Article
http://dx.doi.org/10.4217/OPR.2017.39.2.115
여정실의 항산화 활성
서영완1,2* · 김호준1
1한국해양대학교 해양생명과학부
2한국해양대학교 해양과학기술전문대학원 (49112) 부산광역시 영도구 동삼동 1
Antioxidant Activity of Fruits of Ligustrum japonicum
Youngwan Seo1,2* and Hojun Kim1
1Division of Marine Bioscience, Korea Maritime and Ocean University
2Ocean Science and Technology School, Korea Maritime and Ocean University Busan 49112, Korea
Abstract : The objective of this study is to evaluate the antioxidant activity of the fruits of Ligustrum japonicum. The crude extract was successively fractionated into n-hexane, 85% aqueous methanol (85%
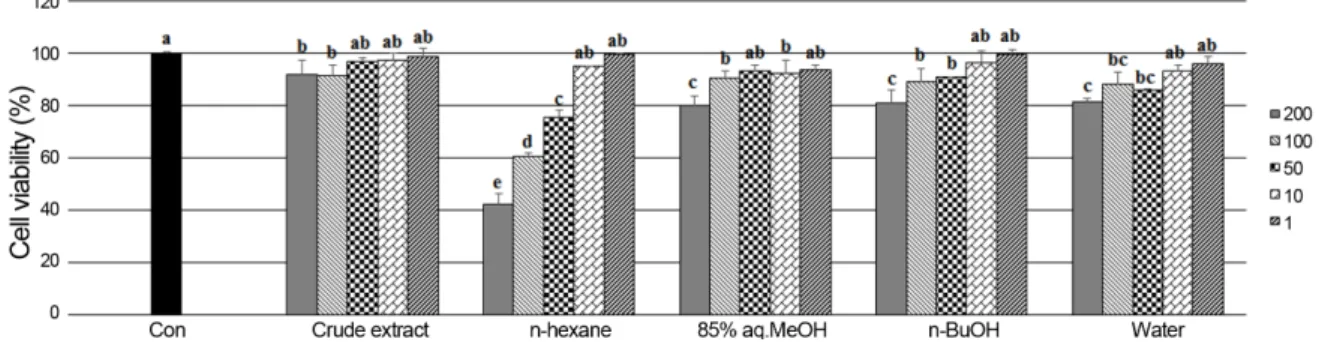
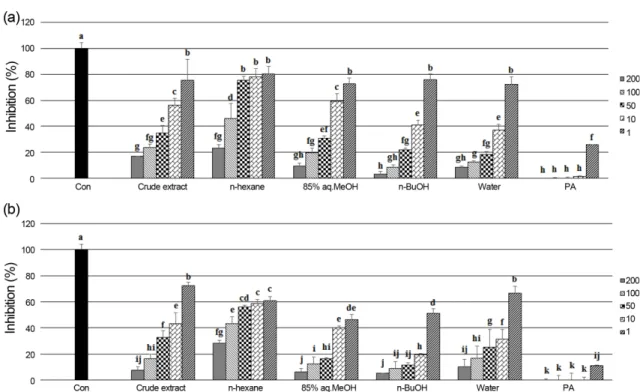
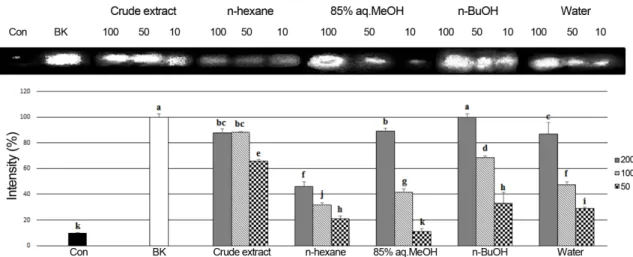
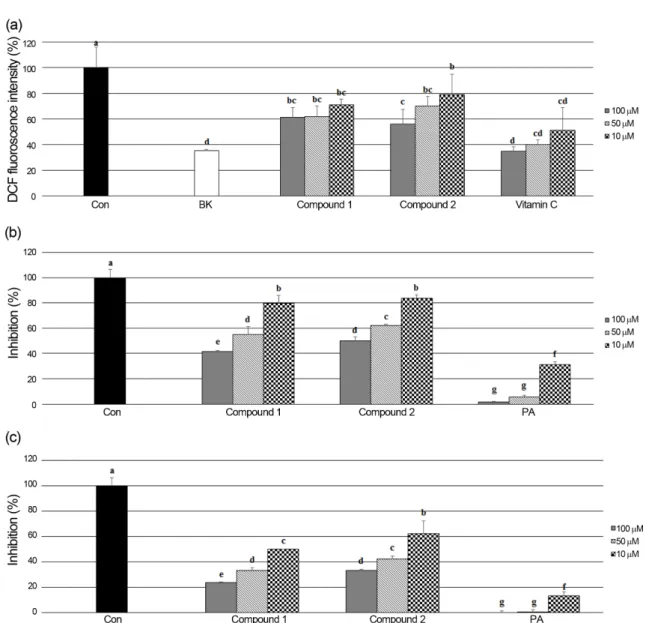
aq.MeOH), n-butanol (n-BuOH), and water fractions by means of solvent polarity. The crude extract and its solvent fractions were evaluated for their antioxidant effect by four different assay systems: scavenging power on peroxynitrite and intralcellular ROS produced in HT-1080 cells; DNA oxidation inhibition; ferric reducing antioxidant power (FRAP). The n-BuOH fraction exhibiting potent antioxidant activity was further purified by C18 silica gel column chromatography and RP-HPLC to give tyrosol (1) and salidroside (2).
The structure of isolated compounds was determined by extensive 2 D NMR experiments such as
1H COSY, NOESY, HSQC and HMBC as well as by comparison with the published spectral data.
Key words : Ligustrum japonicum, antioxidant, reactive oxygen species (ROS), reactive nitrogen species (RNS)
1. 서 론
생체내에서는 내인성 혹은 외인성 자극에 의해서 활성 산소종(reactive oxygen species, ROS)과 활성질소종 (reactive nitrogen species) 이 생성되고 생체내 항산화방어 시스템은 이들이 과다하게 생성되는 것을 억제함으로써 산화환원 반응의 평형을 유지하고 생체 손상을 막는다 (Ziech et al. 2010; Persson et al. 2014). 활성산소종이나 활성질소종은 산소호흡에 의해서 생성되거나 산화환경에 노출되면 생성되기 때문에 원천적으로 이것들의 생성을
차단하기는 어렵다. 낮은 농도로 존재하는 활성산소종은 세포증식, 아폽토시스, 유전자 발현등과 밀접하게 관련되 어 있으며 대식세포내에서 생성되는 활성산소종은 박테리 아, 곰팡이와 같은 외부에서 침입하는 병원성 미생물을 죽 이는데 필수적이다(Poljsak et al. 2013). 하지만 이러한 평 형이 깨어지고 활성산소종이나 활성질소종이 과다생성될 때 산화스트레스가 발생된다(Lopez-Alarcona and Denicola 2013; Pisoschi and Pop 2015). 산화스트레스의 심각성은 심혈관질환, 신경퇴행성 질환, 알츠하이머, 파킨슨, 루게 릭, 폐기종, 심혈관 질환, 염증성 질환, 백내장, 암, 노화등 과 같은 많은 만성적인 건강문제의 발병에 관여하며 100가지 이상의 질병과 연관성이 있다는 것으로 알려져
*Corresponding author. E-mail : [email protected]