Introduction
The Zostera marina bed is very important as rich, productive nursery and feeding area for juvenile and adult fishes (Adams, 1976; Kikuchi, 1966; Pollard, 1984; Klumpp et al., 1989; Kwak, 1997). The Z. marina bed of Jindong Bay, Korea, provide a habitat for variety of invertebrates and small fish, which in turn are the potential food of significant fisheries.
Stephanolepis cirrhifer (family Monocanthidae) is widely distributed in close association with Z.
marina bed along coastline of Korea and harvest- ed commercially due to their economic importance (Huh and Kwak, 1997d, Kim and Kang, 1993).
Despite their commercial value and abundance in a Z. marina bed, very little has been published on the feeding ecology of S. cirrhifer in Korean seagrass bed. To date of the study of S. cirrhifer have concentrated on physiological response and seasonal variation of abundance in the coastal fish assemblages (Yang, 1980; Chang, 1991; Kim et al., 1994; Huh and Kwak, 1997d). Worldwide, the family Monocanthidae was among the com- mon fish group in seagrass beds and feeding
habits of monocanthid species have been reported by several workers (Randall, 1967; Kikuchi, 1974;
Last, 1975; Bell et al., 1978; Conacher et al., 1979). These studies indicated that the diets of monocanthid fish species are extremely diverse and opportunistic feeders.
The aim of this study was therefore to investi- gate feeding habits of S. cirrhifer in a Z. marina bed. Specifically, the objectives are to determine the overall diet of this species and variations in diet in relation to fish size.
Materials and Methods
All the sampling was carried out over eelgrass (Zostera marina) bed in Jindong Bay, Korea (Fig.
1). Z. marina is forming subtidal bands (500~
700 m wide) in the shallow water (⁄3 m) along the shoreline of Jindong Bay.
Stephanolepis cirrhifer were collected with 5 m otter trawl (1.9 cm mesh wing and body, 0.6 cm mesh liner) throughout 2002. Stomachs from fish samples were preserved immediately in 10%
formaline and the length and weight of each fish recorded. Stomach contents were removed after 2 days and transferred to 70% isopropanol for stor-
─
─ 219 ──
Feeding Habits of Stephanolepis cirrhifer in a Zostera marina Bed
Seok Nam Kwak, Gun Wook Baeck and Sung-Hoi Huh*
Korea Inter-university Institutes of Ocean Science,
*Department of Oceanography, Pukyong National University, Pusan 608-737, Korea
Feeding Habits of Stephanolepis cirrhifer collected from a Zostera marina bed in Jindong Bay, Korea, were studied. S. cirrhifer is an omnivore which consumes mainly amphipods (gammarids and caprellids) and eelgrass, Z. marina. Its diet also includes a small amount of copepods, polychaetes, and isopods. S. cirrhifer showed ontoge- netic changes in feeding habits. Small individuals less than 2 cm SL fed mainly on copepods, however, amphipods were heavily selected with increasing fish size. The consumption of eelgrass by S. cirrhifer was 10~20% at all size classes. The dietary breadth of each size class showed a relatively low value.
Key words : Stephanolepis cirrhifer, feeding habits, Zostera marina bed, stomach contents, amphipods, eelgrass
age. Gut contents from each fish were identified as accurately as possible and the (1) occurrence, (2) number of individuals and (3) dry weight of each prey species recorded.
Dietary breadth index was calculated using Levins standardized index (Krebs 1989):
B
i= 1/n-1 (1/ ∑
jP
ij2-1)
Where B
i= Levins standardized index for preda- tor i, P
ij= proportion of diet of predator i that is made up of prey j, and n = number of prey cate- gories. This index range from 0 to 1, with low values indicating diets dominated by a few prey items (specialist predators) and higher values indicating generalist diets (Gibson and Ezzi, 1987; Krebs, 1989).
Results and Discussion
Size distribution
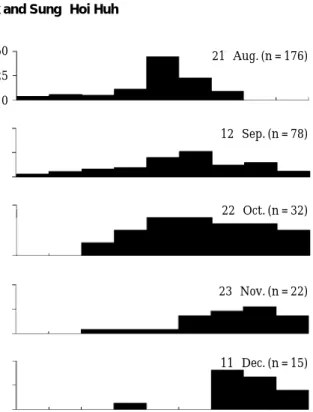
S. cirrhifer were present in a Z. marina bed from August to December 2002 (Fig. 2). Size range was from 1.4 cm to 9.6 cm SL during study period. S. cirrhifer first appeared in August at a mean length class of 5~7 cm SL and inhabited in the study area until December when occurred in size ranged from 4~10 cm SL. Number of indi- viduals peaked at 176 individuals in August and then declined sharply in the remaining periods.
Stomach contents analysis
A total of 291 S. cirrhifer stomachs were exam- ined, of which 22 (7.6%) were empty. The stom-
achs contained 16 identifiable prey components (Table 1). Gammarid amphipods were major prey groups for S. cirrhifer, comprising 47.6% of the diet by weight, 39.7% of the diet by number and occurring in 68.5% of all stomachs examined.
Ampelisca sp., Leucothoe sp. and Ericthonius sp.
were the principal gammarid amphipods items consumed. After gammarid amphipods, caprellid amphipods were important prey groups, making up 27.8% of the diet by weight, 37.2% of the diet by number and 61.4% of the diet by occurrence.
The caprellid amphipods Caprella kroeyeri, C.
scaura were the principal prey items. Eelgrass, Z. marina (16.4% of the diet by weight and 15.5%
of diet by occurrence) was the next largest dietary components. Copepods were next prey group, accounting for 5.3% of the diet by weight. Para- calanus parvus was the principal copepods items fed. Polychaetes and isopods were of little impor- tance and constituted 1.7%, 1.2% of the diet by weight. Crab larvae and algae composed a negli- gible portion of the diet.
Variations in stomach contents in relation to fish size
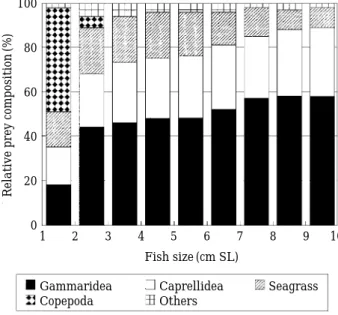
As can be seen in Fig. 3, smaller S. cirrhifer (⁄2
220 Seok Nam Kwak, Gun Wook Baeck and Sung-Hoi Huh
Fig. 1. Location of the study area (the black area).
Fig. 2. Monthly variation in size distribution of Stephano- lepis cirrhifer.
1 2 3 4 5 6 7 8 9 10
Fish size (cm SL) Dagu
35�06
N
Judo
Jindong bay
0 500 m
50 25 0
21-Aug. (n = 176)
12-Sep. (n = 78)
22-Oct. (n = 32)
23-Nov. (n = 22)
11-Dec. (n = 15)
Relative frequency(%)
cm SL) fed mainly copepods. Some gammarid amphipods, caprellid amphipods and eelgrass were also found. Studies of feeding habits of fish species in a Z. marina bed in Kwangyang Bay, copepods were mainly consumed by smaller Leiog- nathus nuchalis, Pholis nebulosa and Syngna- thus schlegei (Huh and Kwak, 1997a, b, c). This indicated that copepods were the most important prey for juvenile fish species in a Z. marina bed.
The portion of the diet attributable to copepods decreased gradually with increasing size, how- ever, proportions of gammarid amphipods and caprellid amphipods increased. On the other hand, the consumption of eelgrass by S. cirrhifer was 10.1~20.3% in all size classes. This onto- genetic change of feeding habits observed in the study area is generally similar to that of S. cir- rhifer in a Z. marina bed in Kwangyang Bay.
Juvenile S. cirrhifer fed on copepods, whereas the larger individuals ingest mainly amphipods (Kwak, 1997). Compared with feeding habits of fish species (family Monocanthidae) in the sea- grass beds worldwide, most of monocanthid spe-
cies undergoes a similar type of size-related changes regardless of location and climate, how- ever, relative percentage of main prey organisms consumed was difference among these species.
For example, Monacanthus chinensis (⁄10 cm SL) in Posidonia australis beds in Quibary, New South Wales, Australia, fed mainly gammarid amphipods, whereas larger individuals (¤10 cm SL) consumed mainly seagrass and gammarid amphipods (Conacher et al., 1979; Bell et al., 1978) and Last (1975) studied amphipods and algae were mainly consumed by larger size of Meuschenia australis, Meuschenia freycineti.
However, Randall (1967) recorded seagrass as a major food item (67%) for the West Indian mono- canthid species and small crustacean adhering to Z. marina were major prey group for monocan- thid fish in Japan (Kikuchi, 1974). This is proba- bly due that monocanthid species have special- ized mode of feeding on encrusting organisms with small mouth, their cryptic coloration and secretive behavior (Bell et al., 1978; Conacher et al., 1979; Kwak, personal observation). Most of monocanthid species use specialized teeth to bite off heavily small crustaceans and pieces of sea- grass and after digesting the encrusting organ- isms, the seagrass pieces are apparently expelled undigested (Bell et al., 1978). Conacher (1977), in experiments of
14C-labelled Posidonia was fed to monocanthid species, found that a considerable amount of the labile carbon in the seagrass was
Table 1. Percent composition of the stomach contents of Stephanolepis cirrhifer by frequency of occur- rence, number and dry weight
Prey organisms Occurrence Number Dry weight
(%) (%) (%)
Crustacea Amphipoda
Gammaridea 68.5 39.7 47.6
Ampithoe sp. 35.3 7.8 9.8
Leucothoe sp. 29.9 7.5 9.2
Ericthonius sp. 26.7 7.1 8.1
Elasmopus sp. 25.8 5.9 7.3
Unidentified 32.5 11.4 13.2
Caprellidea 61.4 37.2 27.8
Caprella kroeyeri 32.6 13.5 10.1
Caprella scaura 30.4 13.3 10.0
Caprella tsugarensis 29.8 10.4 7.7
Copepoda 23.3 20.3 5.3
Paracalanus parvus 19.5 8.1 4.5
Acartia omorii 18.8 7.7 0.4
Pseudodiaptomus marinus 15.5 4.5 0.4 Isopoda
Cymodoce japonica 6.3 1.4 1.2
Brachyura
Crab larvae 7.1 1.4 0.2
Polychaeta 1.2 0.3 1.7
Seagrass
Zostera marina 15.5 16.4
Algae 3.4 0.1
Total 100 100
Fig. 3. Relationships between relative prey composition (DW, %) and body length of Stephanolepis cirrhifer.
100
80
60
40
20
0
1 2 3 4 5 6 7 8 9 10
Fish size (cm SL)
Gammaridea Caprellidea Seagrass
Copepoda Others
Relative prey composition(%)
incorporated in the tissues of these fish. Thus the seagrass portion of the diet mainly represent an animal food type (e.g. amphipods, isopods etc.) and the diet is not herbivorous.
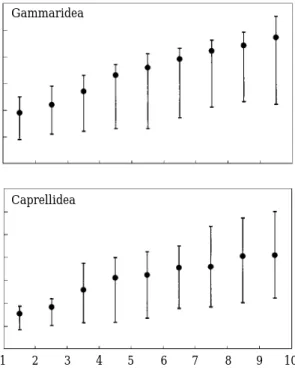
Relationships between size of amphipods and
body length of S. cirrhifer were pesented in Fig.
4. S. cirrhifer fed on larger sizes of prey with increasing fish size. In the case of gammarid amphipods, larger S. cirrhifer more than 4 cm SL fed on larger prey (3.3~4.8 mm, mean length) and caprellid amphipods (13.1~21.6 mm, mean length) were also consumed by larger individuals more than 3 cm SL.
The dietary breadth of S. cirrhifer was low value which is ranged from 0.17 to 0.28 (Fig. 5), i.e. S. cirrhifer targeted a restricted of prey spe- cies all size classes. High dietary breadth of the smaller individuals less than 2 cm SL declined with increasing fish size. This implied that S.
cirrhifer fed mainly on amphipods (gammarids and caprellids) with increasing their size.
Acknowledgements
We are grateful to Hyun Gi, Choo and Seong Oh, Im of Department of Oceanography, Pukyong University for assistance with sampling and data analysis. We also thank two anonymous referees for their constructive comments.
References
Adams, S.M. 1976. Feeding ecology of eelgrass fish communities. Trans. Amer. Fish. Soc., 105 : 514
~519.
Bell, J.D., J.J. Burchmore and D.A. Pollard. 1978.
Feeding ecology of three sympatric species of leatherjackets (Pisces: Monocanthidae) from a Posidonia seagrass habitat in New South Wales.
Aust. J. Mar. Fresh. Res., 29 : 631 ~643.
Chang, H.Y. 1991. Response of marine fish to the electric stimuli. Bull. Fish. Sci. Inst. Kunsan, 7:
21 ~30 (in Korean).
Conacher, M. 1977. Some aspects of the feeding ecol- ogy of the fanbellied leatherjacket Monacanthus chinensis, in seagrass beds. B.Sc. (Hons.) Thesis, University of Sydney.
Conacher, M.J., W.J.R. Lanzing and A.W.D. Lark- um. 1979. Ecology of Botany Bay. II. Aspects of the feeding ecology of Fanbellied Leatherjacket, Monocanthus chinensis (Pisces: Monocanthidae), in Posidonia australis seagrass beds in Quibray Bay, Botany Bay, New South Wales. Aust. J.
Mar. Fresh. Res., 30: 387 ~400.
Gibson, R.N. and I.A. Ezzi. 1987. Feeding relation- ships of a demersal fish assemblage on the wet coast of Scotland. J. Fish Biol., 31 : 55 ~69.
Huh, S.H. and S.N. Kwak. 1997a. Feeding habits of Pholis nebulosa. Korean J. Ichthyol., 9(1) : 22~
222 Seok Nam Kwak, Gun Wook Baeck and Sung-Hoi Huh
Fig. 4. Relationships between size of amphipods and body length of Stephanolepis cirrhifer (total length for Gammaridea and Caprellidea). solid circle and vertical bar represent the mean and range, respec- tively.
Fig. 5. The size-related variations of dietary breadth index of Stephanolepis cirrhifer.
1 2 3 4 5 6 7 8 9 10
Fish size (cm SL)
1 2 3 4 5 6 7 8 9 10
Fish size (cm SL) 6
5 4 3 2 1 0
Gammaridea
Caprellidea 35
30 25 20 15 10 5 0
Size of amphipods(mm)
0.3
0.2
0.1
0
Dietary breadth index
29 (in Korean).
Huh, S.H. and S.N. Kwak. 1997b. Feeding habits of Syngnathus schlegeli in eelgrass (Zostera mari- na) bed in Kwangyang Bay. J. Korean Fish.
Soc., 30(5) : 896 ~902 (in Korean).
Huh, S.H. and S.N. Kwak. 1997c. Feeding habits of Leiognathus nuchalis in eelgrass (Zostera mari- na) bed in Kwangyang Bay. Korean J. Ichthyol., 9(2) : 221 ~227 (in Korean).
Huh, S.H. and S.N. Kwak. 1997d. Species composi- tion and seasonal variations of fishes in eelgrass (Zostera marina) bed in Kwangyang Bay. Korean.
J. Ichthyol., 9(2) : 202 ~220 (in Korean).
Kikuchi, T. 1974. Japanese contributions on con- sumer ecology in eelgrass (Zostera marina L.) beds, with special reference to trophic relation- ships and resources in inshore fisheries. Aqua- culture, 4 : 145~160.
Kikuchi, T. 1966. An ecological study on animal communities of the Zostera marina belt in To- mioka Bay, Amakusa, Kyushu. Publ. Amak.
Mar. Biol. Lab., 1 : 1 ~106.
Kim, I.S. and Y.J. Kang. 1993. Coloured Fishes of Korea. Academy Publishing Co, Seoul., 477pp.
Kim, Y.U., K.H. Han, C.B. Kang and J.R. Koh.
1994. Distribution of ichthyoplankton in Kori, Korea. Bull. Kor. Fish. Soc., 27(5) : 633 ~642 (in Korean).
Klumpp, D.W., R.K. Howard and D.A. Pollard. 1989.
Trophodynamics and nutritional ecology of sea- grass communities. In: Larkum, A.W.D., A.J.
McComb and S.D. Shepherd (ed.), Biology of sea- grasses: A treatise on the biology of seagrasses with special reference to the Australian region.
Elsevier Science Publishers. B. V., New York/
Amsterdam, pp. 394 ~437.
Krebs, C.J. 1989. Ecological methodology. Harper and Row. New York, 654pp.
Kwak, S.N. 1997. Biotic communities and feeding ecology of fish in Zostera marina bed off Dae Island in Kwangyang Bay. Ph.D. thesis. Univer- sity of Pukyong. Korea, 411pp.
Last, P. 1975. Aspects of the taxonomy and ecology of Tasmanian leathejackets (family Monocanthi- dae, Pisces). BSc. (Hons.) thesis, University of Tasmania. 78pp.
Pollard, D.A. 1984. A review of ecological studies on seagrass fish communities, with particular ref- erence to recent studies in Australia. Aquat.
Bot., 18 : 3 ~42.
Randall, J.E. 1967. Food habits of reef fishes of the West Indies. Stud. Trop. Oceano., 5: 665~847.
Yang, Y.R. 1980. Phototaxis of filefish, conger eel and crucian carp. Bull. Kor. Fish. Soc., 13(1) : 1 ~13. (in Korean)
Received : October 11, 2003 Accepted : December 8, 2003
잘피밭에 서식하는 쥐치의 먹이습성 곽 석 남∙백 근 욱∙허 성 회*