pISSN 1598-642X eISSN 2234-7305
Expression of Recombinant Hybrid Peptide Gaegurin4 and LL37 using Fusion Protein in E. coli
Bayarbat, Ishvaanjil1, Jae-Hag Lee2*, and Soon-Youl Lee3*
1Department of Chemistry, School of Natural Science, Mongolian State University of Agriculture, Ulaanbaatar, Khan-Uul District, Zaisan, 17024, Mongol
2Department of Food and Nutrition, Seoil College, Seoul 131-702, Korea
3Department of Biotechnology, Genetic Engineering Research Institute, Hankyong National University, Anseong 456-749, Korea
Received : March 13, 2012 / Revised : March 30, 2012 / Accepted : April 3, 2012
Antimicrobial peptides (AMPs) are important components of living organisms acting against Gram-negative and Gram-positive bacterial and fungal pathogens. Cathelicidin human peptides have a variety of biological activities that can be used in clinical applications. AMPs are not produced naturally in large quantities, and chemical synthesis is also economically impractical, especially for long peptides. Therefore, as an alternative, heterologous expression of AMPs by recombinant techniques has been studied as a means to reduce produc- tion costs. E. coli is an excellent host for the expression of AMPs, as well as other recombinant proteins, because of the low cost involved and its easy manipulation. However, overexpression of AMPs in E. coli has been shown to cause difficulties resulting from the toxicity of the subsequently produced AMPs. Therefore, fusion expression was theorized to be a solution to this problem. In this study, AMPs were expressed as fused proteins with the glutathione S-transferase (GST) binding protein to protect against the toxicity of AMPs when expressed in E. coli. The LL37, and hybrid gaegurin and LL37 (GGN4(1–16)-LL37(17-32), which we designated as GL32, peptides were expressed as GST-fusion proteins in E. coli and the fusion proteins were then purified by affinity columns. The purified peptides were obtained by removal of GST and were con- firmed by western blot analysis. The purified antimicrobial peptides then demonstrated antimicrobial activities against Gram-negative and Gram-positive bacterial strains.
Keywords: Antimicrobial peptide, LL37, gaegurin (GGN4), fusion, heterologous expression, E. coli
Introduction
Increasing resistance of virtually all microbes toward common antibiotics that have provided safety for the last half a century is a major concern presently in the health science [17]. Recent studies have shown that antimicrobial peptides (AMPs) potentially have a role in addressing this problem [for review, 20]. AMPs are peptides and small proteins with microcidal activity. Natural antimicrobial peptides probably represent one of the very first evolved
forms of chemical defense of living eukaryotic cells against invasion by other living organisms [3, 19, 23].
Antimicrobial peptides consisted of 12 to 50 amino acids in multicellular organisms including human, animals, and plants are found on external surfaces such as the skin and the lungs or they are sequestered in granules of neutrophils, from where they can be released to kill microbes [7, 21, 23]. There is now increasing evidence that AMPs play an important role in the immune system of mammals includ- ing humans [2, 12].
Some antimicrobial peptides are synthesized constitutively, for instance the histatins [18] and human β-defensin-1 [22].
LL37 and human β-defensins are often induced in response of an infection and thus can be considered as acute-phase proteins [5, 6].
Many antimicrobial peptides display broad-spectrum
*Corresponding author S.-Y. L.
Tel: +82-31-670-5333, Fax: +82-31-670-5333 E-mail: [email protected]
J.-H. L.
Tel: +82-2-490-7509, Fax: +82-2-490-7820 E-mail: [email protected]
activity against Gram-negative, Gram-positive bacteria, and fungi [13, 20]. Several cationic amphipathic peptides also display antiviral activity in vitro. For example, defensins are able to neutralize herpes simplex virus (HSV), vesicular stomatitis virus, and influenza virus [for review, 20].
The human cationic antimicrobial protein 18 (hCAP-18), also called LL37, is only one cathelicidin antimicrobial peptide isolated from human [1]. Like other cathelicidins, it is first expressed as precursor, the 18 kDa human cationic antimicrobial protein, hCAP-18, which consists of the highly conserved N-terminal cathelin-like domain (or pro- sequence) and the C-terminal antimicrobial peptide domain [4]. The cathelin-like domain neutralizes the antimicrobial activity of its antimicrobial peptide domain. However, cleavage of the hCAP-18 in vivo gives rise to the mature antimicrobial peptide. In the last years LL37 and defensin, another prominent mammalian AMP, were shown to play not only in the innate defense mechanisms against bacteria, viruses, fungi, but also in the regulation of immune cell activation and migration [5].
Gaegurin4 (GGN4), antimicrobial peptide isolated from Rana rugosa Korean frog skin [15], is a 37 residue hydro- phobic, antimicrobial peptide [11] that shows a broad spectrum of antimicrobial activity against Gram-negative, Gram-positive bacteria, fungi and protozoa with slightly different species activities and shows weak hemolytic activities [15].
Recently, AMPs were paid attention as a clinical purpose likely due to the fact that they are less susceptible to the development of bacterial resistance because they disrupt the membrane of bacteria through non-specific peptide lipid interactions [23]. Furthermore, the action of most AMPs is extremely rapid at concentrations very close to minimal inhibitory concentrations, and several AMPs display unusually broad spectrum of activity and kill multi- drug resistant pathogens [20]. Among these antimicrobial peptides, LL37, defensins, histatin are very promising candidate because these originated from human [21]. The highly efficient production of biologically active LL37, with large quantity and low cost, is an absolute need potential clinical application. Recombinant technique can be a solution to cost problem in large production of antimicrobial peptides.
However, heterologous expression for the production of recombinant AMPs may be harmful to the host bacteria [8].
To overcome its problem, we decided to express peptides as fusion protein of glutathione S-transferase (GST) and
AMPs and then purify peptide after cleaving fusion partner and examined the microcidal activity.
Materials and Methods
Reagents and bacterial strains
Briefly, E. coli strain DH5α and BL21 (DE3) were purchased from Invitrogen (USA). B. subtilis strain KCCM 12248 was purchased from Korean Culture Center of Microorganisms (Korea). Gaegurin4 expression vector (pGEX-H6GGN4) was kindly provided by laboratory of molecular genetics, Seoul National University, in Korea [15]. LL37 peptide gene was cloned by PCR amplification based on the reported sequence in Genbank (NCBI, NM_004345). Plasmid DNA purification kit, agarose gel extraction kit and factor Xa were from QIAGEN (USA).
For the SDS-PAGE and western blot reagents, acrylamide and bisacrylamide were from Amerisco (USA). The ex- pression vector pGEX-5X-3, glutathione Sepharose 4B, desalting column, benzamidine Sepharose 4FF (high sub) column and ECL anti-mouse lgG were from GE Healthcare (USA), anti-human LL37/hCAP-18 monoclonal antibody (MAb) was from Hycult Biotechnology (Holland). General reagents were purchased from Sigma (USA).
Vector construction
The expression vector pGEX-5X3-AMPs (LL37 and tandemly fused hybrid GL32 as shown in Fig. 1) were constructed by PCR amplification of AMP genes and subcloning into pGEX-5X3 (Fig. 1). In order to prepare hybrid AMP gene, GGN4(1-16) and LL37(17-32) coding regions were tandemly fused to keep ampiphathic helix properties of the original peptides. The resulting expression vectors were introduced E. coli strain BL21(DE3) to express fusion peptides.
Expression and purification of GST-AMPs fusion proteins
Protein expression was induced with 0.1 mM IPTG and cells were harvested after 6 h of incubation at 30oC. All cells were harvested and washed with PBS and resuspended in 1× PBS buffer (pH 7.4) and 5 mM DTT. After treatment with lysozyme, the cell lysate was sonicated three times on ice. The suspension was treated with Triton X-100 to a final concentration of 1% at 4oC overnight. The GST- AMPs fusion proteins were purified using a 1 ml GSTrap
FF column as suggested by the manufacturer, GE Healthcare (USA). GST-AMPs were eluted from the column at 4oC with 5 volume of elution buffer (50 mM Tris-HCl, pH 8.0, containing 10 mM reduced glutathione and 5 mM DTT). The flow-through was collected in 0.5 ml fractions and reserved until the procedure has been successfully completed. The presence and purity of eluted GST-AMPs were evaluated by SDS-PAGE using 15% SDS gels and western blotting analysis.
Dialysis and Factor Xa cleavage of GST-AMPs fusion proteins
The solution containing GST-AMPs proteins were dialyzed against 400 volumes of Factor Xa cleavage buffer (50 mM Tris-HCl, 150 mM NaCl, and 1 mM CaCl2, pH 7.5) for overnight. For cleavage of AMPs from GST tag, the dialyzed GST-AMPs fusion protein solutions were incubated with Factor Xa cleavage buffer plus 80 U Factor Xa for 16-24 h at 21-23oC. The cleavage reaction was stopped by freezing the sample at 20oC. The enzyme was removed from the eluted protein using Benzamidine Sepharose 4 FF (high sub) column.
SDS-PAGE and western blot
Purified GST-fusion proteins or the lysates of cells with or without IPTG induction were centrifuged and the super- natants were subjected to 12-15% SDS-PAGE. Then the gels were visualized by staining with Coomassie brilliant blue R250 or by silver staining. After Xa factor digestion,
the purified recombinant LL37, GL32 proteins were analyzed by Pepti-PAGE-15% as suggested by manufac- ture. The proteins were transferred electrically onto nitro- cellulose membrane after SDS-PAGE or Pepti-PAGE for western blot analysis using commercial MAb against human LL37 (1:100) followed by ECL detection.
Assay for antimicrobial activity
The recombinants LL37 and GL32 were purified and concentrated by Centricon centrifugal filter as suggested by Millipore Corporation (USA). Antimicrobial activity was measured by determining the minimal inhibitory concen- tration (MIC) [19], which was described with some modi- fications. Briefly, we used E. coli strain DH5α (Gram- negative), B. subtilis strain KCCM 12248 and Staphyloco- ccus aureus strain Newman (Gram-positive). The colonies are diluted into a PBS solution and sterile cotton swab is used to remove an inoculum from the PBS and cover the surface of a petri-dish containing NB agar. Disk impre- gnated with various anti peptides are distributed a circular pattern on the inoculated agar surface and incubated at 37oC, overnight.
Results and Discussion
Construction of the expression vectors
Recently, human-originated AMPs, especially LL37, have been studied extensively because of its promising clinical applications [14]. The production of biologically Fig. 1. Diagram of expression vector for AMPs. (A) Expression vector, pGEX-5X3-AMPs has GST, Xa cleavage site and AMP genes inserted in the multiple cloning site. (B) α helical wheel diagram of GL32 (based Edmundson diagram). The wheel is made perfect amphi- pathic α helix with the right upper part hydrophobic and the left lower part hydrophilic: inner circle, GGN4; outer circle, LL37.
active LL37 in large amounts with reduced costs can be achieved using recombinant techniques.
Here, we have expressed peptides as fusion protein to overcome the possible toxicity of AMP when overexpressed in E. coli. We used the pGEX-5X3-GST vector system with inducible tac promoter. The expression vector were designed to express human AMP gene fused downstream GST gene by inserting the AMP genes into the multiple cloning sites of pGEX-5X3 having IPTG-inducible tac promoter. Between GST and AMP genes, there is Factor Xa site-specific rec- ognition sequences, for the release of intact AMPs (Fig. 1A).
We have chosen LL37 and hybrid peptide of LL37 and gaegurin4 (GGN4) [15]. We designed hybrid antimicrobial peptide of LL37 and gaegurin4 so that the resulting peptide keeps the amphipathic property of original peptides. Nucle- otide sequence for the peptide from amino acid 1 to amino acid 16 of gaegurin4 was connected to the nucleotide sequence for the peptide from 17 to amino acid 32 of LL37 (Fig. 1B) so that the number of the amino acids of the resulting hybrid peptide be 32. Hybrid antimicrobial peptide gene (gaegurin(1-16)-LL37(17-32), we designated as GL32) was also subcloned into expression vector to give GST-fusion hybrid protein.
Induction of fusion protein expression
To determine whether pGEX-5X3-AMPs (LL37 and GL32) could express the fusion proteins, E. coli BL21 cells harboring the plasmids were treated with 0.1 mM IPTG.
First, we examined the cell viability and cell growth rate when the fusion proteins were overexpressed. Overexpres- sion of the fusion proteins GST-LL37 and GST-GL32 did not limit the cell growth indicating that GST is able to sequester LL37 and the GST moiety completely blocked host cell toxicity resulting from antimicrobial activity.
The GST-AMP (LL37 and GL32) fusion proteins are consisted of the following regions sequentially from its N- terminus: the GST domain (consisted of 240 residues), Factor Xa cleavage site (4 residues), and the AMPs domain (37 residues). Thus, the fusion protein would have 280 amino acid residues which correspond to the molecular weight of about 30 kDa. Indeed, the expression of an approximately 30 kDa protein was induced in the presence of IPTG for both GST-LL37 (Fig. 2A) and GST-GL32 (Fig.
2B).
The amount of the expressed fusion protein was esti- mated to be 15~30% of the total protein in E. coli BL21
harboring pGEX-5X3-LL37 plasmid as judged from Coo- massie Blue staining (data not shown).
Purification of GST-AMPs
GST-AMP (LL37 and GL32) fusion proteins were purified from the supernatants of cell lysates using a 1 ml GSTrap FF (GE Healthcare, USA) column to be >95%
purity (Fig. 2). Fig. 2 shows the fusion proteins at the different steps of purification, for example, cell lysate, throw flow, wash and elution state. Intense bands corres- ponding to the molecular weights of the expected proteins were shown: an approximately 26 kDa protein for GST and an approximately 30 kDa protein for GST-AMPs. The presence of a band at 26 kDa may be caused by intracellular cleavage of the fusion protein, or by translational pausing at the junction between GST and the fusion partner.
Excessive sonication in the step of cell lysate preparation may also lead to denaturation and breakdown of the fusion protein, which results in the appearance of a band at 26 kDa. The recombinant GST-AMP fusion proteins expressed in this condition were from soluble fractions, not from inclusion bodies.
Production of recombinant LL37 (rLL37) and GL32 (rGL32)
The recombinant LL37 and GL32 peptides can be harvested from the GST-fusion proteins by the cleavage with Factor Xa to remove GST. The fusion protein was digested with an appropriate site specific protease Xa (Ile- Fig. 2. SDS-PAGE of recombinant GST-LL37 and GST-GL32 fusion protein purified at different stage. A, GST- LL37 and B, GST-GL32 were purified from E. coli BL-21 using a glutathione affinity chromatography. CL, cell lysate that is the extract of IPTG induced E. coli BL-21 (DE3) containing the expression vector pGEX-5X3. TF, throw flow; W, wash; E, elution. The overex- pressed recombinant GST-LL37 (approximately 30 kDa), GST- GL32 (approximately 34 kDa) fusion proteins were analyzed by 15% SDS-PAGE.
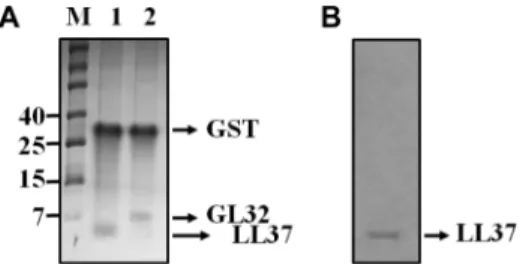
Glu-Gly-Arg) while the fusion protein is bound to Glu- tathione Sepharose 4B (GE Healthcare, USA). Cleavage of the bound fusion protein eliminated the extra step of separating the released protein from GST. Fig. 3A shows the separation of GST and AMPs when stained with silver staining.
Western blot analysis showed that one single band corresponding to approximately 4.5 kDa, the same size as the chemically synthesized LL37 was observed with the western blot analysis (Fig. 3B).
In vitro antimicrobial activity assay of recombinant LL37 and GL32
The recombinant LL37 (rLL37) and GL32 (rGL32) were purified and were concentrated by Centricon (Amicon bioseparation, USA) as suggested by manufacturer. And the inhibitory effects of recombinant rLL37 and rGL32 were tested against three bacteria, E. coli, B. subtilis and Staphylococcus aureus by Agar disk diffusion test method (Fig. 4A and 4B). Both the recombinant rLL37 and rGL32
like chemically-synthesized LL37 showed clear zone of in- hibition against E. coli, B. subtilis, and S. aureus suggesting rLL37 and rGL32 has the antimicrobial activity as expected. rLL37 and rGL32 showed better activity against Gram-positive bacteria for both rLL37 and rGL32 even though they showed not much different antibacterial activity for Gram-negative and Gram-positive bacteria.
Compared to rLL37, rGL32 showed lower activity because of unknown reason. One possible reason could be the less perfect amphipathicity of the resulting hybrid peptide compared to that of the original LL37. These recombinant AMPs also showed dose dependent activity against B.
subtilis as shown in Fig. 4C.
In summary, we showed the expression of antimicrobial peptide as fusion GST-AMP with inducible promoter and the antimicrobial activity of the resulting recombinant AMP against Gram-negative and Gram-positive bacteria.
In other words, this technique can be used as the produc- tion of the active recombinant AMPs. This result supports the recent report to express the AMPs in E. coli by express- ing as fusion protein to protect the possible toxicity of AMPs when in expressed [9, 10, 16]. For example, AMPs were heterologously expressed in E. coli as fusion protein with Trx [9, 10] and as fusion protein with carbonate binding protein in order to express AMP using recombinant technique [16]. Also we have succeeded the expression of the active hybrid AMP because we designed the hybrid gene so that they keep amphipathic helix properties of the original peptides.
REFERENCES
1. Andersson, E., O. E. Sorensen, B. Frohm, N. Borregaard, A.
Egesten, and J. Malm. 2002. Isolation of human cationic antimicrobial protein-18 from seminal plasma and its associ- ation with prostasomes. Hum. Reprod. 10: 2529-2534.
2. Bals, R. 2000. Epithelial antimicrobial peptides in host defense against infection. Respir. Res. 1: 141-150.
3. Boman, H. G., I. Faye, G. H. Gudmundsson, J. Y. Lee, and D.
A. Lid-holm. 1991. Cell free immunity in Cecropia. A model system for antibacterial proteins. Eur. J. Biochem. 210: 23- 31.
4. Boman, H. G. 2003. Antibacterial peptides: basic facts and emerging concepts. J. Intern. Med. 254:197-215.
5. Frasca, L. and R. Lande. 2012. Role of defensins and cathe- licidin LL37 in auto-immune and auto-inflammatory dis- eases. Curr. Pharm. Biotechnol. [Epub ahead of print].
6. Frohm, M., B. Agerberth, G. Ahangari, M. Stahle-Backdahl, S. Liden, H. Wigzell, and G. H. Gudmundsson. 1997. The Fig. 3. SDS-PAGE of recombinant LL37 (rLL37) and GL32
(rGL32) and western blot analysis of rLL37. A, Lane M, pro- tein size marker (kDa); Lane 1 and 2, rLL37 or rGL32 with GST digested with Factor Xa and analyzed by Pepti-Gel 15%. B, west- ern blot analysis of the purified rLL37.
Fig. 4. Antimicrobial activity of purified recombinant LL37 (rLL37) and GL32 (rGL32) in vitro. A, antimicrobial activity of chemically synthesized LL37 and recombinant rLL37 against 1, E.
coli; 2, B. subtilis and 3, Staphylococcus aureus. B, antimicrobial activity of recombinant GL32 against 1, E. coli; 2, B. subtilis, 3, S.
aureus and C, dose dependent antibacterial activity of rLL37 and rGL32 against B. subtilis with relative amount of the purified AMPs.
expression of the gene coding for the antibacterial peptide LL37 is induced in human keratinocytes during inflamma- tory disorders. J. Biol. Chem. 272: 15258-15263.
7. Harder, J. J., Bartels, E., Christophers, and J. M. Schroder.
1997. A peptide antibiotic from human skin. Nature 387:
861-862.
8. Ingham, A. B. and R. J. Moore. 2007. Recombinant produc- tion of antimicrobial peptides in heterologous microbial sys- tems. Biotechnol. Appl. Biochem. 47: 1-9.
9. Jing, X. L., X. G. Luo, W. J. Tian, L. H. Lv, Y. Jiang, N.
Wang, and T. C. Zhang. 2010. High-level expression of the antimicrobial peptide plectasin in Escherichia coli. Curr.
Microbiol. 61: 197-202.
10. Krahulec, J, M. Hyrsová, S. Pepeliaev, J. Jílková, Z. Cerný, and J. Machálková. 2010. High level expression and purifi- cation of antimicrobial human cathelicidin LL-37 in Escher- ichia coli. Appl. Microbiol. Biotechnol. 88: 167-175.
11. Kwon, S. Y., B. A. Carlson, J. M. Park, and B. J. Lee. 2000.
Structural organization and expression of the gaegurin 4 gene of Rana rugosa. Biochim. Biophys. Acta. 1492: 185-190.
12. Lai, Y. and R. L. Gallo. 2009. AMPed up immunity: how antimicrobial peptides have multiple roles in immune defense. Trends. Immunol. 30: 131-141.
13. Miyasaki, K. T. and R. I. Lehrer. 1998. β-sheet antibiotic peptides as potential dental therapeutics. Int. J. Antimicrob.
Agents. 9: 269-280.
14. Nijnik, A. and R. E. Hancock. 2009. The roles of cathelici- din LL-37 in immune defences and novel clinical applica- tions. Curr. Opin. Hematol. 16:41-47.
15. Park, J. M., J. E. Jung, and B. J. Lee. 1994. Antimicrobial peptides from the skin of a Korean frog, Rana Rugosa. Bio- chem. Biophy. Res. Commun. 205: 948-954.
16. Ramos, R., L. Domingues, and M. Gama. 2010. Escherichia coli expression and purification of LL37 fused to a family III carbohydrate-binding module from Clostridium thermocel- lum. Protein Expr. Purif. 71: 1-7.
17. Smith, R. D. and J. Coast. 2002. Antimicrobial resistance: a global response. Bull. World Health Organ. 80: 126-133.
18. Tsai, H. and L. A. Bobek. 1998. Human salivary histatins:
promising anti-fungal therapeutic agents. Crit. Rev. Oral Biol. Med. 9: 480-497.
19. Turner, J., Y. Cho, N. N. Dinh, A. J. Waring, and R. I.
Lehrer. 1998. Activities of LL-37, a cathelin-associated anti- microbial peptide of human neutrophils. Antimicrob. Agents.
Chemother. 42: 2206-2214.
20. Wiesner, J. and A. Vilcinskas. 2010. Antimicrobial peptides, The ancient arm of the human immune system. Virulence 1:
440-464.
21. Wilmes, M., B. P. Cammue, H. G. Sahl, and K. Thevissen.
2011. Antibiotic activities of host defense peptides: more to it than lipid bilayer perturbation. Nat. Prod. Rep. 28: 1350- 1358.
22. Yang, Y. H., G. G. Zheng, G. Li, X. J. Zhang, Z. Y. Cao, R.
Qing, and K. F. Wu. 2004. Expression of bioactive recombi- nant GSLL-39, a variant of human antimicrobial peptide LL37, in E.coli. Protein Expr. Purif. 37: 229-235.
23. Zasloff, M. 2002. Antimicrobial peptides of muticellular organisms. Nature 415: 389-395.
국문초록
Glutathione S-Transferase에 융합한 재조합 Hybrid Peptide Gaegurin-LL37의 대장균에서의 발현 바야르바트 이쉬반질1·이재학2·이순열3*
1몽골농과대학, 화학공학과, 2서일대학교 식품영양과, 3한경대학교 생명공학과
항균 펩타이드(Antimicrobial peptides(AMPs)는 그람 양성, 그람 음성 세균과 진균병원체에 대항하는 생명체에서 중요한 역할을 하는 물질이다. 인간의 Cathelicidin 항균 펩타이드는 임상학적으로 사용할 수 있는 여러 가지의 생물 학적 활성을 가진다. 항균 펩타이드의 생산 비용은 재조합 방법으로 낮출 수가 있다. 대장균은 저렴하며 손쉬운 조 작이 가능하기 때문에 다른 재조합 단백질처럼 항균 펩타이드의 발현에 훌륭한 숙주가 될 수 있다. 그러나 대장균에 서의 항균 펩타이드의 과발현은 항균 펩타이드가 과발현 되었을 때 대장균에 독성을 보일 수 있으므로 어려움이 보 고가 되어있다. 본 연구에서는 이러한 문제점을 극복하고자 항균 펩타이드를 Glutathione S-transferase(GST) 결합 단 백질에 융합하여 항균펩타이드의 독성을 감소시키도록 설계하여 발현을 시도하였다. 이 때 발현한 항균 펩티드는 LL37과, gaegurin4과 LL37의 잡종 펩타이드 GGN4-LL37(GL32로 명명)를 GST에 융합되도록 벡터를 구축하고 설계 하여 대장균에서 GST 융합단백질로 발현시켰다. 융합 단백질은 친화력 컬럼을 사용하여 분리하고 GST를 절단하여 항균펩타이드 만을 분리하였고 분리한 펩타이드는 웨스턴 블롯팅으로 확인하였고 그람 양성, 그람 음성 세균에 대하 여 항균 활성을 나타내는 것을 확인하였다.