive effects of fibro- blast growth factor-1. Exp. Dermatol. 18, 889-892.
29. Nakayama, F., Hagiwara, A., Umeda, S., Asada, M., Goto, M., Oki, J., Suzuki, M. and Imamura, T. 2010. Post treatment with an FGF chimeric growth factor enhances epithelial cell proliferation to improve recovery from radiation-induced intestinal damage. Int. J. Radiat. Oncol. Biol. Phys. 78, 860- 867.
30. Nakayama, F., Yasuda, T., Umeda, S., Asada, M., Imamura, T., Meineke, V. and Akashi, M. 2011. Fibroblast growth fac- tor-12 (FGF12) translocation into intestinal epithelial cells is dependent on a novel cell-penetrating peptide domain. J.
Biol. Chem. 286, 25823-25834.
31. Papadopoulou, L. C. and Tsiftsoglou, A. S. 2013. The poten- tial role of cell penetrating peptides in the intracellular de- livery of proteins for therapy of erythroid related disorders.
Pharmaceuticals 6, 32-53.
32. Qing, G., Ma, L. C., Khorchid, A., Swapna, G. V. T., Mal, T. K., Takayama, M. M., Xia, B., Phadtare, S., Ke, H., Acton, T., Montelione, G. T., Ikura, M. and Inouye, M. 2004.
Cold-shock induced high-yield protein production in Escher- ichia coli. Nat. Biotechnol. 22, 877-882.
33. Rinas, U., Tsai, L. B., Lyons, D., Fox, G. M., Stearns, G., Fieschko, J., Fenton, D. and Bailey, J. E. 1992. Cysteine to serine substitutions in basic fibroblast growth factor: effect on inclusion body formation and proteolytic susceptibility during in vitro refolding. Nat. Biotechnol. 10, 435-440.
34. Rosano, G. L. and Ceccarelli, E. A. 2014. Recombinant pro-
tein expression in Escherichia coli: advances and challenges.
Front Microbiol. 5, 172.
35. Schwarze, S. R., Ho, A., Vocero-Akbani, A. and Dowdy, S.
F. 1999. In vivo protein transduction: Delivery of a bio- logically active protein into the mouse. Science 285, 1569- 1572.
36. Schwarze, S. R., Hruska, K. A. and Dowdy, S. F. 2000. Protein transduction: Unrestricted delivery into all cells? Trends Cell Biol. 10, 290-295.
37. Sheng, Z., Chang, S. B. and Chirico, W. J. 2003. Expression and purification of a biologically active basic fibroblast growth factor fusion protein. Protein. Expr. Purif. 27, 267-271.
38. Song, J. A., Koo, B. K., Chong, S. H., Kwak, J., Ryu, H. B., Nguyen, M. T., Vu, T. T. T., Jeong, B., Kim, S. W. and Choe, H. 2013. Expression and purification of biologically active human FGF2 containing the b‘a’ domains of human PDI in Escherichia coli. Appl. Biochem. Biotechnol. 170, 67-80.
39. Sun, C., Li, Y., Taylor, S. E. and Marston, F. A. O. 2015.
Halo Tag is an effective expression and solubilisation fusion partner for a range of fibroblast growth factors. PeerJ 3, e1060.
40. Taylor, G., Hoare, M., Gray, D. R. and Marston, F. A. O.
1986. Size and density of inclusion bodies. Bio/Technology 4, 553-557.
41. Wang, J., Hong, A., Ren, J. S., Suna, F. Y., Shia, Y. J., Liua, K., Xiea, Q. L., Daia, Y., Lia, Z. Y. and Chen, Y. 2006.
Biochemical properties of C78SC96S rhFGF-2: a double point-mutated rhFGF-2 increases obviously its activity. J.
Biotechnol. 121, 442-447.