Agric. Chem. Biotechnol. 47(3), 157-160 (2004)
Note
Expression of Thermotoga maritima Endo- β-1,4-xylanase Gene in E. coli and Characterization of the Recombinant Enzyme
Hyang-Sik Yoon1, Nam Soo Han2 and Chung Ho Kim*
Department of Food and Nutrition, Seowon University, Cheongju 361-742, Korea
1Team for Food Research & Development, Chungcheongbuk-do Agricultural Research and Extension Services, 363-883, Korea
2Department of Food Science and Technology, Chungbuk National University, Cheongju 361-763, Korea Received July 27, 2004; Accepted August 19, 2004
Gene encoding endo-1,4-β-xylanase (EC 3.2.1.8) was isolated from hyperthermophilic microorganism, Thermotoga maritima by polymerase chain reaction. Open reading frame (ORF) of xylanase gene is 3,180 bp long and encodes 1,059 amino acid residues. Estimated molecular weight of polypeptide is 119,642 Da. To analyze enzymatic activity and biochemical properties of xylanase, ORF of xylanase gene was introduced into E. coli expression vector, and overexpressed in E. coli BL21. Purified recombinant xylanase showed highest activity at 90oC, and pH 6.0, and was stable below 90oC. D-xylose- and xylooligosacharide-producing activities of purified recombinant xylanase was tested using oatspelts arabinoxylan and birchwood xylan as substrates. Enzyme activity of recombinant enzyme was 170 U/ml on oatspelts arabinoxylan.
Key words: D-xylose, xylooligosaccharide, endo-β-1,4-xylanase, thermostable enzyme, Thermotoga maritima
Lignocellulosic biomass is a constant source of sustainable and renewable energy in nature and is comprised of about 23% lignin, 40% cellulose, and 33% hemicellulose.1) The monomers of various lignocellulosic agricultural and industrial wastes are useful in the production of different antibiotics, alcohols, animal feed, chemicals and fuels.2,3)
Xylan, a major component of hemicellulose and, after cellulose, the second most abundant polysaccharide in nature4) is found in large amounts in wood and plant material. It consists of a backbone of D-xylose residues, and often has α- L-arabinofuranose and glucuronic acid side chains. Most xylans are also acetylated. Degradation of xylan requires the action of several different enzymes. Among these, endo-1,4- β-xylanase (EC 3.2.1.8) is the most important because it initiates the degradation of xylan into xylooligosaccharides.5)
Xylanases from various microorganisms including fungi and yeasts have been isolated and characterized.6-9) Several xylanases, mainly from Bacillus spp., have been cloned and expressed in Escherichia coli10-13) but with only a few from thermophilic organisms.10,13) Xylanases, which showed optimum enzymatic activity between 70 to 80oC, have been reported.14-16) However, none of the enzymes was stable if kept long term at above 65oC.
The extreme environment inhabited by Thermotoga maritima (growth range 55-80oC with an optimum at 80oC)17) has imposed severe selective pressure upon the enzymatic outfit of this microorganism, which must be sufficiently
thermophilic as well as thermostable to guarantee survival at very high temperatures. Therefore, T. maritima appears to be appropriate for the isolation of highly thermostable enzymes, whose biochemical and structural analyses may be extremely helpful in unraveling the molecular basis of thermostability.
Additionally, the thermophilic enzymes produced by this organism may prove to be valuable catalysts for industrial applications.
According to GenBank analysis, a hyperthermophilic microorganism, Thermotoga maritima, is known to have a gene encoding xylanase. With optimal and maximal growth temperatures of 80 and 90oC, respectively, it represents one of the most thermophilic eubacterial genus yet described, as well as being one of the most ancient.18) In this study, the gene encoding xylanase was cloned from Thermotoga maritima, and expressed in E. coli through recombination into pRSET expression vector. The biochemical properties, and D-xylose- and D-xylooligosaccharide-producing activities of recombinant xylanase were also characterized.
Materials and Methods
Bacterial strains. Chromosomal DNA of T. maritima (ATCC 43589D) was purchased from ATCC to clone the xylanase-encoding gene. E. coli TOP10 and E. coli BL21 grown in LB19) were used for transformation and plasmid propagation, and for the expression of recombinant xylanase gene.
Enzymes and chemicals. Restriction and DNA-modifying enzymes were purchased from Promega. All enzymes were used as recommended by their manufactures. Ni2+-NTA- agarose was purchased from Qiagen. Taq DNA polymerase
*Corresponding author
Phone: +82-43-299-8745; Fax: +82-43-299-8745 E-mail : [email protected]
158 Hyang-Sik Yoon et al.
and dNTPs were purchased from Boehringer Mannheim.
Other chemicals were purchased from Sigma Chemical Co.
Construction of expression plasmid. Open reading frame (ORF) of xylanase gene was introduced into the pRSET-B E.
coli expression vector through PCR amplification. The xylanase-specific primers were 5'-CTTCGCGGATCCGATG CAAGTCAGGAAGAGA-3' and 5'-GAAGCGGGATCCTC ACTTGATGAGCCTGAG-3' containing translation initiation and termination codons (underlined), respectively. PCR product was digested with BamHI and subcloned into the pRSET-B vector to produce pRBTmXyl. Polymerase chain reaction was carried out in a volume of 50µl containing 10 mM Tris-HCl/pH 8.3, 1.5 mM MgCl2, 0.2 mM dNTPs, 100 ng of template DNA, 150 ng of each primer, and 2.5 units of Taq DNA polymerase. DNA was amplified as follows: 5 min at 94oC; followed by 30 cycles of 1 min at 94oC, 1 min at 52oC, 2 min at 72oC; and final extension at 72oC for 10 min.
Expression of xylanase gene in E. coli. pRBTmXyl was transformed into E. coli BL21, and induced by adding 1 mM IPTG at 37oC for 4 h. The cell pellet was resuspended in 4 ml of lysis buffer (50 mM NaH2PO4/pH 8.0, 300 mM NaCl, 10 mM imidazole) and sonicated for three cycles of 4 min each on ice. Crude extract of the recombinant enzyme was purified by Ni2+-NTA-agarose column chromatography and eluted with a 10 to 250 mM imidazole gradient. Protein samples were analyzed through discontinuous SDS-polyacrylamide gel electrophoresis,20) and their protein concentrations were determined by the Bradford method using bovine serum albumin as a standard.21)
Enzyme assay of xylanase activity. Assay of xylanase activity was carried out by the methods of Winterhalter et al.22) Xylanase activity was assayed at a reaction volume of 100µl containing 0.5% of oatspelts arabinoxylan, 50 mM sodium phosphate buffer (pH 6.0), and purified xylanase. After incubation at 90oC for 10 min, reaction was stopped by addition of two volumes of dinitrosalicylic acid reagent, and the resulting color developed as a result of xylose liberation was measured at 550 nm. One unit of xylanase activity was defined as the amount of enzyme required to liberate 1µmol of xylose reducing-sugar equivalent per min in the reaction mixture under the assay conditions.
Determination of biochemical properties of xylanase.
The effects of pH on the activity of xylanase were determined in a series of 50 mM each sodium citrate buffer with pH values from 3.0 to 6.0, and 50 mM phosphate buffer with pH values from 6.0 to 8.0. The effects of temperature on the activity and stability of xylanase were determined at temperatures ranging from 60 to 110oC. The activity of xylanase was assayed by the standard method described above.
Analysis of sugars. Sugar analysis was carried out by TLC. The reaction mixture was spotted onto a silica gel TLC plate (F254; Merck) and developed three times in a solution containing acetonitrile and water (85 : 15). The sugars were visualized by dipping the TLC plate into a solution containing 5% (v/v) H2SO4 and 0.5% (w/v) α-naphthol in ethanol,
followed by baking at 110oC for 10 min.
Results and Discussion
Isolation and expression of a gene for xylanase. To isolate the gene encoding xylanase from T. maritima, PCR was carried out using xylanase-specific primers, which were designed according to the GenBank analysis (AE001693).
The PCR product was digested with BamHI and introduced into the pRSET-B vector to produce pRBTmXyl. The ORF of xylanase gene is 3,180 bp long and encodes 1,059 amino acid residues with a molecular weight of 119,462 Da. The deduced amino acid sequence of xylanase had 91, 87, 43 and 38%
identities with xylanases from Thermotoga sp. FjSS3-B.1 (AAD32593), T. neapolitana (CAA87069), Caldicellulosiruptor sp. Rt69B.1 (AAB95325) and Thermoanaerobacterium thermosulfurigenes (AAB08046), respectively.
To analyze the enzymatic activity and the substrate specificities of xylanase, pRBTmXyl was transformed, expressed in E. coli BL21, and analyzed through discontinuous SDS-PAGE. Recombinant xylanase was expressed in E.
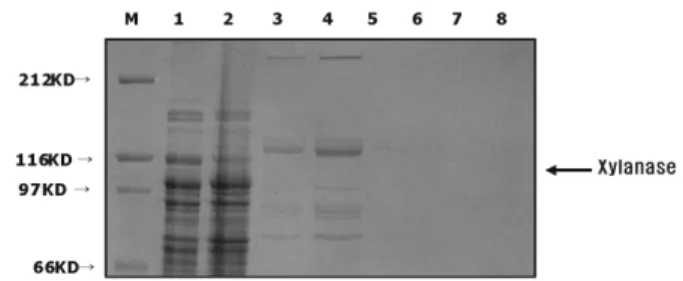
coli BL21, and purified by Ni2+-NTA-agarose column chromatography (Fig. 1). The estimated molecular weight of recombinant xylanase containing hexahistidine domain derived from the pRSET vector was approximately 120,000 Da, in agreement with the value calculated from the deduced amino acids. The enzyme activity of the recombinant enzyme was 170 U/ml on oatspelts arabinoxylan.
Biochemical properties of recombinant xylanase. The effects of temperature and pH on the activity of recombinant xylanase are shown in Fig. 2. A 0.5% arabinoxylan was incubated for 10 min with 0.17 U of purified recombinant xylanase at various temperatures and pHs, and the changes in color were measured at 550 nm.
The effects of temperature on the activity of recombinant xylanase were tested at pH 6.0 from 60 to 110oC. The recombinant xylanase showed the highest activity at around 90oC (Fig. 2A). Thermal stability of the recombinant xylanase
Fig. 1. Expression of recombinant xylanase in E. coli. ORF of T. maritima xylanase gene was introduced into E. coli expression vector, pRSET-B and expressed in E. coli. Protein extracts prepared from induced E. coli and their purified recom- binant enzymes were analyzed by 8.5% SDS-PAGE and stained with Coomassie Blue. Lane M: Molecular weight marker. Lane 1: total extract of E. coli harboring pRSET-B plasmid only.
Lane 2: total extract of E. coli harboring pRBTmXyl. Lane 3-8:
purified recombinant xylanase eluted by Ni2+-NTA-agarose affin- ity column
Expression of Thermotoga maritima Xylanase Gene in E. coli 159
was tested. Residual activity after preincubation between 60- 110oC (in the absence of substrate) was determined as a function of the incubation time. The enzyme was stable below 80oC with a half-life of activity of about 1 hr at 90oC (data not shown).
The effects of pH on the activity of recombinant xylanase were tested at 90oC in the pH range 3.0 to 8.0. The recombinant xylanase showed the highest activity at around pH 6.0 (Fig. 2B).
Recombinant xylanase produces xylooligosaccharides from arabinoxylan. The D-xylose- and xylooligosacharide- producing activities of the purified recombinant xylanase were tested using oatspelts arabinoxylan and birchwood xylan as the substrates. Purified recombinant xylanase (0.85 U) was incubated with 2.5% (w/v) oatspelts arabinoxylan, and the reaction was carried out at 90oC for 30 min. Various xylooligosaccharides were released from oatspelts arabinoxylan (Fig. 3). The reaction mixtures contained xylobiose as the main reaction product, xylotriose, and various xylooligosaccharides.
Furthermore, when the incubation time was extended to 4 days, very low level of xylose was also detected, and most of the xylotriose was converted into xylobiose (data not shown).
These results demonstrated that D-xylose and xylooligosaccharide could be produced from arabinoxylan using recombinant xylanase. Studies on the substrate specificity and other biochemical properties of recombinant xylanase on natural polysaccharides are under progress.
References
1. Aspinall, G. O. (1959) Structural chemistry of the hemicel- lulose. Adv. Carbohydr. Chem. 14, 429-468.
2. Wyman, C. E. (1994) Ethanol from lignocellulosic biom- ass: technology, economics and opportunities. Bioresour.
Technol. 50, 3-16.
3. Beg, Q. K., Kapoor, M., Mahajan, L. and Hoondal, G. S.
(2001) Microbial xylanases and their industrial applica- tions: a review. Appl. Microbiol. Biotechnol. 56, 326-338.
4. Luthi, E., Jasmat, N. B. and Bergquist, P. (1990) Xylanase from the extremely thermophilic bacterium “caldocellum saccharolyticum”: Overexpression of the gene in Escheri- chia coli and characterization of the gene product. Appl.
Environ. Microbiol. 56, 2677-2683.
5. Biely, P. (1985) Microbial xylanolytic systems. Trends Bio- technol. 3, 286-290.
6. Desphande, V., Lachke, A., Mishra, C., Keskar, S. and Rao.
M. (1986) Mode of action and properties of xylanase and β-xyolsidase from Neurosporacrassa. Biotechnol. Bioeng.
28, 1832-1837.
7. Desphande, M. B. and Sadana, J. C. (1989) Purification and characterization of a cellulase-free endo-1,4-β-xylanase of Sclerotium rolfsii UV-8 mutant. Enzyme Microb. Technol.
11, 678-684.
8. Matsuo, M. and Yasui, T. (1988) Xylanase of Malbranchea pulchella var. sulfurea. Methods. Enzymol. 160, 671-674.
9. Mountfort, D. O. and Asher, R. A. (1989) Production of xylanase by the ruminal anaerobic fungus Neocallimastis frontalis. Appl. Environ. Microbiol. 55, 1016-1022.
10. Ghangas, G. V., Hu, Y.-J. and Wilson, D. B. (1989) Clon- ing of a Thermomonospora fusca xylanase gene and its Fig. 2. Effects of temperature and pH on the activity of
recombinant xylanase. Xylanase activity was assayed by the standard method as described in Materials and Methods. (A) Effects of temperature on the activity of xylanase were deter- mined between 60 to 110oC. (B) Effects of pH on the activity of xylanase were determined in a series of 50 mM each sodium citrate buffer (pH 3.0-6.0) and phosphate buffer (pH 6.0-8.0).
Fig. 3. Reaction activity of recombinant xylanase on oatspelts arabinoxylan and birchwood xylan. Each substrate (2.5%, w/v) was incubated at 90oC for 30 min with 0.85 U of purified recombinant xylanase in final volume of 200µl, and 3µl of each reaction mixtures was spotted onto a TLC plate and analyzed. Lane 1: G1-G7. Lane 2: xylose. Lane 3: the reac- tion product incubated with E. coli crude extract and oatspelts arabinoxylan. Lane 4: oatspelts arabinoxylan. Lane 5: the reac- tion product incubated with purified recombinant xylanase and oats spelts arabinoxylan. Lane 6: birchwood xylan. Lane 7: the reaction product incubated with purified recombinant xylanase and birchwood xylan.
160 Hyang-Sik Yoon et al.
expression in Escherichia coli and Streptomyces lividand. J.
Bacteriol. 171, 2963-2969.
11. Gilbert. H. J., Sullivan, D. A., Jenkins, G., Kellett, L. E., Minton, N. P. and Hall, J. (1988) Molecular cloning of mul- tiple xylanase genes from Pseudomonas fluorescens subsp.
cellulosa. J. Gen. Microbiol. 134, 3239-3247.
12. Kudo, T., Ohkoshl, A. and Horlkoshi, K. (1985) Molecular cloning and expression of a xylanase gene of alkalophilic Aeromonas sp. no. 212 in Escherichia coli. J. Gen. Micro- biol. 131, 2825-2830.
13. Yang, R. C. A., MacKenzie, R., Bilous, D. and Narang, S.
A. (1989) Hyperexpression of a Bacillus circulans xylanase gene in Escherichia coli and characterization of the gene product. Appl. Environ. Microbiol. 55, 1192-1195.
14. Gruninger, H. and Fiechter, A. (1986) A novel, highly ther- mostable D-xylanase. Enzyme Microb. Technol. 8, 309-314.
15. Tan, L. U. L., Mayers, P. and Saddler, J. N. (1987) Purifica- tion and characterization of a thermostable xylanase from a thermophile fungus Thermoascus aurantiacus. Can. J.
Microbil. 33, 689-692.
16. Sipat, A., Taylor, K. A., Lo, R. Y. C., Forsberg, C. W. and Krell, P. J. (1987) Molecular cloning of a xylanase gene from Bacteroides succinogenes and its expression in Escher- ichia coli. Appl. Environ. Microbiol. 53, 447-481.
17. Stetter, K. O., Fiala, G., Huber, G., Huber, R. and Segerer,
R. (1990) Hyperthermophilic microorganisms. FEMS Micro- biol. Rev. 75, 117-124.
18. Huber, R., Langworthy, T. A., Konig, H., Thomm, M., Woese, C. R., Sleytr, U. B. and Stetter, K. O. (1986) Ther- motoga maritima sp. nov. represents a new genes of unique extremely thermophilic eubacteria growing up to 90oC.
Arch. Microbiol. 144, 324-333.
19. Sambrook, J., Fritsch, E. F. and Maniatis, T. (1989) Molecu- lar Cloning: A Laboratory Manual, 2nd edn., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY.
20. Kim, Y. H., Kwon, T. K., Park, S. S., Seo, H. S., Cheong, J.-J., Kim, C. H., Kim, J.-K., Lee, J. S. and Choi, Y. D.
(2000) Trehalose synthesis by sequential reactions of recombinant maltooligosyltrehalose synthase and maltooli- gosyltrehalose trehalohydrolase from Brevibacterium hel- volum. Appl. Environ. Microbiol. 66, 4620-4624.
21. Bradford, M. M. (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 72, 248-254.
22. Winterhalter, C. and Liebl, W. (1995) Two Extremely Ther- mostable Xylanases of the Hyperthermophilic Bacterium Thermotoga maritima MSB8. Appl. Environ. Microbiol., 61, 1810-1815.