531
©The Korean Society of Food Science and Technology
콜라비 새싹 추출물과 무순 추출물의 항산화 및 지방세포 분화 억제 활성
이영준·김재환·오지원·신기해·이종석·조주현1·박진주2·임정호2·이옥환*
강원대학교 식품생명공학과, 1(주)휴럼 중양연구소, 2한국식품연구원
Antioxidant and Anti-adipogenic Effects of Kohlrabi and Radish Sprout Extracts
Young-Jun Lee, Jae-Hwan Kim, Ji-Won Oh, Gi-Hae Shin, Jong Seok Lee, Ju-Hyun Cho1, Jin-Ju Park2, Jeong-Ho Lim2, andOk-Hwan Lee*
Department of Food Science and Biotechnology, Kangwon National University
1Hurum Central Research Institute
2Korea Food Research Institute
Abstract Common cruciferous vegetables, kohlabi (Brassica oleracea) and radish (Raphanus sativus), contain phytochemicals such as glucosinolates and carotenoids. Therefore, this study investigated the antioxidant and anti- adipogenic effects of kohlrabi sprout extract (KSE) and radish sprout extract (RSE). The total carotenoid and glucosinolate contents of KSE and RSE were 39.50±0.67 and 76.73±2.75 mg/g, respectively. The total glucosinolate contents of KSE and RSE were 2.65±0.02 and 8.13±0.54 mg/g, respectively. The in vitro-antioxidative activities of KSE and RSE were significantly increased in a dose-dependent manner. Furthermore, β-carotene and glucosinolate-enriched KSE and RSE significantly inhibited lipid accumulation and reactive oxygen species production during the adipogenesis of 3T3-L1 preadipocytes. These results suggest that glucosinolate-enriched KSE and RSE, especially RSE, can be used in the treatment of obesity and as a natural source of antioxidants.
Keywords: kohlrabi, radish, sprout extract, glucosinolate, β-carotene
서 론
최근 경제의 발전과 더불어 삶의 질 향상으로 인해 전 세계적 으로 비만이 심각한 사회적 문제로 대두되었다. 비만은 우리가 음식 등을 섭취하여 체내로 들어오는 에너지가 운동 및 대사활 동으로 인해 소비되어지는 에너지보다 많게 될 경우 신체의 에 너지 불균형으로 인하여 지방이 축적되면서 발생하게 된다(1). 이 러한 비만은 고지혈증, 고혈압, 동맥경화 및 2형 당뇨병을 포함 한 대사증후군을 유발하는 질병으로 최근, 비만의 예방 및 치료 를 위한 연구가 활발히 진행되고 있다(2). 특히, 웰빙(well-being) 및 식품안전에 대한 관심이 높아짐에 따라 약물이나 합성된 첨 가물보다는 천연물 및 약용식물 등의 섭취를 통하여 이를 달성 하고자 다양한 생리활성을 가지는 천연물에 대한 검색 및 연구 가 활발히 이루어지고 있다(3).
콜라비(Brassica oleracea, Kohlarbi)와 무(Raphanus sativus, Radish)는 브로콜리, 양배추, 케일, 겨자 등과 함께 십자화과 (Cruciferae) 식물에 속한다. 십자화과 식물은 β-carotene, 루틴, 식이
섬유소 및 다양한 비타민류, 특히 비타민 C와 E를 풍부하게 함 유하는 것으로 알려져 있다. 또한, 십자화과 채소는 glucosinolate 성분이 함유되어 있으며, glucosinolate는 십자화과 식물의 대표적 인 2차대사 산물로서 sulfate를 포함하는 β-D-glucoside의 형태로 십자화과 채소의 종류에 따라 함유하는 glucosinolate의 종류 및 함량이 다른 것으로 알려져 있다(4). 예를 들어 Ciska 등(5)은 콜 라비에 함유되어 있는 glucosinolate는 glucoraphanin인 반면, 무의 경우 glucoraphenin이 주로 함유되어 있다고 보고하였다. 이러한 glucosinolate 및 그 분해산물은 예로부터 통풍, 설사 및 두통의 치료를 위해 이용되었다(6).
새싹 채소란 일반적으로 종자를 발아시킨 후 약 1주일 정도 된 채소의 싹을 말한다. 종자에는 싹을 틔우고 성체가 되기까지 열악한 환경을 극복하고 성장하기 위한 다양한 영양소가 포함되 어 있으며, 새싹이 돋아나는 시기에는 성장과 생명 유지를 위하 여 다량의 영양소를 합성하게 된다(7). 뿐만 아니라 새싹 채소는 생육기간이 짧아 농약을 사용하지 않기 때문에 농약에 대한 소 비자의 우려도 적은 편이어서 최근 새싹 채소에 대한 소비자의 관심도가 증가하고 있다(7). Fahey 등(8)의 연구결과에 따르면, 브 로콜리에 다량 함유되어 있는 glucosinolate의 한 종류인 sul- foraphane의 경우 성숙한 브로콜리보다 어린 브로콜리 새싹에 존 재하는 sulforaphane의 함유량이 약 40배 이상 높은 것으로 보고 하였다. 이와 같은 이유로 새싹 채소에 대한 소비자들의 관심은 높으나, 아직 새싹 채소에 대한 연구는 미비하며, 아직 초기단계 에 불과하다.
본 연구에서는 십자화과 채소 중 콜라비 새싹 및 무순 추출물
*Corresponding author: Ok-Hwan Lee, Department of Food Science and Biotechnology, Kangwon National University, Chuncheon, Gangwon 200-701, Korea
Tel: 82-33-250-6454 Fax: 82-33-259-5565
E-mail: [email protected]
Received January 7, 2014; revised February 18, 2014;
accepted February 19, 2014
내에 존재하는 carotenoid와 glucosinolate의 함량을 조사하였다. 또 한, 다양한 항산화 모델을 통하여 항산화 활성을 평가하였으며, 3T3-L1 지방세포의 분화시 콜라비 새싹 및 무순 추출물과 그들 의 지표성분인 β-carotene, glucoraphanin 및 glucoraphenin (Fig.
1)을 처리함으로써 분화 단계에서의 지방 축적 억제와 활성산소 생성 억제 활성을 평가하여, 추후 십자화과 새싹채소를 이용시 기초자료를 제공하고자 하였다.
재료 및 방법
추출물 제조 및 시약
본 연구에서 사용된 콜라비 새싹 추출물(kohlrabi sprout extract, KSE) 및 무순 추출물(radish sprout extract, RSE)은 한국식품연구 원으로부터 분양받은 것으로 제조과정은 다음과 같다. 콜라비 새 싹 및 무순의 무게에 각각 100배의에탄올(80%)을 가한 후, 상 온에서 약 24시간 교반하여 추출하였다. 추출액은 Whatman No. 1 여과지(Whatman Ltd., Maidstone, Kent, England)로 2회 여과 한 후 하루 동안 정치시킨 뒤 다시 한번 여과하였다. 그 후 회전식 진공 농축기(Eyela Co., Tokyo, Japan)를 사용하여 40oC에서 감압 농축 한 후, 동결건조(FD5510, Ilshin Lab. Co., Seoul, Korea) 하 여 추출물을 제조하였다.
실험에 사용된 시약인 3-isobutyl-1-methylxanthine (IBMX), Oil red O (ORO), dexamethasone (DEX), N-acetyl-L-cysteine (NAC), isopropanol, potassium ferricyanide, 1,1-diphenyl-2-picryl hydrazyl radical (DPPH), trichloroacetic acid (TCA), 2,4,6-tris(2-pyridyl)-s- triazine (TPTZ), potassium persulfate, insulin, iron chloride, 2,2'- azino-bis(3-ethylbenzothiazoline-6-sulfonic acid) diammonium salt (ABTS), β-carotene, nitrotetrazolium blue chloride (NBT), methyl tertiary butyl ether (MTBE), butylated hydroxytoluene (BHT), sinigrin hydrate 등은 sigma (Sigma-aldrich Co., St. Louis, MO, USA)로부터 구입하였고, Dulbecco’s Modified Eagle’s medium (DMEM), bovine serum (BS), fetal bovine serum (FBS), phosphate- buffered saline (PBS), penicillin-streptomycin (P/S), 및 trypsin-EDTA 는 Gibco (Gaithersburg, MD, USA)로부터 구입하였다. Gluc- oraphanin 및 glucoraphenin은 Cfm Oskar Tropitzsch (Marktred- witz, Germany)로부터 구매하였으며, 3T3-L1 지방세포는 American Type Culture Collection (ATCC, CL-173, Manassas, VA, USA) 으로부터 분양 받아 사용하였다.
Total carotenoid 함량 분석
새싹채소의 total carotenoid 함량은 Kim 등(9)의 방법을 이용하 여 측정하였다. 동결 건조된 시료 0.2 g에 0.1%의 BHT를 포함하 는 에탄올 6 mL을 혼합하여 85oC 드라이오븐에서 예열시켰다. 그 후, 80% KOH 120 µL를 첨가하여 다시 85oC 드라이오븐에서 10 min 동안 반응시킨 후 반응액이 충분히 냉각되면 증류수와 hexane 용액을 3 mL 첨가하고 3,500×g에서 15 min 동안 원심분 리하여 상층액을 취하였다. 상층액을 분리한 잔여물에 다시 증류 수와 hexane 용액을 각 3 mL씩 첨가하고, 동일한 과정을 1회 반 복하여 상층액이 총 2회 수집된 것을 질소 농축시킨 후, 잔여물 을 MeOH/MTBE (1:1)에 녹여 분석에 사용하였다. 분석은 450 nm에서 microplate reader (Molecular Devices, Sunnyvale, CA, USA)를 이용하여 흡광도를 측정하여 total carotenoid함량을 계산 하였다. 표준물질로는 β-carotene을 사용하였다.
Total glucosinolate 함량 분석
새싹채소의 total glucosinolate 함량은 Kestwal 등(10)의 방법을 응용하여 측정하였다. 동결 건조된 시료 0.1 g에 99% 에탄올 3 mL 을 넣고 30oC에서 100 rpm으로 20시간 동안 교반하여 용액을 제 조하였다. 그 후, 상층액을 여과지(Whatman, No. 1)에 여과시켜 남은 잔여물에 에탄올 3 mL를 넣고 교반을 한 뒤 다시 여과액과 합하여 실험에 이용하였다. 시액 20 µL와 2 mM sodium tetrachlo- ropalladate solution 300µL를 혼합하여 실온에서 30분간 정치시킨 후 450 nm에서 흡광도를 측정하여 함량을 구하였다. 이 때 표준 물질로는 sinigrin hydrate를 사용하였다.
DPPH radical 소거능 측정
DPPH radical 소거능은 수소 공여능을 측정하는 방법으로 비 교적 간단하고 짧은 시간 내 항산화 활성을 평가할 수 있기 때 문에 널리 이용되는 방법 중 하나이다(11). 이러한 DPPH radical 소거능은 Kim 등(12)의 방법을 변형하여 측정하였다. 0.4 mM DPPH 용액 0.8 mL과 시료를 각각 0.2 mL 첨가하여 vortex를 이 용하여 진탕하고 암소에서 10 min 동안 방치하였다. 그 후, micro- plate reader를 이용하여 517 nm에서 흡광도를 측정하였으며 이때 대조군으로 ascorbic acid (ASA)를 사용하였다. 시료의 DPPH radical 소거능은 다음 식에 의해 나타내었다.
DPPH radical 소거능(%)= ×100
ABTS radical 소거능 측정
ABTS radical 소거능은 potassium persulfate와 반응하여 생성된 ABTS·+이 항산화성 물질에 의해 제거되면서 청록색이 탈색되 어지는 것을 측정하는 방법이다(13). 활성의 측정은 Roberta 등 (14)의 방법을 응용하여 평가하였다. 2.6 mM의 potassium persul- phate와 7.4 mM의 ABTS를 각각 제조하여 섞은 후, 암소에 16시 간 동안 방치하여 양이온(ABTS+)을 형성시킨 후, 734 nm에서 흡 광도의 값이 0.70±0.02가 되도록 에탄올을 이용하여 희석하였다.
희석된 ABTS·+용액 1 mL에 시료 20 µL를 첨가한 뒤 6 min 후 흡광도를 측정하였다. 시료의 ABTS radical 소거능은 다음 식에 의해 나타내었다.
ABTS radical 소거능= ×100
Ferric ion reducing antioxidant power (FRAP) 활성 측정 FRAP 활성 측정은 전자 공여능을 측정하지 않고 산화 및 환 원 반응을 측정방법으로 항산화 물질이 노란색 계열의 Fe(III)- TPTZ를 푸른색 계열의 Fe(II)-TPTZ로 환원시키는 원리를 이용한 다(15). 시료의 FRAP 활성은 Benzie와 Stranin(16)의 방법을 변형 하여 측정하였다. Acetate buffer (pH 3.6, 300 mM):TPTZ (10 mM):
FeCl3·6H2O (20 mM)를 각각 제조하여 10:1:1의 비율로 섞어 FRAP용액을 제조하였다. 제조한 FRAP 용액 1.5 mL, 시료 50 µL 및 증류수 150 µL를 각각 혼합하여 10 min 동안 상온에서 정치 후, 590 nm에서 흡광도를 측정하였다.
환원력 측정
환원력의 측정은 Oyaizu(17)의 방법을 변형하여 측정하였다. 시 1 ATest
AControl ---
⎝ – ⎠
⎛ ⎞
1 ATest AControl ---
⎝ – ⎠
⎛ ⎞
료 1 mL과 sodium phosphate buffer (pH 6.6, 200 mM) 5 mL 및 1% potassium ferricyanide 5 mL을 각각 차례로 가하여 vortex를 이 용하여 진탕하고 50oC의 수욕 상에서 20 min 반응시킨 뒤, 10%
TCA 용액 5 mL 가하여 1,790×g에서 10min 동안 원심분리 하였다.
그 후 원심분리 된 용액의 상등액 2.5 mL을 취한 후 증류수 2.5 mL 및 iron chloride 0.5 mL을 가하여 700 nm에서 흡광도를 측정하였다.
세포독성 평가
3T3-L1 지방세포에 대한 시료의 세포 독성평가는 XTT(2,3-bis (2-methoxy-4-nitro-5-sulfophenyl)-2H-tetrazolium-5-carboxanilide innersalt) assay kit (WelGene, Seoul, Korea)를 이용하여 측정하였 다. 지방세포는 실험 전날 1×106 cell 농도로 96-well plate에 seed- ing을 하고 시료를 2 µL를 처리하여 24시간 동안 배양하였다.
40µL의 XTT working solution을 각각의 well에 첨가하여 혼합하 였다. 그 후 96-well plate를 CO2 incubator에서 4시간 동안 배양 한 후, 450 nm 흡광도 값에서 690 nm의 흡광도 값을 뺀 결과 값 을 계산하였다(18).
3T3-L1 세포 배양 및 분화
3T3-L1 전지방세포를 24-well plate에 각각 1×106으로 seeding한 후, BS (10%) 및 P/S (1%)를 함유한 DMEM (89%)에서 cell이 100% confluence가 될 때까지 배양하였다. 그로부터 2일 후, 지 방세포의 분화유도 물질인 IBMX (0.5 mM), dexamethason (1 µM) 및 insulin (10 µg/mL), FBS (10%) 및 P/S (1%)를 함유한 medium 으로 전지방세포를 지방세포로 분화유도 하였다. 2일 마다 insulin (10µg/mL), FBS(10%) 및 P/S(1%)가 함유된 medium으로 교체하 여주었다. 이 때 시료의 효과를 관찰하기 위하여 각각의 시료를 처리하여 주었으며, positive control(양성 대조군)으로 항산화제인 NAC (10 mM 1.63 mg/mL)를 처리하였다(17).
ORO staining
지방세포의 분화과정에서 세포 내 지방 축적량을 측정하고자 ORO 시약을 이용하여 염색하였다. Medium을 제거한 후, 10%
formalin 용액 0.5 mL를 첨가하여 5분간 실온에서 방치한 뒤 제 거하였다. 그 후 동량의 formalin 용액을 세포에 처리하여 1시간 동안 실온에서 정치한 후, formalin을 제거하고 60% isopropanol 용액으로 세척하여 세포를 완전히 건조시켰다. 완전히 건조된 세 포들은 미리 제조해 둔 ORO working solution (ORO:DDW=6:4) 으로 염색 한 후, 증류수로 3-4회 세척 후 완전히 건조시켰다. 세 포 내 축적된 지방 성분과 결합한 ORO는 100% isopropanol을 이 용하여 모두 용출시킨 후, microplate reader를 이용하여 490 nm 에서 흡광도를 측정하였다(19).
NBT assay
지방세포 내 reactive oxygen species (ROS) 생성량을 측정하기 위해 분화된 지방세포에 PBS를 이용하여 2회 세척 후, 0.2% NBT 용액 0.2 mL를 첨가하여 CO2 incubator에서 90 min 동안 반응시 켰다. 염색된 cell을 증류수로 2회 세척 후 완전히 건조시킨 후, KOH solution (1 N KOH:DMSO=7:3)을 이용하여 dark blue for- mazan을 모두 용출시킨 후 570 nm에서 흡광도를 측정하였다(20).
통계분석
실험결과는 SAS package (release 9.3)를 이용하여 one-way ANOVA 분석 수행하였고 평균값의 통계적 유의성은 p<0.05 수 준에서 검정하였다.
결과 및 고찰
콜라비 새싹 및 무순 추출물의 total carotenoid 및 glucosi- nolate 함량 비교
십자화과 식물에는 다양한 종류의 carotenoid와 glucosinolate 성 분을 함유하는 것으로 알려져 있으며(4), 이러한 식물의 2차 대 사산물들은 항산화, 항비만, 항염증 등의 다양한 생리활성을 갖 는다(21). 따라서 본 연구에서는 콜라비 새싹 추출물(KSE) 및 무 순(RSE) 추출물 내에 존재하는 total carotenoid 및 glucosinolate 함량을 분석하였다. 측정 결과(Table 1), KSE의 total carotenoid 및 glucosinolate의 함량은 각각 39.50±0.67 및 2.65±0.02 mg/g으 로 나타났다. RSE의 경우, total carotenoid 및 glucosinolate의 함 량이 각각 76.73±2.75 및 8.13±0.54 mg/g으로 나타났으며, RSE의 경우 KSE보다 total carotenoid 및 glucosinolate의 함량이 높게 나 타났다. Ciska 등(5)의 연구결과에서도 무순의 total glucosinolate 함량(126.42±26.20 mg/100 g)이 콜라비의 total glucosinolate 함량 (28.46±6.76 mg/100 g) 보다 높은 것으로 나타나 본 실험의 결과 와 유사한 경향을 나타냈다.
콜라비 새싹 추출물 및 무순 추출물의 항산화 효과
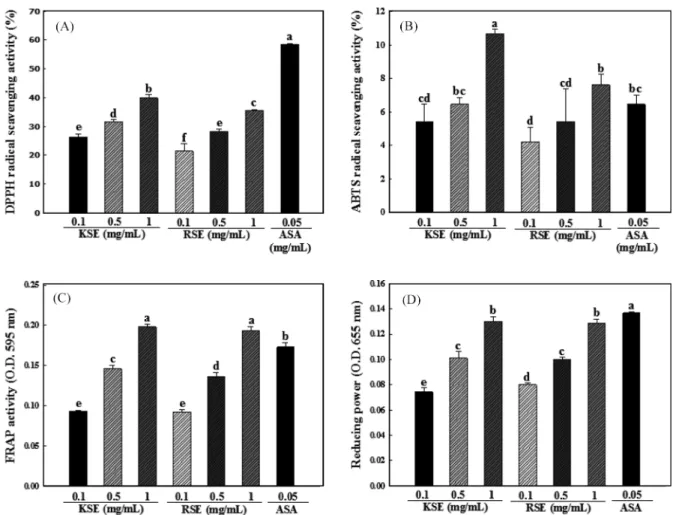
다양한 in vitro 항산화 실험을 이용하여 KSE 및 RSE의 항산 화 활성을 평가하였다. DPPH assay의 경우 가장 간단한 항산화 활성 평가법이지만, DPPH 시약이 유기 용매에 녹기 때문에 친 수성 물질을 측정 할 경우 제약이 따르며(13), 517 nm에서 흡광 도를 측정하기 때문에 평가시 시료의 색에 의해 영향을 받을 수 있다(22). 또 Huang 등(23)의 연구에 따르면 DPPH assay는 eugenol 및 그와 유사한 구조를 가지는 페놀 성분을 측정할 경우, 항산화 활성이 반대로 나오기도 한다. ABTS assay는 친수성 및 소수성 물질의 항산화 활성 평가가 가능하며, 넓은 pH범위를 가 지며 이온 세기에 영향을 받지 않기 때문에 식품의 항산화 활성 을 평가할 때 많이 사용된다(13). 그러나 ABTS assay의 경우 end- point assay로, 항산화 물질의 농도 및 반응시간 등이 고려되어야 한다(24). FRAP assay는 분석이 간단하며 신속하다는 장점을 갖 는다. 그러나 글루타치온이나 단백질과 같이 -SH (thiols)기를 포 함하는 특정 성분의 경우 항산화 활성을 측정하지 못한다는 단 점이 있다(25). 따라서 본 연구에서는 4가지 항산화 실험 (DPPH radical scavenging, ABTS radical scavenging, FRAP, reducing power assay)을 이용하여 다양한 농도에서 콜라비 새싹 추출물 (KSE) 및 무순 추출물 (RSE)의 항산화 활성을 평가하였으며 ascorbic acid (ASA)를 양성대조군으로 이용하였다.
4종의 항산화 실험을 이용하여 KSE 및 RSE의 항산화 활성을 측정한 결과는 Fig. 2와 같다. DPPH 및 ABTS assay의 경우 KSE와 RSE 모두 농도 의존적으로 항산화 활성이 증가하였으며, KSE의 항산화 활성이 RSE의 항산화 활성보다 유의적으로 높게 나타났다(p<0.05). 반면 FRAP 활성 및 reducing power의 경우 KSE 및 RSE 모두 농도 의존적으로 상승하였지만, DPPH 및 ABTS assay와 달리 KSE 와 RSE간의 항산화 활성은 유의적인 차이를 보이지 않았다. 또, 양성대조군으로 사용한 ASA 또한 Table 1. Total carotenoid and glucosinolate contents of KSE and RSE
KSE RSE
Total carotenoid (mg/g) 39.50±0.67 76.73±2.75 Total glucosinolate (mg/g) 02.65±0.02 08.13±0.54
DPPH assay의 경우 시료보다 높은 활성을 보인 반면, ABTS 및 FRAP assay에서는 시료보다 낮은 활성을 보였다. 이러한 결과는 앞에서 기술하였듯이 항산화 실험마다 반응 메커니즘이 다르며, 특정 성분에 대하여 반응정도가 다르기 때문으로 사료된다. 양배 추 등 13종의 야채 총 927개의 시료를 이용해서 oxygen radical absorbance capacity (ORAC) value와 FRAP 활성을 측정한 Ou 등(25)의 연구에 따르면 ORAC value는 peroxyl radical 소거능을
확인하고, FRAP 활성은 Fe(III) 억제 효과를 나타내는 것으로 그 원리가 서로 다르므로 각 시료간의 경향이 실험마다 다르다고 발 표하였다. 이와 같은 이유로 최근 많은 연구에서는 시료의 항산 화 활성을 평가 할 경우, 한 가지 방법으로 평가하는 것은 옳지 못하며, 다양한 방법을 이용하여 항산화 활성을 평가하기를 권장 하고 있다.
Fig. 1. Structure of β-carotene (A), glucoraphanin (B), and glucoraphenin (C).
Fig. 2. DPPH radical scavenging activity (A), ABTS radical scavenging activity (B), FRAP activity (C) and reducing power (D) of KSE and RSE at various concentration. Each values are the means±SD of samples. Bars with different letters indicate statistically significant differences among groups at p<0.05 by one-way ANOVA.
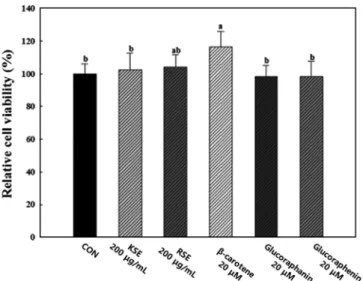
콜라비 새싹 및 무순 추출물의 세포 독성 평가
3T3-L1 지방세포에 대한 시료의 세포독성 결과는 Fig. 3과 같 다. 추출물(KSE 및 RSE) 및 지표성분(β-carotene, glucoraphanin 및 glucoraphenin)을 각각 200 µg/mL 및 20 µM의 농도로 세포에 처리하였을 때, 세포 내 독성을 보이지 않았으며 현미경 상에서 morphology의 변화도 관찰되지 않았다. 따라서 본 연구에서 추출 물과 지표성분을 각각 200 µg/mL 및 20 µM 농도로 지방세포에 처리하여 지방세포 분화억제 및 ROS 생성 저감 효능평가를 실 시하였다.
콜라비 새싹 및 무순 추출물의 지방축적 억제 및 ROS 생성 억제 활성
지방세포의 분화시 KSE 및 RSE 처리에 따른 지방세포 분화 억제 활성을 측정하기 위하여 ORO 염색을 통해 지방세포 내 생 성된 지방축적량을 평가하였다(Fig. 4A). KSE 및 RSE를 처리한 군의 경우 아무것도 처리하지 않은 음성대조군(CON)에 비해 지 방축적량이 각각 80.09±2.28%, 65.42±2.66%로 유의적인 감소하 였다. 양성대조군으로 사용한 NAC 처리군의 경우, 대조군에 비 해 지방축적을 유의적으로 감소시키는 것으로 나타났다. NAC는 항산화제로 지방세포 분화과정에 처리하였을 경우, 분화관련 전 사인자들의 발현을 억제하여 지방세포 내 중성지방의 축적을 억 제하는 것으로 알려져 있다(26).
한편, 지방세포의 분화 및 지방의 축적은 세포 내 산화적 스 트레스와 밀접한 관계를 갖는 것으로 알려져 있다(18,27). 3T3-L1 전지방세포는 지방세포로 분화하는 과정에서 세포 내로 유입되 는 포도당을 저장하기 위하여 지방 합성과 관련된 pentose phos- phate pathway (PPP) 작동함에 따라 nicotinamide adenine dinu- cleotide phosphate (NADPH)가 생성된다. 이렇게 생성된 NADPH 는 NADPH oxidase 4 (NOX4)에 의하여 NADP+로 전환되어지는 데, 이 때 세포 내 다량의 superoxide를 생성되며, 지방세포에서 과도하게 생성된 산화적 스트레스는 지방세포의 분화를 촉진시 키며 macrophage를 자극하여 또 다른 산화적 스트레스를 생성하 기 때문에 비만의 주요 원인 중에 하나라고 보고되고 있다(28).
따라서 본 연구에서 NBT assay를 이용하여 지방세포의 분 화과정에서 KSE 및 RSE 처리에 따른 ROS 생성 억제 활성을 평 가하였다. KSE 및 RSE를 처리한 군의 경우 음성대조군(CON)에 비해 ROS 생성량이 각각 69.67±8.11 및 63.03±6.00%로 유의적인 감소하는 경향을 나타내었다(Fig. 4B).
이상의 결과를 통해 지방세포의 분화과정에서 KSE 및 RSE의 처리는 지방세포의 분화 및 지방축적을 억제하였고 세포 내 ROS 생성을 저해하므로, 지방세포의 분화가 촉진되는 것을 억제하는 Fig. 3. Effects of KSE, RSE, β-carotene, glucoraphanin, and
glucoraphenin on the cell viability. Cell viability was determined using the XTT assay. Each value is the mean±SD of the results from five different plates (n=5). Statistical analysis was performed using the one-way ANOVA (p<0.05).
Fig. 4. Effect of the KSE and RSE on lipid accumulation (A) and reactive oxygen species (ROS) production (B) in 3T3-L1 cells during the adipogenesis. CON, control cells which were differentiated with MDI; NAC, positive control cells which were differentiated with MDI in the presence of NAC. All values are presented as the means±SD. Bars with different letters indicate statistically significant differences among groups at p<0.05 by one-way ANOVA.
것으로 사료된다. 이는 콜라비 추출물을 갓 태어난 돼지에서 분 리한 지방전구세포에 처리한 후 지방세포의 분화억제를 측정한 Song 등(29)의 연구에서 콜라비 추출물이 지방전구세포의 분화를 억제한 결과와 일치한다.
β-Carotene, glucoraphanin 및 glucoraphenin의 지방축적 억제 및 ROS 생성 억제 활성
KSE 및 RSE의 주요성분으로 알려진 β-carotene, glucoraphanin 및 glucoraphenin의 지방 축적 및 ROS 생성 억제 활성을 평가하 였다. 지방세포의 분화시 β-carotene, glucoraphanin 및 glucoraphe- nin을 처리한 결과, 음성대조군(CON)에 비하여 지방 축적량이 각 각 31.28±4.70, 16.23±4.00 및 20.72±4.01%으로 감소하였다 (p<0.05). 특히, glucosinolate인 glucoraphanin 및 glucoraphenin의 경우 β-carotene보다 높은 지방 축적 억제 활성을 보였다 (Fig.
5A). NBT assay를 통하여 ROS 생성 억제 활성을 측정한 결과, β-carotene, glucoraphanin 및 glucoraphenin를 처리한 군의 경우 ROS 생성량이 각각 43.71±4.51, 30.01±3.04 및 34.73±5.36%으로 유의적으로 감소하였다(Fig. 5B).
β-Carotene, glucoraphanin 및 glucoraphenin은 지방 세포의 분화 시 지방 축적 억제 및 ROS 생성 억제 활성을 갖는다. Kameji 등 (30)의 연구에서 β-carotene이 축적된 3T3-L1 지방세포가 대조군 에 비해 인슐린감수성과 ROS 저해 관련 유전자의 발현을 감소 시킨다는 결과와 유사하다. 이와 같은 이유로 인하여 total glu- cosinolate 및 carotenoid 함량이 높은 RSE가 KSE보다 높은 지방 축적 억제 활성을 보인 것으로 사료된다.
요 약
십자화과 채소인 콜라비 및 무에는 glucosinolate 성분이 함유 되어 있으며, 특히 새싹 채소의 경우 성숙한 채소보다 glucosinolate 의 함량이 더 높은 것으로 알려져 있다. 이와 같은 이유로 최근 새싹채소에 대한 관심은 높으나, 아직 새싹채소에 대한 연구는 미비한 편이다. 따라서 본 연구에서는 십자화과 채소인 콜라비 새싹 추출물(KSE) 및 무순 추출물(RSE)의 total carotenoid 및
glucosinolate의 함량을 분석하였고 이들 추출물의 항산화 및 anti- adipogenic 활성을 평가하였다. KSE의 total carotenoid 및 glucos- inolate 함량은 각각 39.50±0.67 및 2.65±0.02 mg/g이며, RSE의 total carotenoid 및 glucosinolate 함량은 각각 76.73±2.75 및 8.13
±0.54 mg/g으로 KSE보다 높았다. 다양한 항산화 평가 모델(DPPH, ABTS, FRAP, reducing power)을 통하여 항산화 활성을 측정한 결과, KSE 및 RSE는 농도 의존적으로 항산화 활성이 증가하는 경향을 나타내었다. 또한, 지방세포의 분화과정에서 KSE 및 RSE 의 처리는 지방세포의 지방축적 및 ROS 생성을 억제하였으며, KSE 및 RSE의 주요 성분으로 알려진 β-carotene, glucoraphanin 및 glucoraphenin의 처리는 지방 축적 억제 및 ROS 생성 저감 활 성을 나타내었다. 이상의 결과로부터, β-carotene, glucoraphanin 및 glucoraphenin이 풍부한 RSE 및 KSE는 항산화 활성 및 지방세포 분화억제 효능을 갖으며, 천연물 유래 항비만 소재로서 활용 가 능성이 높은 것으로 기대된다.
감사의 글
본 연구는 지식경제부 광역경제권 선도산업 육성사업과제(과제 번호:C1009939-01-01) 및 한국식품연구원 위탁연구 개발과제(과 제번호:C1008688-01-02)로 수행한 연구의 일부로 이에 감사드립 니다.
References
1. Dewulf EM, Cani PD, Neyrinck AM, Possemiers S, Van Holle A, Muccioli GG, Deldicque L, Bindels LB, Pachikian BD, Sohet FM, Mignolet E, Francaux M, Larondelle Y, Delzenne NM. Inu- lin-type fructans with prebiotic properties counteract GPR43 over- expression and PPARγ-related adipogenesis in the white adipose tissue of high-fat diet-fed mice. J. Nutr. Biochem. 22: 712-722 (2011)
2. Kopelman PG. Obesity as a medical problem. Nature 404: 635- 643 (2000)
3. Lee YJ, Yoon BR, Kim DB, Kim MD, Lee DW, Kim JK, Lee OH. Antioxidant activity of fermented wild grass extracts. Korean J. Food Nutr. 25: 407-412 (2012)
Fig. 5. Effect of the β-carotene, glucoraphanin, and glucoraphenin on lipid accumulation (A) and reactive oxygen species (ROS) production (B) in 3T3-L1 cells during the adipogenesis. CON, control cells which were differentiated with MDI; NAC, positive control cells which were differentiated with MDI in the presence of NAC. All values are presented as the means±SD. Bars with different letters indicate statistically significant differences among groups at p<0.05 by one-way ANOVA.
4. Hwang ES. Changes in glucosinolate component content in urine after ingestion of fresh and cooked broccoli. Korean J. Food Cook. Sci. 26: 804-810 (2010)
5. Ciska E, Martyniak-Przybyszewska B, Kozlowska H. Content of glucosinolates in cruciferous vegetables grown at the same site for two years under different climatic conditions. J. Agr. Food Chem. 48: 2862-2867 (2000)
6. Kim YK, Kim GH. Changes in 3-butenyl isohiocyanate and total glucosinolates of seeds and young seedlings during growth of Korean Chinese cabbages. Korean J. Food Preserv. 10: 365-369 (2003)
7. Lee JW, Lee DY, Cho JG, Baek NI, Lee YH. Isolation and iden- tification of sterol compounds from the red kohlrabi (Brassica oleracea var. gongylodes) Sprouts. J. Appl. Biol. Chem. 54: 207- 211 (2010)
8. Fahey JW, Haristoy X, Dolan PM, Kensler TW, Scholtus I, Stephenson KK, Talalay P, Lozniewski A. Sulforaphane inhibits extracellular, intracellular, and antibiotic resistant strains of Heli- cobacter pylori and prevents benzo[a]pyrene-induced stomach tumors. Proc. Natl. Acad. Sci. 99: 7610-7615 (2002)
9. Kim HJ, Fonseca JM, Choi JH, Kubota C. Effect of methyl jas- monate on phenolic compounds and carotenoids of romaine let- tuce (Lactuca sativa L.). J. Agr. Food Chem. 55: 10366-10372 (2007)
10. Kestwal RM, Lin JC, Bagal-Kestwa D, Chiang BH. Glucosino- lates fortification of cruciferous sprouts by sulphur supplementa- tion during cultivation to enhance anti-cancer activity. Food Chem. 126: 1164-1171 (2011)
11. Koleva II, van Beek TA, Linssen JP, de Groot A, Evstatieva LN.
Screening of plant extracts for antioxidant activity: a comparative study on three testing methods. Phytochem. Anal. 13: 8-17 (2002)
12. Kim JH, Park JH, Park SD, Choi SY, Seong JH, Hoon KD. Prep- aration and antioxidant activity of health drink with extract pow- ders from Safflower (Carthamus tinctorius L.) seed. Korean J.
Food Sci. Technol. 34: 617-624 (2002)
13. Arnao MB. Some methodological problems in the determination of antioxidant activity using chromogen radicals: a practical case.
Trends Food Sci. Tech. 11: 419-421 (2000)
14. Re R, Pellegrini N, Proteggente A, Pannala A, Yang M, Rice- Evans C. Antioxidant activity applying an improved ABTS radi- cal cation decolorization assay. Free Radic. Biol. Med. 26: 1231- 1237 (1999)
15. Jeong JW, Lee YC, Jung SW, Lee KM. Flavor components of citron juice as affected by the extraction method. Korean J. Food Sci. Technol. 26: 709-712 (1994)
16. Benzie I, Strain J. The ferric reducing ability of plasma(FRAP) as a measure of antioxidant power: the FRAP assay. Anal. Bio- chem. 239: 70-76 (1996)
17. Oyaizu M. Studies on products of the browning reaction. Antioxi- dative activities of products browning reaction prepared from glu- cosamine Jpn. J. Nutr. Diet. 44: 307-315 (1986)
18. Furukawa S, Fujita T, Shimabukuro M, Iwaki M, Yamada Y, Nakajima Y, Nakayama O, Makishima M, Matsuda M, Shimo- mura I. Increased oxidative stress in obesity and its impact on metabolic syndrome. J. Clin. Invest. 114: 1752-1761 (2004) 19. Lee OH, Seo MJ, Choi HS, Lee BY. Pycnogenolinhibits lipid
accumulation in 3T3-L1 adipocytes with the modulation of reac- tive oxygen species (ROS) production associated with antioxidant enzyme responses. Phytother. Res. 26: 403-411 (2012)
20. Lee YJ, Yoon BR, Choi HS, Lee BY, Lee OH. Effect of Sargas- sum micracanthum extract on lipid accumulation and reactive oxygen species (ROS) production during differentiation of 3T3- L1 preadipocytes. Korean J. Food Preserv. 19: 455-461 (2012) 21. Larson RA. The antioxidants of higher plants. Phytochem. 27:
969-978 (1988)
22. Prior RL, Wu X, Schaich K. Standardized methods for the deter- mination of antioxidant capacity and phenolics in foods and dietary supplements. J. Agr. Food Chem. 53: 4290-4302 (2005) 23. Huang D, Ou B, Prior RL. The chemistry behind antioxidant
capacity assays. J. Agr. Food Chem. 53: 1841-1856 (2005)
24. Krishnaiah D, Sarbatly R, Nithyanandam R. A review of the anti- oxidant potential of medicinal plant species. Food Bioprod. Pro- cess. 89: 217-233 (2011)
25. Ou B, Huang D, Hampsch-Woodill M, Flanagan JA, Deemer EK.
Analysis of antioxidant activities of common vegetables employ- ing oxygen radical absorbance capacity (ORAC) and ferric reduc- ing antioxidant power (FRAP) assays: a comparative study. J.
Agr. Food Chem. 50: 3122-3128 (2002)
26. Blumberg JM, Tzameli I, Astapova I, Lam FS, Flier JS, Hollen- berg AN. Complex role of the vitamin D receptor and its ligand in adipogenesis in 3T3-L1 cells. J. Biol. Chem. 28: 11205-11213 (2006)
27. Lee OH, Kwon YI, Hong HD, Park CS, Lee BY, Kim YC. Pro- duction of reactive oxygen species and changes in antioxidant enzyme activites during differentiation of 3T3-L1 adipocyte. J.
Korean Soc. Appl. Biol. Chem. 52: 70-75 (2009)
28. Lee YJ, Han OT, Choi HS, Lee BY, Lee OH. Antioxidant and anti-adipogenic effects of PineXol. Korean J. Food Sci. Technol.
45: 97-103 (2013)
29. Song MY, Lee JJ, Cha SS, Chung CS. Effects of kohlrabi (Bras- sica oleracea var. Gongylodes) on proliferation and differentiation of pig preadipocytes and 3T3-L1 cells. KSAST. 55: 19-23 (2013) 30. Kameji H, Mochizuki K, Miyoshi N, Goda T. β-Carotene accu-
mulation in 3T3-L1 adipocytes inhibits the elevation of reactive oxygen species and the suppression of genes related to insulin sensitivity induced by tumor necrosis factor-α. Nutrition 26:
1151-1156 (2010)