Phylogenetic Analysis of Carassius auratus and C. cuvieri in Lake Yedang Based on Variations of Mitochondrial CYTB Gene Sequences
Gye-Woong Kim, Sung-Duck Joe, Hack-Youn Kim and Hee-Bok Park*
Department of Animal Resources Science, College of Industrial Sciences, Kongju National University, Yesan 32439, Korea Received July 9, 2020 /Revised August 26, 2020 /Accepted September 7, 2020
Two crucian carp species (Carassius auratus and C. cuvieri) inhabit Lake Yedang in South Korea, and C. auratus is known to be native to Korea. Classification of these two freshwater fish species is often confused because of their morphological similarity. To distinguish the two species, we conducted phy- logenetic and population genetic analyses of C. auratus and C. cuvieri based on their mitochondrial DNA sequences of the cytochrome b gene (CYTB). We also compared our partial CYTB sequence (<1,056 bp) with 10 Chinese, nine Japanese, and two Russian crucian carp fishes. The results of our phylogenetic analysis showed that C. auratus and C. cuvieri were clearly divided into two phylogroups.
The nucleotide diversity (π) of C. auratus from Korea, China, and Japan showed a range of 0.146%~
0.421%, while the range of π of C. cuvieri from Korea and Japan was lower than those of C. auratus (0.0%~0.054%). Moreover, the comparison of CYTB divergence among crucian carp fishes in China, Japan, and Korea indicated that Korean Carassius fishes were distantly related to those from China and Japan, with two exceptions: the pairwise F
stvalue between Korean C. auratus and northern Chinese C. auratus was not significantly different. In addition, no significant genetic divergence be- tween Korean and Japanese C. cuvieri was detected. We conclude that, despite the morphological sim- ilarities, C. auratus and C. cuvieri should be considered as separate freshwater fish resources in con- servation efforts for genetic diversity.
Key words : Carassius auratus, C. cuvieri, CYTB gene, genetic diversity, phylogeny
*Corresponding author
Tel : +82-41-330-1244, Fax : +82-41-330-1249
*E-mail : [email protected]
This is an Open-Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/3.0) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
서 론
붕어는 조기어강(Actinopterygii), 잉어목(Cypriniformes), 잉 어과(Cyprinidae), 붕어속(Carassius)으로 분류 할 수 있다. 붕어 속에 속하는 대표적인 어종으로는 Carassius carassius (Lin- naeus, 1758), C. cuvieri Temmink and Schlegel, 1846, 그리고 C. auratus (Linnaeus, 1758)을 들 수 있다[16]. 여기서, C. aur- atus의 경우 지리적 분포에 따라 여러 아종(subspecies)로 구분 할 수 있는데, 구체적으로 살펴보면, C. a. auratus (중국), C.
a. gibelio (유럽, 일본 및 중국), C. a. cuvieri (일본), C. a. langs- dorfii (일본) 등으로 분류 할 수 있다[4]. 그리고, 금붕어(gold fish)는 관상용으로 개량된 붕어로서 학명이 C. a. auratus이다 [9].
한국 자연산 붕어는 1934년에 처음 학계에 보고되었는데, 함경북도 청진에서 채집되어 C. carassius로 분류된 경우를 제 외하고, Mori와 Uchida는 그 외의 모든 다른 한반도 서식 자연
산 붕어들을 C. auratus로 분류하여 보고한 바 있다[14]. 이 후, 남 등(1989)은 영남지역 수계에서 채집한 자연산 붕어를 C.
auratus로 보고 하였다[15]. 우리나라 담수역에는 C. auratus로 분류되어 있는 자연산 붕어 외에도 1970년대에 내수면 자원조 성을 목적으로 일본에서 도입한 C. cuvieri 도 널리 서식하고 있는데 일반적으로는 떡붕어로 알려져 있다[7]. 붕어속은 형태 학상 유사성과 다양성 때문에 현재 분류체계가 명확히 확립 되어있지 않다. 이러한 붕어속 분류의 제한을 극복하기 위하 여 RAPD (random amplified polymorphic DNAs)와 미토콘 드리아 유전체 염기서열 같은 DNA 표지들이 분자계통학 연 구에 이용되고 있다. 윤과 박(2006)은 RAPD 마커를 이용하여 예산과 당진 지역에 서식하고 있는 국내 자연산 붕어(C. aur- atus)와 떡붕어(C. cuvieri)간의 유전적 특성을 보고하였다[18].
정 등(2009)은 우리나라 자연산 붕어속 어류종에 존재하는 유 전적 변이를 미토콘드리아 displacement-loop (D-loop) 염기 서열을 이용하여 분석한 바 있고, 강 등(2014)은 우리나라 낙동 강, 영산강, 금강 유역의 C. auratus와 C. cuvieri 집단 간에서 유전적 다양성과 분자수준의 계통 유연관계를 미토콘드리아 cytochrome c oxidase I (COI) 유전자 염기서열의 다형성을 이용하여 연구한 결과를 보고 하였다[7].
충남 예산군에 위치한 예당호는 유역면적이 37,360 ha로서
대한민국의 농업용수 전용공급 저수지 중에서 가장 넓다. 예
당호에 서식하는 붕어속으로는 형태학상으로 분류된 한국산
Table 1. Species, sample collection region, number of individuals, source nucleotide sequences of genus Carassius and one outgroup species used in this study
Species name Sample collection region Na Genbank accession number
C.auratus
C.cuvieri
C.carassius Cyprinus carpiob
Korea (South) Korea (North) China (North) China (South)
Japan Korea (South)
Japan Russia
Japan
5 1 5 5 4 5 5 2 2
MT155797C, MT155798C, MT155799C, MT155800C, MT155801C KM261774.1
KJ735898.1, KJ735900.1, KJ735901.1, KJ735904.1, KJ735907.1 HQ689824.1, HQ689826.1, HQ689830.1, HQ689835.1, HQ689837.1 AB368691.1,AB368697.1, AB368698.1, AB368699.1
MT155802C, MT155803C, MT155804C, MT155805C, MT155806C HQ689913.1, AP011237.1, AB045144.1, NC_010768.1, LC097913.1 HQ689909.1, HQ689908.1
AB158807.1, AB158806.1
anumber of individuals; bindicates outgroup species in the phylogenetic analysis; cGenBank sequence ID obtained from this study.
붕어와 외래도입종 떡붕어가 보고 된 바 있다[8]. 본 연구는 두 붕어속 어종의 미토콘드리아 cytochrome b (CYTB) 유전자 염기서열과 기존에 미국 국립생물정보학센터(NCBI) Genbank 데이터베이스에 등록되어 있는 붕어속 어류의 동일유전자 염 기서열 자료를 이용하여, 붕어와 떡붕어간의 유전적 다양성을 추정하고, 분자계통분류학 분석을 통해 예당호 붕어속 어종과 중국, 일본, 러시아 붕어속 어종과의 유전적 특성을 구명하기 위하여 수행되었다.
재료 및 방법
공시동물 및 DNA 분리
충남 예당호 수계에서 형태학적으로 분류한 붕어와 떡붕어 각각 5마리씩으로부터 근육조직 샘플이 제공되어 본 연구에 사용 하였다. 공시동물은 공주대학교의 동물실험계획서에 의 거 공주대학교 동물실험윤리위원회에서 승인된 동물실험방 법(승인번호: 2020-04)에 따라 수행되었다. 이 근육 샘플의 게 놈 및 미토콘드리아 DNA는 Qiagen DNeasy blood&tissue kit (Qiagen, Germany)을 이용하여 분리하였다. 이 DNA는 1%
agarose 겔 전기영동을 통하여 확인하였고, NanoDrop ND-2000 spectrophotometer (Thermo Scientific, USA)로 순 도와 농도를 측정한 후 PCR 반응의 주형으로 사용하였다. 그 리고 북한산, 중국산, 일본산 및 러시아산 C. auratus, C. cuvieri 및 C. carassius의 CYTB 유전자 서열은 NCBI Genbank에 등록 된 염기서열 자료를 이용하였다. 참조 분류군(outgroup)은 잉 어과에 속하면서 분류학적으로 Carassius 속 어종에 가까운 잉 어(Cyprinus carpio)를 선정하여 사용하였다. 연구에 사용한 어 류들의 학명, 채집지역과 GenBank Accession Number는 Table 1에 나타내었다.
미토콘드리아 cytochrome b (CYTB) 유전자의 증폭과 염 기서열결정
추출한 DNA를 주형으로 미토콘드리마 CYTB의 증폭을 위
해서 PCR primer를 Primer3 프로그램을 이용하여 디자인 하 였고[10], 참조 염기서열은 NCBI Genbank에서 검색한 AP 011237.1 (Carassius cuvieri mitochondrial DNA, complete ge- nome)을 이용하였다. CYTB 유전자 1,141 bp 중에 1,056 bp에 해당하는 부위를 커버 할 수 있도록 597 bp 크기의 PCR 산물 을 증폭 할 수 있는 첫 번째 프라이머 쌍(CYTB_1F: 5‘-CTA GTT GAC CTA CCC ACA CCA-3’, CYTB_1R: 5‘-GTC TGC GTC CGA GTT TAA TCC-3’)과 729 bp 크기의 PCR 산물을 증폭 할 수 있는 두 번째 프라이머 쌍(CYTB_2-1F: 5‘-GCC TTT GTC GGT TAT GTC CTC-3’, CYTB_2-1R: 5‘-TGC TAG TGG GAA GAG GAC AAG-3’)을 디자인 하였다. PCR은 10×반응 완충액, 10 mM dNTP, 각각 5 pmole forward/reverse primer set, 1.0 units Taq DNA polymerase (GenetBio, Korea)와 50 ng DNA 용액에 멸균한 탈이온수를 첨가하였다. ABI-200 PCR (Perkinelmer, USA)을 이용하여 95℃ 5분 초기변성 후, 94℃
1분, 57℃ 1분, 72℃ 1분으로 구성된 연쇄반응을 35회 반복한 후 72℃에서 7분간 최종 신장하였다. 이 단일 PCR 조건으로 두 쌍의 프라이머 set을 증폭하였다. PCR 증폭 산물은 1.0%
agarose 겔 상에서 전기영동하여 확인하였다. 증폭된 PCR 산 물은 Bioneer PCR purification Kit (Bioneer, USA)을 이용하 여 정제하였고, 정제된 PCR 산물을 주형으로 PCR 증폭에 씌 였던 primer set을 이용하여 염기서열을 결정하였다(COSMO genetech, Korea).
DNA 염기서열 자료분석
결정된 염기서열들의 편집은 STADEN package의 Pregap4 와 Gap 프로그램을 이용하여 수행되었다[1]. 이를 통해 얻은 염기서열 자료의 다중염기서열정렬(multiple sequence align- ment)은 DIALIGN (version 2.2) 프로그램을 이용하였다[13].
염기서열 변이부위 탐색, 집단 내와 집단 간 유전적 다양성 추정은 DnaSP (ver. 5.1)를 사용하였다[12]. 추가적으로 집단 간 유전적 차이 정도를 나타내는 Weir and Cockerham’s F
st값을 추정하기 위해서 Arlequin (ver. 3.5)를 사용하였다[2, 17].
Fig. 1. Gel electrophoresis of PCR products using the two pri- mer set (CYTB_1F and CYTB_1R for the 597 bp products;
CYTB_2-1F and CYTB_2-1R for the 729 bp products) for distinction of two crucian carp species. M, 100 bp ladder size marker (GenetBio, Korea); lanes 1 and 3, C. auratus;
lanes 2 and 4, C. cuvieri; (-), negative control.
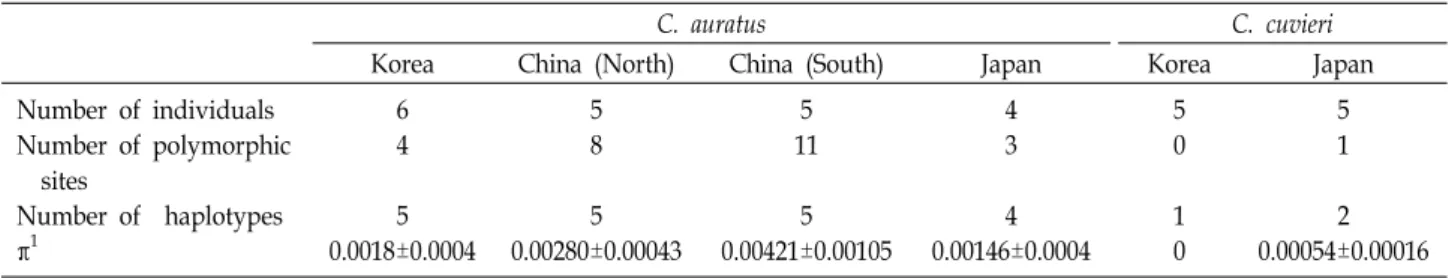
Table 2. Summary statistics showing the genetic diversity of C. auratus and C. cuvieri in China, Japan, and Korea
C. auratus C. cuvieri
Korea China (North) China (South) Japan Korea Japan
Number of individuals Number of polymorphic
sites
Number of haplotypes π1
6 4 5 0.0018±0.0004
5 8 5 0.00280±0.00043
5 11
5 0.00421±0.00105
4 3 4 0.00146±0.0004
5 0 1 0
5 1 2 0.00054±0.00016
1Nucleotide diversity (π) ± standard deviation of π
Weir and Cockerham’s F
st값은 일반적으로 0에서 1의 값을 취하는데, 0에 가까울수록 집단 간 유전적 차이의 정도가 낮 고, 1에 가까울수록 집단 간 유전적 차이의 정도가 높은 것으 로 해석 할 수 있다. 또한 Weir and Cockerham’s F
st값은 음의 값을 취 할 수도 있는데, 이는 유전적 차이의 정도가 낮은 것으 로 간주 할 수 있다[17]. 추정된 F
st값에 대한 통계적 유의성을 검정하기 위하여 10,000회의 Permutation을 실시하였으며, 통 계적 임계치는 5%로 하였다(p<0.05) [5].
분자 계통수의 작성은 Neighbor joining (NJ) 방법을 이용 하였다. NJ 방법에 기반 한 계통수 작성을 위해서는 Maxim- um composite likelihood 모델을 적용하였다. NJ 계통수는 MEGA7 package를 이용하였다[11]. 작성된 NJ 계통수의 in- ternal node에 대한 신뢰성을 확인하기 위하여 10,000회의 bootstrapping을 실시하였다(Felsenstein, 1985) [3].
결과 및 고찰
붕어 CYTB 유전자의 집단 내 다형성
본 연구에서는 Carassius 속에 속하는 어종들의 분자 계통분 류학적 관계를 알아보기 위하여 충남 예당호 수계에서 포획 한 붕어(C. auratus)와 떡붕어(C. cuvieri)의 CYTB 유전자 서열 을 PCR을 통하여 증폭하였다. PCR 산물 서열 길이는 각각 597
bp 와 729 bp를 나타내었고 이 PCR 산물의 염기서열을 확보 하였다(Fig. 1, Fig. 2) DIALIGN과 DnaSP 프로그램을 이용하 여 다중염기서열정렬과 단일염기다형성(single nucleotide polymorphism, SNP) 검출을 수행한 결과, 한국산 붕어의 경 우 4개의 SNP가 확인되었고 5개의 haplotype으로 구분되었 다. 떡붕어의 경우 SNP가 전혀 검출되지 않고 모든 개체들이 같은 haplotype으로 추정되었다(Table 2). 이러한 결과는 충남 예당호 수계에 서식하는 떡붕어들이 아주 제한된 숫자의 모계 에서 유래되었음을 추정 할 수 있다.
C. auratus와 C. cuvieri의 지역별 개체 간의 유전적 차이를 Nucleotide diversity (π)로 나타낸 결과는 Table 2와 같다. 한 국산, 일본산, 중국 북부산 C. auratus 의 π 는 0.0018-0.00280의 범위를 보여 줌으로서 채집 지역을 달리하는 세 집단의 개체 사이에 뚜렷한 차이를 보이지 않았다. 그러나 중국 남부산 C.
auratus 의 π 는 0.00421을 보여 줌으로서 다른 세 집단에 비하 여 높은 개체 간 유전적 차이를 보였다. Gao (2012) 등은 미토 콘드리아 D-loop 및 CYTB 유전자를 이용한 붕어(C. auratus) 의 유전적 다양성 연구에서 중국 남부지역에서 서식하는 붕어 의 유전적 다양성이 북부지역 붕어에 비해 높은 경향을 보인 다는 결과를 보고한 바 있다[4].
C. cuvieri의 경우, C. auratus에 비해서도 낮은 종내 유전적 변이 수준을 보였는데, 한국산 C. cuvieri 의 경우 π가 0이었고 일본산 C. cuvieri는 0.00054을 나타내었다.
붕어 CYTB 유전자의 집단 간 다형성
집단 간 유전적 차이 정도를 나타내는 Weir and Cocker- ham’s Fst 값, 집단 간 평균 염기서열 차이 및 Dxy 값을 추정 결과는 Table 3에 나타내었다. 전체 F
ST값의 범위는 -0.03~0.99 로 나타났다. F
ST값의 유의성을 살펴보면 한국산 붕어 집단 (KR_Ca)과 중국 북부산 붕어 집단(CN_Ca_Nor), 그리고 한국 산 떡붕어 집단(KR_Ccu)과 일본산 떡붕어 집단(JP_Ccu)을 제 외하고는 모든 집단 간 비교에서 유의한 차이가 관찰되었다 (p<0.05). 따라서, 우리나라, 중국, 일본의 붕어 및 떡붕어 집단 간의 유전적 분화 정도에서 우리나라와 중국 북부의 붕어 집단 과 우리나라와 일본의 떡붕어 집단 외에는 유전적 분화 정도 수치(F
ST) 상으로 유의적 차이가 나타난다는 것을 확인하였다.
이러한 결과는 집단 간 평균 염기서열 차이 및 D
xy값을
Fig. 2. An alignment set of partial mitochondrial CYTB gene sequences of C. auratus and C. cuvieri from Lake Yedang generated using the CLUSTALW program (URL:genome.jp/tools-bin/clustalw). Locations of PCR primers (CYTB_1F and CYTB_1R;
CYTB_2-1F and CYTB_2-1R) are indicated.
Table 3. Mitochondrial CYTB gene divergence between Carassius populations in China, Japan, and Korea
KR_Ca CN_Ca_North CN_Ca_South JP_Ca KR_Ccu JP_Ccu
KR_Ca - 0.0023±0.00073
(2.20)
0.014±0.0036 (13.27)
0.018±0.006 (17.17)
0.069±0.028 (63.67)
0.069±0.023 (64.07) CN_Ca
_North -0.03NS - 0.014±0.0035
(15.40)
0.016±0.0045 (18.20)
0.070±0.031 (65.00)
0.069±0.023 (73.60) CN_Ca
_South 0.77* 0.74* - 0.019±0.0053
(21.20)
0.073±0.0032 (67.80)
0.071±0.024 (75.80)
JP_Ca 0.89* 0.86* 0.84* - 0.074±0.037
(68.50)
0.072±0.027 (77.40)
KR_Ccu 0.98* 0.98* 0.97* 0.99* - 0.00041±0.00038
(0.40)
JP_Ccu 0.98* 0.98* 0.97* 0.99* 0.25NS -
KR: Korea, CN: China, JP: Japan, Ca: C. auratus, Ccu: C. cuvieri; Upper diagonal: average number of nucleotide substitutions per site between populations (Dxy) and its standard deviation; values in parenthesis represent the average number of nucleotide differences between populations. Lower diagonal: values represent the Weir and Cockerham’s Fst value between populations; asterisk represents significance support for Fst values (p<0.05), NS represents no significance.
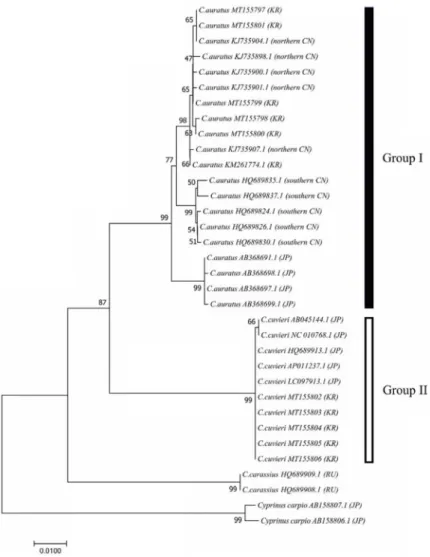
Fig. 3. A neighbour-joining (NJ) tree of CYTB gene sequence constructed to demon- strate the phylogenetic relationship of the genus Carassius and Cyprians carpio. The group I is composed of C. auratus and the group II consists of C. cuvieri. Numbers at the nodes represent the bootstrap sup- port values derived from 10,000 replicates.
The scale indicates the genetic distance.
통해서도 뒷받침 되었다. 우리나라 C. cuvieri와 일본산 C. cu- vieri 에는 평균 염기서열 차이가 0.4개(D
xy=0.00041)이었고, 우 리나라 C. auratus와 중국 북부지역에 서식하는 C. auratus 간 에도 염기서열 차이가 2.2개(D
xy=0.0023)로 추정됨으로서 밀접 한 유전적 유연관계를 보였다. 가장 큰 유전적 차이를 보인 두 집단은 일본산 C. auratus와 일본산 C. cuvieri였는데 평균염 기서열차이가 77.4개(D
xy=0.072)이었으며 높은 F
ST값(0.99)을 보였다. 우리나라 C. auratus와 C. cuvieri 간에도 평균 63.67개 (D
xy=0.069)의 염기서열 차이를 보였고 F
st값(0.98)도 높게 추 정되었다. 이러한 결과는 Kang 등(2014)이 우리나라 낙동강, 영산강, 금강 유역의 C. auratus와 C. cuvieri 집단을 COI 유전자 분석을 했을 때, 두 어종 집단이 서로 분리된다는 결과를 보고 한 것과 유사하여, 본 연구의 분석 결과를 뒷받침해 주는 것이 라 생각된다[7].
분자계통분류학적 관계
NJ 방법을 이용하여 Carassius 속 어류의 계통수를 작성하 였으며 Fig. 3에 나타내었다. 작성된 계통수에서 C. auratus, C. cuvieri, 그리고 C. carassius 종이 계통분류학적인 위치에서
각각의 phylogroup을 형성하였다.
C. auratus의 경우 북한산 붕어(NCBI acc. no. KM261774.1) 를 포함한 한국산 C. auratus가 중국 북부산 C. auratus와 한 그룹으로 묶여 매우 가까운 유연관계를 나타내었다. 이러한 결과와 유사하게 Jung 등(2009)은 미토콘드리아 D-loop 염기 서열을 이용한 분자계통분류학 분석을 통해서 중국 북부에서 포획된 sample이 일본에서 비롯된 sample 보다 남한 지역의 Carassius속 어류 종과 유전적 거리가 가깝다는 결과를 보고 한 바 있다[6]. Gao 등(2012)은 중국 북부산 C. auratus는 한반 도와 인접한 Heilongjiang 지역에서 포획 된 집단이며, 중국 남부산 C. auratus는 Anhui 지역에서 포획 된 집단으로 보고한 바 있다[4]. 이는 같은 C. auratus종 이지만 한반도 및 중국 북부 산, 중국 남부산 그리고 일본산 붕어의 경우 계통분류학적으 로 차이를 보임으로서 분포지역에 따른 유전적 분화 양상이 관찰되었다.
C. cuvieri 종의 경우 한국산과 일본산이 같은 그룹으로 묶임 으로서 분포지역에 따른 유전적 다형성을 나타내지 않았다.
이것은 1970년대에 내수면 자원조성을 목적으로 도입된 일본
산 C. cuvieri가 40년이 넘는 지역적인 격리에도 불구하고 유의
적인 유전적 분화 양상이 일어나지 않은 것으로 생각된다.
본 연구에서 충남 예당호 수계에서 서식 하고 있는 붕어와 떡붕어는 서로 다른 종으로서 유전적 거리를 확인 할 수 있었 고, 3개국 붕어 속 집단 간의 유연관계 분석에서는 우리나라 붕어와 중국 북부 붕어 간 밀접한 유전적 유연관계를 확인 할 수 있었다. 또한, 우리나라와 일본 서식 떡붕어 간의 지역적 격리에 따른 유전적 다양성을 관찰 할 수 없었다. 그 밖의 붕어 속 집단 간에는 유의한 유전적 차이를 지역간 비교를 통해 확인 할 수 있었다. 본 연구의 결과는 우리나라의 유용한 담수 어 자원인 붕어의 유전적 특성 규명 및 보존을 위한 구체적인 과학적 근거로서 활용 가능 할 것이라 생각된다. 또한, 앞으로 전체 미토콘드리아 DNA 염기서열을 이용하여 유전적 분석이 이루어진다면 좀 더 객관적인 유전적 유연관계 분석이 이루어 질 것으로 생각된다.
감사의 글
본 연구는 2019년 공주대학교 교내연구지원사업(과제번호:
2019-0260-01)에 의해 수행 된 것입니다.
The Conflict of Interest Statement
The authors declare that they have no conflicts of interest with the contents of this article.
References
1. Bonfield, J. K., Smith, K. F. and Staden, R. 1995. A new DNA sequence assembly program. Nucleic Acids Res. 23, 4992-4999.
2. Excoffier, L. and Lischer, H. E. 2010. Arlequin suite ver 3.5:
a new series of programs to perform population genetics analyses under Linux and Windows. Mol. Ecol. Resour. 103, 564-567.
3. Felsenstein, J. 1985. Confidence limits on phylogenies: an approach using the bootstrap. Evolution 394, 783-791.
4. Gao, Y., Wang, S. Y., Luo, J., Murphy, R. W., Du, R., Wu, S. F., Zhu, C. L., Li, Y., Poyarkov, A. D., Nguyen, S. N., Luan, P. T. and Zhang, Y. P. 2012. Quaternary palaeoenvir- onmental oscillations drove the evolution of the Eurasian Carassius auratus complex Cypriniformes, Cyprinidae. J. Bio- geogr. 39, 2264-2278.
5. Hudson, R. R., Boos, D. D. and Kaplan, N. L. 1992. A stat- istical test for detecting geographic subdivision. Mol. Biol.
Evol. 9, 138-151.
6. Jung, J., Song, K. H., Lee, E. and Kim, W. 2009. Mitochondrial genetic variations in fishes of the genus Carassius from South Korea: proximity to northern China rather than Japanese Islands? Hydrobiologia 635, 95-105.
7. Kang, J. H., Noh, E. S., Lim, J. H., Han, H. K., Kim, B. S.
and Lim, S. K. 2014. Genetic differentiation of Carassius aur- atus and C. cuvieri by the cytochrome C oxidase I gene anal- ysis. J. Aquac. Res. Develop. 53, 1-4.
8. Kim, S. Y., Song M. Y., Jeon, H. J., Kim, K. H., Lee, W. O.
and Park, H. W. 2019. Fish species composition and com- munity structure in lake Yedang, Korea. Kor. J. Ichthyol. 31, 101-109.
9. Komiyama, T., Kobayashi, H., Tateno, Y., Inoko, H., Gojobori, T. and Ikeo, K. 2009. An evolutionary origin and selection process of goldfish. Gene 430, 5-11.
10. Koressaar, T. and Remm, M. 2007 Enhancements and mod- ifications of primer design program Primer3. Bioinformatics 23, 1289-1291.
11. Kumar, S., Stecher, G. and Tamura, K. 2016 MEGA7: Molec- ular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 337, 1870-1874.
12. Librado, P. and Rozas, J. 2009. DnaSP v5: a software for comprehensive analysis of DNA polymorphism data. Bioin- formatics 25, 1451-1452.
13. Morgenstern, B. 2004. DIALIGN: multiple DNA and protein sequence alignment at BiBiServ. Nucleic Acids Res. 32, W33- W36.
14. Mori, T. and Uchida, K. 1934. A revised catalogue of the fishes of Korea. J. Chosen Nat. Hist. Soc. 19, 12-13.
15. Nam, M. M., Hong, J. Y. and Seo, B. K. 1989. Morphological variation of the Crusian Carp. Carassius auratus Linnaeus from Yongnam Area in Korea. Kor. J. Ichthyol. 11, 54-63.
16. Takada, M., Tachihara, K., Kon, T., Yamamoto, G., Iguchi, K., Miya, M. and Nishida, M. 2010. Biogeography and evo- lution of the Carassius auratus-complex in East Asia. BMC Evol. Biol. 12, 7.
17. Weir, B. S. and Cockerham, C. C. 1984 Estimating F-statistics for the analysis of population structure. Evolution 38, 1358- 1370.
18. Yoon, J. M. and Park, S. Y. 2006. Genetic comparison be- tween crucian carp Carassius auratus Linnaeus) and Crucian Carp C. cuvieri Temminck and Schlegel). J. Anim. Sci. Technol.
48, 637-650.
초록:예당호 붕어와 떡붕어의
CYTB유전자를 이용한 유연관계 분석
김계웅․조성덕․김학연․박희복*
(공주대학교 산업과학대학 동물자원학과)