보 문
논과 밭 토양의 황산염 환원세균 군집 구조 비교
이중배 ・ 박경량*
한남대학교 대덕밸리캠퍼스 생명시스템과학과
Comparison of community structure of sulfate reducing bacteria in rice paddy and dry farming soils
Jung Bae Lee and Kyeong Ryang Park*
Department of Biological Science and Biotechnology, Hannam University, Daejeon 305-811, Republic of Korea
(Received February 12, 2015; Accepted March 26, 2015)
ABSTRACT: The goal of this study was to identify relationships between the composition of sulfate reducing bacterial assemblages and terminal restriction fragment length polymorphism (T-RFLP) patterns in rice paddy and dry farming soils. Samples of organic farming soils, conventional farming soils, and dry field farming soils were collected in August and November. Analyses of the soil chemical composition revealed similar total nitrogen, total carbon and total inorganic phosphorus levels; however, the moisture content and total carbon were higher than in the other soils in both August and November, respectively. Sulfate reducing bacteria utilizing lactic acid were more widely distributed than those that used acetic acid, and the number of sulfate reducing bacteria in organic farming soil was most abundant. Phylogenetic analysis based on 181 clones revealed that most showed low similarity with cultured sulfate reducing bacteria, but more than 90% similarity with an uncultured sulfate reducing bacteria isolated from the environment. T-RFLP analysis revealed that fragments of 91, 357, 395, and 474 bp were most common, and the community structure of sulfate reducing bacteria changed seasonally.
Key words: dry farming soil, phylogenetic tree, rice paddy soil, sulfate reducing bacteria, T-RFLP
*For correspondence. E-mail: [email protected];
Tel.: +82-42-629-8770; Fax: +82-42-629-8769
미생물에 의한 유기물 분해는 산소를 최종 전자수용체로 사용하는 호기 호흡과 산소가 존재하지 않는 혐기 환경에서 산소 이외의 무기물을 최종 전자수용체로 사용하는 혐기 호 흡, 그리고 유기물을 최종 전자수용체로 사용하는 발효과정으 로 구분할 수 있다(Wind and Conrad, 1997). 이중 혐기 호흡은 질산염이 존재하면 탈질(denitrification) 과정을 통해, 망간과 철 같은 금속이온이 존재하면 금속이온 환원, 황산염이 존재 하면 황산염을 황화수소로 환원하여 에너지를 획득한다.
일반적으로 물에 잠긴 토양은 혐기 환경을 유지하지만, 질 산염, 철, 망간 등의 금속이온 농도는 매우 낮아 이들을 전자수 용체로 이용하는 미생물 활성은 높지 않다(Westermann, 1993). 대신 물에 잠긴 혐기 환경에서 나타나는 가장 일반적인 탄소순환은 메탄 생성(methanogenesis)과 황산염 환원(sulfate reduction) 과정이다. 그러나 보통 담수 환경은 황산염의 농도
가 낮아 대부분의 혐기성 분해는 메탄 생성과정으로 진행된 다. 그 반면 해양과 염습지(salt marshes)는 적당량의 황산염이 함유되어 있어 혐기 분해의 약 70%가 황산염 환원과정으로 이 루어지나(Barton and Tomei, 1995), 일부 연구 결과에 의하면 담수 환경도 해양이나 염습지처럼 황산염 환원 과정도 비교적 많이 진행되는 것으로 확인되고 있다(Detmers et al., 2001).
일반적으로 세균의 분포와 다양성 조사방법으로 여러 분자 생물학적 방법이 이용되고 있다. 이 중 16S rRNA 유전자를 기 초로 한 분석은 미생물 다양성과 분포에 대한 다양한 정보를 제공한다. 그리나 배양이 불가능하거나 특정 미생물을 확인할 경우 해당 미생물이 가지고 있는 functional gene을 이용하여 연구한다(Leloup et al., 2006). 이와 같은 functional gene 중 모 든 황산염 환원세균에서 발견되는 이화성 황산염 환원효소 (EC 1.8.99.1; dissimilatory sulfate reductase, dsr)는 황산염 환 원에 필수적인 효소로, 황산염(SO42-)을 아황산염(SO32-)로 환 원하는 효소다. 따라서 약 1.9 kb 크기의 α-subunit (dsrA)와 β-
subunit (dsrB)의 두 부분으로 구성되어 있는 이 유전자는 황산 염 환원세균의 분포와 다양성 연구에 이용된다(Wagner et al., 1998).
본 연구는 친환경 농업에 대한 관심이 증가하고 있는 현 시 점에서 친환경 논 농사인 유기 농법과 농약을 사용하는 관행 농법 논 토양, 그리고 비료와 농약을 사용하는 고추 밭 토양에 존재하는 황산염 환원세균의 분포와 다양성 차이를 확인한 연 구로, 작물의 생장이 가장 왕성한 8월과 추수 후인 11월에 토 양을 채취하여 계절에 따라 논과 밭 토양에 분포하는 황산염 환원세균의 군집 구조 변화를 이화성 황산염 환원효소를 이용 하여 조사하였다.
재료 및 방법
토양시료의 채집 및 세균 분리
전환기를 2년이상 경과하여 친환경 농법으로 인증 받고, 생 물농약은 사용하지 않으면서 우렁이를 이용하여 농사를 하고 있는 유기 농법 토양과 농약과 비료를 사용하여 농사를 하고 있는 관행 농법 토양, 그리고 고추를 재배하는 밭 토양 등 총 3 종류 토양 시료를 본 실험에 사용하였다. 토양은 작물이 성장 하는 시기와 추수한 후인 8월과 11월, 총 2회에 걸쳐 충북 오창 면 가곡리, 석우리, 탑리, 신평리 등에서 채취하였다. 토양 채 취는 농작물의 근권에 가까운 부위의 10 cm 이상 깊이에서 농 법 별로 3지점, 각 지점마다 일정 반경의 3곳에서 토양을 채취 한 후 하나의 sampling bag에 혼합하여 그 토양을 대표하는 대 표시료로 사용하였다. 채취한 토양 시료는 4℃를 유지하여 실 험실로 운반하였고, DNA를 분리하기 위한 토양은 –80℃에서, 나머지 토양은 4℃로 보관하여 사용하였다. 그리고 혐기 배양 은 basal carbonate-yeast extract-trypticase (BCYT) 배지(Touzel and Albagnac, 1983)를 기본 배지로 하여 실온에서 배양하였다.
토양성분 분석
토양 내 수분은 건토 중량법으로 건조기(FO-600M, Jeio tech)에서 110℃에서 24시간 건조한 후 무게를 측정하였고, 총 질소(total nitrogen; TN)는 Kjeldahl 법으로 측정하여 0.001 N H2SO4로 적정하였다. 총 탄소(total carbon; TC)는 0.05 g의 시 료를 탄소 측정기(SSM-500A, Shimadzu)를 이용하여 측정하였 고, 총 무기인(total inorganic phosphorus; TPi)은 1 g의 시료를 처 리하여 1-amino-2-naphtol-4-sulfonic acid로 발색 시킨 후 440 nm 에서 분광기(Lambda EZ201, Perkinelmer)로 측정하였다.
황산염 환원세균 수의 조사
토양시료 내에 기질로 초산과 젖산을 이용하는 황산염 환 원세균의 수는 MPN (most probable numbers) 방법(Balch et al., 1979)으로 실험한 후, 그 결과를 most probable numbers calculator (version 4.04, United States Environmental protection agency, Ohio) 프로그램을 이용하여 산정하였다.
Dissimilatory sulfate reductase (dsr)의 PCR (polymerase chain reaction)
Ultra Clean Soil DNA kit (MoBio)을 이용하여 토양세균의 DNA를 분리한 후, GeneAmp PCR system 9700 (Perkin- Elmer)을 사용하여 약 1.9 kb 크기의 황산염 환원 유전자를 증 폭 하였다. 이때 primer는 DSR 1F (5'-ACS CAC TGG AAG CAC G-3')와 DSR 4R (5'-GTG TAG CAG TTA CCG CA-3') 을 사용하였으며, PCR 반응은 initial denaturation (95℃, 15 min)을 시행 후, denaturation (94℃, 30 sec), annealing (58℃, 30 sec), elongation (72℃, 90 sec)을 30회 반복한 후 최종적으 로 elongation (72℃, 5 min)을 실시하였다(Castro, 2002). 그리 고 terminal restriction fragment length polymorphism (T- RFLP) 실험은 5' 위치에 6-FAM (carboxyfluorescein) 형광을 표지 한 DSR 1F을 사용하였고(Wagner et al., 1998), 이때 PCR 은 annealing 온도를 58℃ 대신 54℃로 낮추어 수행하였다.
PCR 반응이 끝난 후 5 µl PCR products를 0.7% agarose gel (0.5 x TBE buffer)에서 전기영동 하여 PCR 산물을 확인하였다.
dsr 유전자 염기서열 분석
dsr primer로 증폭한 PCR product를 pGEM-T Easy Vector (Promega)와 3:1 비율로 적정하고 T4-DNA ligase를 첨가한 후 4℃에서 12시간 동안 ligation 시켰다. 여기서 얻어진 ligate 는 1 ml X-gal (20 mg/ml)과 100 µl IPTG (20 mg/ml) 그리고 1 ml ampicillin (20 mg/ml)이 포함된 LB plate에 도말하여 blue-white colony 선별법으로 선별하였다. 선별된 colony는 DSR 1F와 DSR 4R primer를 이용하여 direct PCR을 통해 insert를 확인하고, insert가 확인된 clone의 PCR product를 HhaⅠ (Promega) 제한효소로 처리하고 4% agarose gel (0.5 x TBE buffer)로 각 clone의 유의성을 비교하였다. 이중 1% 이상 의 빈도를 차지하는 group을 선정한 후 ABI3730-XL Capillary DNA Sequencer (Applied Biosystems)로 염기서열을 분석하 고, 이를 GenBank (http://ncbi.nlm.nih.gov)와 비교한 후 BLASTN 프로그램으로 상동성 검색을 수행하였다. 이때 sequence alignment는 Bioedit (version 5.0.9) program으로 실
Table 1. Chemical composition of the rice paddy and dry field farming soils Field station Sampling Moisture
(%)
TN (g/kg)
TC (g/kg)
TPi (mg/kg)
Organic August November
26.21 0.13 1.63 57.0
26.77 0.13 1.57 54.3
Conventional August November
21.96 0.11 1.16 38.3
20.87 0.12 3.07 51.7
Dry-field August November
16.15 0.11 1.20 113.2
13.33 0.11 1.14 110.5
TN, total nitrogen; TC, total carbon; TPi, total inorganic phosphorus
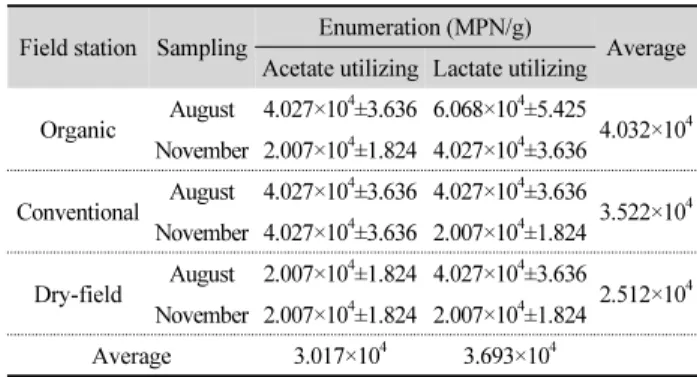
Table 2. MPN enumerations of sulfate reducing bacteria Field station Sampling Enumeration (MPN/g)
Average Acetate utilizing Lactate utilizing
Organic August 4.027×104±3.636 6.068×104±5.425
4.032×104 November 2.007×104±1.824 4.027×104±3.636
Conventional August 4.027×104±3.636 4.027×104±3.636
3.522×104 November 4.027×104±3.636 2.007×104±1.824
Dry-field August 2.007×104±1.824 4.027×104±3.636
2.512×104 November 2.007×104±1.824 2.007×104±1.824
Average 3.017×104 3.693×104
All values are expressed in units of MPN per gram (wet weight) of soil.
Each value represents the average of three independent experiments.
행하여 검토하고, 정렬된 염기자료 행렬은 Molecular Evolu- tionary Genetics Analysis (MEGA) software version 5.05 (Tamura et al., 2011)로 Kimura 2- parameter model (Kimura, 1980)을 이용하여 Neighbor-joining 방법(Saitou and Nei, 1987)으로 각각의 계통수를 작성하였고, 신뢰성 확보를 위해 bootstrap (Felsenstein, 1985)을 1,000회 수행하였다.
T-RFLP 분석
T-RFLP의 분석은 제한효소 RsaI을 사용하여 처리한 후, Genetic Analyzer (ABI PRISM® 3100; Applied Biosystems)로 분석하였고, T-RFLP분석을 통해 얻은 signal data는 Genotyper (version 3.7NT, Applid Biosystem)로 peak area를 계산하였 다. 이후 Excel (version 2010, Microsoft Corporation)로 전환 하여 noise 등을 제거한 후 Minitab (Release 14.1, Statistical software)으로 PCA profile을 작성하였다.
결 과
논과 밭 토양의 성분 및 함량
토양의 성분 분석 결과 수분 함량이 평균 약 20.88%, 총 질 소 0.12 g/kg, 총 탄소 1.62 g/kg, 총 무기인 70.8 mg/kg로 확인 되었다(Table 1). 수분 함량은 유기 농법서 평균 약 26.49%, 관 행 농법에서 약 21.42%, 고추 밭에서 약 14.74%로 밭 농사보 다는 논 농사의 수분 함량이 높았고 유기 농법이 관행 농법의 수분 함량 보다 높게 나타났다. 총 질소는 유기 농법에서 평균 약 0.13 g/kg, 관행 농법 약 0.11 g/kg, 밭은 약 0.11 g/kg로 토양 에 따라 큰 차이가 나타나지 않았다. 총 탄소는 유기 농법이 약 1.60 g/kg, 관행 농법 약 2.12 g/kg, 밭 약 1.14 g/kg로 수분 함량 과 마찬가지로 밭 토양보다는 논 토양의 총 탄소량이 높게 나 타났고, 논 농사법 사이에서는 관행 농법이 유기 농법의 탄소
량 보다 조금 높은 것으로 확인되었다. 총 무기인은 유기 농법 에서 평균 약 55.65 mg/kg, 관행 농법에서는 약 45 mg/kg, 고추 밭에서는 약 111.85 mg/kg로 밭 토양의 총 무기인 량이 논 토 양 보다 두 배 가량 높은 것으로 확인되었는데, 이는 밭에 비료 를 많이 주기 때문에 나타난 결과로 추정된다.
계절별로는 수분 함량은 유기 농법을 제외한 토양에서 11 월에 감소하였고, 총 질소는 유기 농법을 제외한 두 토양에서 11월에 증가하였다. 그리고 관행 농법의 토양만 총 탄소량이 11월에 증가하였고, 총 무기인은 유기 농법에서는 11월에 감 소하고, 관행 농법은 11월에 증가하였다. 그러나 밭은 8월과 11월에 모두 총 무기인이 110 mg/kg으로 비교적 높은 함량을 나타냈다.
논과 밭 토양의 황산염 환원세균의 분포
8월과 11월 토양 시료에서 초산과 젖산을 이용하는 황산염 환원세균의 분포를 MPN으로 확인한 결과, 유기 농법 토양 시 료에서 평균 4.032 × 104 MPN/g, 관행 농법에서 평균 3.522 × 104 MPN/g, 밭 토양에서는 평균 2.512 × 104 MPN/g의 균이 확 인 되었다(Table 2). 토양 별로는 일반적으로 논 토양의 황산염 환원세균이 밭 토양보다 더 많이 분포하고, 유기 농법의 토양 에 관행 농법의 토양보다 더 많은 황산염 환원세균이 분포하 고 있는 것으로 나타났다.
기질 별로는 아세트산을 이용하는 황산염 환원세균은 평 균 3.017 × 104 MPN/g, 젖산을 이용하는 황산염 환원세균은 평균 3.693 × 104 MPN/g으로 나타나, 전체적으로 젖산을 이 용하는 황산염 환원세균이 비교적 많이 분포하는 것으로 확 인되었다. 계절적으로는 대체로 여름에서 가을로 갈수록 황 산염 환원세균의 분포가 조금씩 줄어들었지만 큰 차이를 나타 내지 않았다.
Fig. 1. Neighbor-joining dsr α-subunit tree obtained from organic farming on August. The clones are named according to the agricultural methods and the season. Bootstrap values (expressed as percentages of 1,000) are shown at branching points. Bar, 0.05 substitution per nucleotide position.
Fig. 2. Neighbor-joining dsr α-subunit tree obtained from organic farming on November. The clones are named according to the agricultural methods and the season. Bootstrap values (expressed as percentages of 1,000) are shown at branching points. Bar, 0.05 substitution per nucleotide position.
황산염 환원세균의 군집 구조
논과 밭에서 분리된 총 181개 황산염 환원세균의 clone으로 phylogenetic tree를 작성한 결과 총 9개의 cluster가 확인되었 고(Figs. 1-6), 이중 알려지지 않는 황산염 환원세균의 그룹은 별도로 cluster 10에 배열하였다. Cluster DSR-1은 18개의 clone이 속하였고 주로 논 토양에서 많이 나타나며, 밭 토양은
8월 토양에서 일부 확인 되었다(Figs. 1-3, 5). Cluster DSR-2는 전체 클론 중 가장 많은 26개의 클론을 포함하고 논 토양과 밭 토양에 골고루 분포하고 있었다(Figs. 1-5). Cluster DSR-3은 DSR-2 다음으로 두 번째 많은 22개의 클론들을 포함하고 있 으며 조사된 모든 토양에서 나타났다(Figs. 1-5). Cluster DSR-4는 4개의 clone이 속하며 8월의 유기 농법 토양(Fig. 1) 과 관행 농법 토양(Fig. 3), 그리고 밭 토양(Figs. 5 and 6)에서
Fig. 3. Neighbor-joining dsr α-subunit tree obtained from conventional farming in August. The clones are named according to the agricultural methods and the season. Bootstrap values (expressed as percentages of 1,000) are shown at branching points. Bar, 0.05 substitution per nucleotide position.
Fig. 4. Neighbor-joining dsr α-subunit tree obtained from conventional farming in November. The clones are named according to the agricultural methods and the season. Bootstrap values (expressed as percentages of 1,000) are shown at branching points. Bar, 0.05 substitution per nucleotide position.
확인되었다. Cluster DSR-5는 전체의 시료 중 계절에 관계없 이 가장 고른 분포를 가지는 그룹으로 18개의 clone이 이 그룹 에서 나타났다(Figs. 1-6). Cluster DSR-6은 14개의 클론이 속 해 있으며 11월의 관행농과 8월과 11월의 유기농과 밭 토양에 서 확인되었다(Figs. 1, 2, 4, 6). Cluster DSR-7은 11월 유기 농 법 토양의 1개의 clone (Fig. 2)이, 그리고 11월 관행 농법 토양 에서 발견된 1개의 clone은 cluster DSR-8 (Fig. 4)에, 11월 밭
토양의 1개 clone은 cluster DSR-9에 위치하였다(Fig. 5). 그리 고 cluster DSR-10에는 13개의 클론이 속해 있으며 이들 클론 끼리의 상동성은 거의 99%로 나타났다.
DSR clone library의 계절에 따른 변화
계절별로 클론들의 변화를 확인해 본 결과, 유기 농법에서 는 8월에 cluster DSR-5과 DSR-1 (60%)이 높은 비율을 차지하
Fig. 5. Neighbor-joining dsr α-subunit tree obtained from dry field farming in August. The clones are named according to the agricultural methods and the season. Bootstrap values (expressed as percentages of 1,000) are shown at branching points. Bar, 0.05 substitution per nucleotide position.
Fig. 6. Neighbor-joining dsr α-subunit tree obtained from dry field farming in November. The clones are named according to the agricultural methods and the season. Bootstrap values (expressed as percentages of 1,000) are shown at branching points. Bar, 0.05 substitution per nucleotide position.
였지만, 11월의 토양에서는 cluster DSR-3과 DSR-6 (45%)의 비중이 높아지고 cluster DSR-7 (4%)이 새롭게 나타나는 것을 확인하였다(Fig. 7A and B). 그리고 관행 농법에서는 8월에 cluster DSR-1과 DSR-3 (75%)이 가장 많이 분포하지만, cluster DSR-1은 11월 토양에서는 나타나지 않고 cluster DSR-3의 분 포도 줄어들었지만, 8월 토양에 비해 cluster DSR-10의 분포
는 약 40%로 정도 높아진 것을 확인하였다(Fig. 7C and D). 밭 토양의 경우 8월에는 cluster DSR-2가 가장 많은 비중(44%)을 차지하고, 11월에는 DSR-2의 분포가 줄어들고 다른 cluster 들이 골고루 분포되어 나타나는 것을 확인할 수 있었다(Fig.
7E and F). 또 밭 토양의 경우 논 토양의 클론들 보다 더 다양한 클론들이 분포하였다.
Fig. 7. Seasonal distribution of DSR clones in organic farming soil for August (A) and November (B), conventional farming soil on August (C) and November, (D) and dry field farming soil for August (E) and November (F).
Fig. 8. PCA ordering generated from T-RFLP profiles for dsr gene in August and November soils. O1, organic farming soil for August (circle);
02, organic farming soil for November (triangle); C1, conventional farming soil for August (squares); C2, conventional farming soil for November (inverted triangle); T1, dry field farming soil for August (diamond); T2, dry field farming soil for November (cross).
T-RFLP를 이용한 PCA profile 분석
T-RFLP 분석 결과 밭 토양 시료 하나가 다른 14개의 시료들 과는 큰 차이를 나타내 이 시료의 data는 제거 후 분석하였다.
그 결과 8월과 11월의 유기 농법, 관행 농법 그리고 밭 토양의 황산염 환원세균이 분리되어 나타나는 것을 확인하였다(Fig.
8). 논 농법의 시료의 경우 각 토양들은 계절에 따라 황산염 환 원세균들이 분리되어 나타났지만, 유기 농법과 관행 농법의 황산염 환원세균은 분리되지 않았다. 논 토양과 밭 토양의 경 우 8월의 유기 농법이 밭 토양과 겹쳐져 나타났으나 계절의 변 화에 따라 11월의 경우 유기 농법과 밭 토양이 분리되어 나타 났다. 전체적으로 각각의 토양에 따라 계절별로 황산염 환원 세균이 분리되어 나타났으나 8월의 유기 농법과 관행 농법, 그 리고 유기농법과 밭 토양이 겹쳐져 나타나고, 11월 시료의 경 우 관행 농법과 밭 토양이 겹쳐져 나타났다.
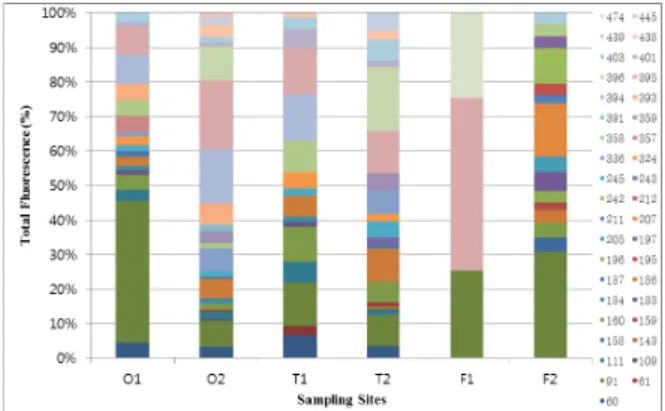
Fig. 9. Community dynamics on the dsr gene in soils sample by T-RFLP analysis. Community dynamics for the dsr gene in organic, conventional and dry field farming soils determined by T-RFLP analysis. Y-axis values represent percentages of total fluorescence. Samples are labeled according to the agricultural methods and the season. O, organic farming soil; C, conventional farming soil; T, dry field farming soil; 1, August; 2, November.
T-RFLP 군집구조
T-RFLP에서 확인된 염기를 비교한 결과(Fig. 9), 8월의 시 료는 60, 61, 91, 111, 160, 183, 184, 186, 187, 245, 324, 336, 357, 358, 393, 394, 401, 403, 438, 445, 474 bp의 총 21개 bp가, 11월 시료에서는 60, 91, 109, 111, 143, 158, 159, 160, 184, 186, 187, 195, 196, 197, 205, 207, 211, 212, 242, 243, 245, 324, 336, 358, 359, 391, 393, 394, 395, 396, 401, 403, 438, 439, 445 bp의 총 35개 bp가 나타났다. 이중 8월의 유기 농법 토양에서 는 91 bp의 peak area가 가장 높았고 11월에는 336 bp와 357 bp 의 peak area가 높았다. 또 8월의 관행 농법 토양은 여러 bp가 골고루 분포되었으나 11월의 토양은 유기 농법과 동일하게 336 bp와 357 bp의 peak area가 높았다. 밭 토양의 경우 8월에 는 91 bp, 357 bp, 474 bp의 peak area가 높았으나, 11월에는 91 bp만이 비교적 높고, 여러 bp가 골고루 분포하고 있어 다양한 미생물 군집이 분포하고 있음을 확인하였다.
고 찰
논과 밭 토양의 화학적 성분을 분석하고, 미생물이 가지고 있는 functional gene 중 이화성 황산염 환원효소를 대상으로 클론 라이브러리를 구축하고, RFLP (restriction fragment length polymorphism)와 T-RFLP 패턴을 분석하여 황산염 환원세균 의 분포와 다양성 그리고 군집구조를 파악하였다.
본 실험에 사용된 유기 농법, 관행 농법, 그리고 밭 토양의 수분 함량 분석 결과 장마철이 지난 8월에 채취한 토양의 수분
함량이 가장 높았고, 겨울로 갈수록 전반적으로 낮아지는 경 향을 보였다. 그러나 유기 농법 토양은 수분함량이 계절별로 차이가 없었고 다른 토양에 비해 비교적 높은 수분 함량을 나 타내었다. 이 결과는 국내 금화지구 3개 초지 토양의 수분함량 인 20–40%와 비슷하지만(Park and Chang, 1994), 미국 플로 리다 주 Everglades의 습지 토양 수분 함량 92.67%에 비해 매 우 적은 함량이다(Chauhan et al., 2004). 토양의 총 탄소는 11 월 관행 농법 토양의 함량이 가장 높았고 전체적으로 관행 농 법의 토양을 제외하고 여름에 채취한 토양의 총 탄소량이 약 간 높은 것을 알 수 있었다. 총 질소의 함량은 전체 토양이 비슷 하였으며, 8월보다 11월의 함량이 약간 많은 것으로 나타났으 나, 김해지구 초지 토양의 총 질소 함량(Lee et al., 1998)과 비 교 하였을 때 매우 낮은 함량을 나타내었다.
초산과 젖산을 이용하는 황산염 환원세균은 모든 토양에서 평균 104 MPN/g 정도의 균이 존재하고, 논과 밭 토양의 비교 에서는 논 토양에 더 많은 황산염 환원세균이 분포하였다. 논 토양끼리의 비교에서는 유기 농법 토양이 관행 농법의 토양 보다 황산염 환원세균의 분포가 우세하였다. 그러나 본 실험 에서 검출된 황산염 환원세균의 수는 미국 플로리다 주 Everglades의 습지 중 인산에 의해 오염된 F1 (eutrophic) 지역 에 존재하는 황산염 환원균수 1.5 × 106 MPN/g, 청정지역인 U3 (oligotrophic)에 존재하는 황산염 환원균수 9.2 × 105 MPN/g의 균 보다 10배에서 많게는 100배 더 적게 분포하고 있는 것을 알 수 있었다(Castro, 2003).
dsr 유전자를 이용한 계통학적 분석에서 유기 농법 토양과 관행 농법 토양, 밭 토양에서 분리된 총 181개 clones을 Thermodesulfovibrio islandicus를 outgroup으로 하여 phylogenetic tree를 분석한 결과 총 10개의 분류군이 나타났다. 이중 DSR-1 cluster는 미국 플로리다 습지(Castro et al., 2002)와 중국 황토 고원의 clone과, DSR-2는 오스트레일리아의 메콰리 늪지 (Gavin et al., 2010), 아제르바이잔의 화산재(Saxena, 2013), 중국 타오위안현 논 토양(He et al., 2010)과 해안 매립지(Wu et al., 2009), 독일 산성 지역(Doris et al., 2011)의 클론과, DSR-3 는 미국 일리노이주의 산성광산 배수(Burns et al., 2012)와 산 타바바라의 대륙주변(Benjamin et al., 2009)의 클론과, DSR-4 는 중국 주장강에서 확인된 클론(Lijing et al., 2009)과, DSR-6 은 독일의 습지(Pester et al., 2012)와 미국 Oak Ridge의 집수 구역(Hansel et al., 2008), 뉴욕의 양파 재배 토양의 클론 (Carmen et al., 2007)과, DSR-8은 미국의 혐기적 슬러지 공법 의 생물 반응조 클론(Kovacik et al., 2010)과 높은 상동성을 갖 는 것으로 확인되었다. 따라서 본 실험에서 분리된 181개의 clone 중 131개 clone이 현재까지 알려진 배양 가능한 황산염
환원세균과 매우 낮은 상동성을 나타내고 uncultured 클론들 과 유사함이 확인되어 지구의 혐기성 환경에는 다양한 황산염 환원세균이 분포함을 다시 한번 확인하였다. 또 DSR-4는 Desulfobacterales 목과 80–84%의 상동성을, 계절에 관계없이 가장 고른 분포를 가지는 DSR-5는 Syntrophobacterales 목과 70–80%의 상동성을, DSR-6는 Syntrophobacterales 목과 62–
70%의 상동성을, DSR-7은 Archaeoglobles 목과 높은 상동 성을, DSR-8는 Desulfovibrionales 목과 84%의 상동성을, DSR-9은 Clostridiales 목과 70–93%의 상동성을 나타내고, Clostridiales 목 중에서도 Desulfotomaculum aeronauticum와 93%로 가장 높은 상동성을 나타내어, 181개의 clone 중 cultured bacteria로 확인된 50개의 clone은 약 70%가 Syntrophobacterales 목에 속하는 것으로 확인되었다.
T-RFLP 분석 결과 유기 농법, 관행 농법, 밭 각각의 시료의 경우 8월과 11월 토양의 황산염 환원세균들이 모두 분리되어 나타나는 것을 확인하였다. 유기 농법과 관행 농법의 경우 계 절에 따라 분리되어 나타났고, 8월의 유기 농법 토양은 밭 토양 과 겹쳐 나타났으나 11월에는 분리되어 나타났다.
따라서 논과 밭 토양을 비교할 경우 밭 토양은 주년 재배로 경작을 하기 때문에 계절의 변화에 따라 다양한 황산염 환원 세균의 군집이 나타나고, 논 토양은 비교적 황산염 환원세균 군집구조의 변화가 적은 것으로 생각된다.
T-RFLP에서 관찰된 bp와 군집구조 비교분석 결과 8월의 유 기 농법 토양에서 91 bp의 peak area가 높았고, 11월로 갈수록 336 bp과 357 bp가 높아졌다. 또 8월의 관행 농법은 여러 bp의 비중이 골고루 분포되었으나 11월은 유기 농법과 같이 336 bp과 357 bp 가 높아졌다. 밭 토양의 경우 8월에는 91 bp, 357 bp, 474 bp 이 3개 의 bp가 높았으나, 11월에는 91 bp의 비중만이 늘어나고 다른 bp 는 골고루 분포하고 있어 다양성이 높아진 것을 확인하였다. 이 결과는 미국 플로리다 주 Everglades의 습지에서 검출된 클론들 의 bp 중 황산염 환원세균의 분포가 가장 많았던 41 bp와 188 bp 와는 다른 결과로(Castro, 2003), 토양의 종류에 따라 황산염 환 원세균의 종류가 달라짐을 다시 한번 확인할 수 있었다.
적 요
본 연구는 논과 밭 토양의 황산염 환원세균의 군집구조와 T-RFLP 패턴을 조사한 논문으로, 유기 농법 토양과 관행 농법 토양 그리고 밭 토양 총 3종류의 토양을 8월과 11월에 채집하 여 실험하였다. 토양 성분 분석 결과 총 질소, 총 탄소, 총 인의 값은 모든 토양이 비슷하게 나타났고 계절별로는 수분의 함량
은 8월에, 총 탄소는 11월에 가장 높게 나타났다. 황산염 환원 세균은 초산보다 젖산을 기질로 이용하는 황산염 환원세균이 더 많이 분포하고, 유기 농법 토양에 황산염 환원세균이 가장 많이 분포하는 것으로 나타났다. 각 토양에서 얻은 총 181개 클론으로 계통학적 분석을 한 결과, 대부분의 클론들은 배양 가능한 황산염 환원세균과는 매우 낮은 상동성을 보였으나, 자연계에서 확인되는 클론들과는 90% 이상의 높은 상동성을 나타내었다. T-RFLP 분석 결과 91, 357, 395, 474 bp의 분포가 가장 높았고, 계절에 따라 황산염 환원세균의 군집 구조가 달 라지는 것을 확인하였다.
감사의 말
이 논문은 2014년 한남대학교 학술연구 조성비 지원에 의 하여 연구되었음.
References