80 책임저자:이원섭
660-702, 진주시 칠암동 90 경상대학교 의과대학 내과학교실 Tel: 055-750-8733, Fax: 055-758-9122
E-mail: [email protected] or [email protected] 정진명
660-702, 진주시 칠암동 90 경상대학교 의과대학 신경외과학교실 Tel: 055-750-8110 or Fax: 055-758-9122 E-mail: [email protected]
접수일:2011년 2월 1일, 1차 수정일:2011년 2월 10일, 2차 수정일:2011년 2월 15일, 게재승인일:2011년 2월 18일
Correspondence to:Won Sup Lee
Department of Internal Medicine, Gyeongsang National University School of Medicine, 90, Chilam-dong, Jinju 660-702, Korea
Tel: +82-55-750-8733, Fax: +82-55-758-9122 E-mail: [email protected] or [email protected] Co-Corresponding Author:Jin-Myung Jung
Department of Neurosurgery, Gyeongsang National University School of Medicine, 90, Chilam-dong, Jinju 660-702, Korea
Tel: +82-55-750-8110, Fax: +82-55-758-9122 E-mail: [email protected]
Anthocyanins Isolated from Vitis coignetiae Pulliat Suppress NF-κB-regulated Genes Involved in Cell proliferation, Anti-apoptosis, Invasion and Angiogenesis
Min Jeong Kim1, Won Sup Lee1, Jing Nan Lu1, Jeong Won Yun1, Seong-Hwan Chang2, Yung Hyun Choi3, Jin-Myung Jung4, Chung Ho Ryu5, Ock Jin Park6, Soon Chan Hong7 and Sung Chul Shin8 Departments of 1Internal Medicine, 4Neurosurgery and 7Surgery, Institute of Health Sciences, Gyeongsang National
University School of Medicine, Jinju 660-702, 2Department of Surgery, Konkuk University School of Medicine, Seoul 143-701, 3Department of Biochemistry, Dongeui University College of Oriental Medicine and Department of Biomaterial Control (BK21 Program), Dongeui University Graduate School, Busan 614-052,
5Division of Applied Life Science (BK21 Program) and Institute of Agriculture and Life Science, and
8Chemistry, Research Institute of Life Science, Gyeongsang National University, Jinju 660-701,
6Department of Food and Nutrition, Hannam University, Daejeon 305-811, Korea
NF-κB activation is closed related to the cancer cell proliferation, invasion, angiogenesis, metastasis, and drug resistance. We previously suggested that anthocyanins from fruits of Vitis coignetiae Pulliat (AIMs), which had been used as a remedy for inflammatory diseases and cancers, should have anti-cancer properties. We tried to validate their effects on NF-κB-regulated gene products in Hep3B human hepatoma cells. The AIMs inhibited the cell proliferation of Hep3B cells in a dose-dependent manner.
To clearly demonstrate their effects, we used TNF-α which is an activator of NF-κB. AIMs suppressed the NF-κB-regulated proteins (Cyclin D1, COX-2, C-myc, Bcl-2, Bcl-xL, XIAP, cIAP1, cIAP2, MMP-2, MMP-9, ICAM, and VEGF) induced by TNF-α. In summary, this study indicates that the AIMs have inhibitory effects on NF-κB downstream proteins involved in cell proliferation, anti-apoptosis, cell invasion, and angiogenesis. This study provides the evidence that the anthocyanins isolated from Vitis coignetiae Pulliat might be useful in the treatment of human hepatocellular carcinoma. (Cancer Prev Res 16, 80-85, 2011)
Key Words: Anthocyanin, IMs, Hep3B, TNF-α, NF-κB
INTRODUCTION
The incidence of cancer and cancer-related mortality in- creases with age. These elderly patients have a higher incidence of serious side effects from conventional chemotherapeutic
agents. For the management of these patients, new less toxic drug development is essential. Recently dietary agents have been reported to safely modulate physiological function and en- hance anti-cancer activity.1,2) Most of cancer patients eventually die of metastasis. Thus the reduction in chances of metastasis can be an alternative strategy to conquer cancer. To develop
metastasis, the cancer cells have to evade immune surveillance, invade tissue and overcome apoptotic stimuli. In addition, after the cells reach blood stream they need the abilities of adhesion, proliferation, and angiogenesis. Nuclear factor-κB (NF-κB) regulates these genes involved in cancer cell invasion, an- ti-apoptosis, adhesion, proliferation, and angiogenesis.3,4) Vitis coignetiae Pulliat (Known as Meoru in Korea) belongs to the grape family, which has traditionally been used in Korean folk medicine for the treatment of inflammatory dis- orders and cancer. The intense dark red hue of the fruit is re- flecting an abundance of anthocyanin pigments. The anthocya- nins have been reported to have anti-angiogenic and anti-in- vasive properties.5,6) We previously the anthocyanins isolated from Vitis coignetiae Pulliat have anti-cancer activities.7) Here, we focused the effects of AIMs on NF-κB-regulated proteins involved in cancer cell proliferation, invasion, anti-apoptosis, and angiogenesis in human hepatocellular carcinoma cells.
MATERIALS AND METHODS 1. Cell culture and chemicals
Hep3B human hepatocellular carcinoma cells from the ATCC (Rockville, MD) were cultured in RPMI 1640 medium (Invitrogen Corp, Carlsbad, CA) supplemented with 10% FBS (GIBCO BRL, Grand Island, NY), 100 U/ml penicillin, and 100μg/ml streptomycin in an incubator at 37oC in a humidi- fied atmosphere of 95% air and 5% CO2. Molecular mass markers for proteins were obtained from Pharmacia Biotech (Saclay, France). For Western blotting, antibodies against Bcl-2, Bcl-xL, X-linked inhibitor of apoptosis protein (XIAP), cellular inhibitor of apoptosis protein-1 (cIAP-1), cIAP-2, COX-2, cyclin D1, c-Myc, matrix metalloproteinase-2 (MMP- 2), MMP-9, VEGF, intercellular adhesion molecule-1 (ICAM-1) were purchased form Santa Cruz Biotechnology (Santa Cruz, CA, USA). Antibody against β-actin was from Sigma (Beverly, MA). Peroxidase-labeled donkey anti-rabbit and sheep an- ti-mouse immunoglobulin, and an enhanced chemiluminescence (ECL) kit were purchased from Amersham (Arlington Heights, IL). All other chemicals not specifically mentioned were pur- chased from Sigma Chemical Co. (St. Louis, MO). The compo- sition of the anthocyanins isolated from Vitis coignetiae Pulliat (known as Meoru in Korea) is as follows: delphinidin-3,5-diglu- coside (1): cyanidin-3,5-diglucoside (2): petunidin-3,5-digluco- side (3): delphinidin-3-glucoside (4): malvdin-3,5-diglucoside
(5): peonidin-3,5-diglucoside(6): cyanidin-3-glucoside (7): petu- nidin-3-glucoside (8): peonidin-3-glucoside (9): malvidin-3-glu- coside (10) = 3.5: 3.4: 7.1: 23.9: 8.0: 9.6: 9.1: 16.1:5.7: 13.4.
2. Cell proliferation assays
For the cell viability assay, the cells were seeded onto 24-well plates at a concentration of 5×104 cells/ml, grown to 70% confluence and then treated with the indicated concen- trations of the anthocyanins isolated from Vitis coignetiae Pulliat (AIMs) for the indicated times. Control cells were supple- mented with media containing 0.1% DMSO (vehicle control).
Cell viability was determined by trypan blue-exclusion methods.
3. Cell invasion assay
For the cell invasion assays, Hep3B cells were cultured in serum-free media overnight. 5×104 cells were loaded onto pre-coated Matrigel 24-well invasion chambers (BD Bioscien- ces, San Jose, CA) with the indicated treatment, and 0.5 ml of medium containing 20% FBS serving as a chemoattractant was added to the wells of the plate. The Matrigel invasion chambers were incubated for 18 h, and the cells on the Matrigel were removed. Invading cells were fixed with 10%
formalin, stained with DAPI, and counted.
4. Western blotting
The concentrations of the proteins of cell lysate and of xeno- graft tumor samples were determined by means of the Bradford protein assay (Biorad lab, Ricmond, CA, USA) using bovine serum albumin as the standard. The proteins of the extracts were resolved by electrophoresis, electrotransferred to a poly- vinylidene difluoride membrane (Millipore, Bedford, MA), and then the membrane was incubated with the primary antibodies followed by a conjugated secondary antibody to peroxidase.
Blots were developed with an ECL detection system.
5. Statistics
Each experiment was performed in triplicate. The results were expressed as means±SD. Significant differences were de- termined using the Student's t-test. Statistical significance was defined as p<0.05.
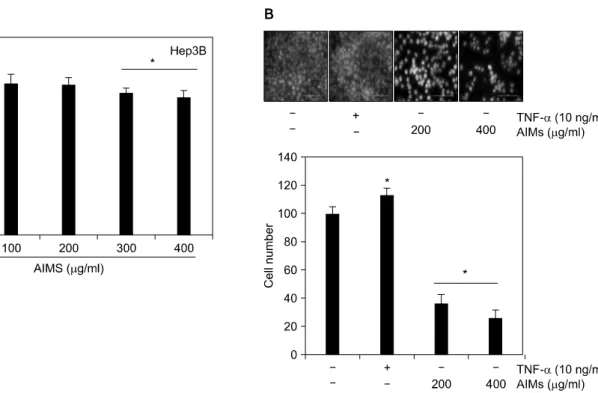
Fig. 1. Dose-dependent antiproliferative effects of AIMs on Hep3B human hepatocellular carcinoma cells. (A) Cell proliferation was assessed by trypan blue-exclusion methods. Hep3B cells were seeded at the density of 5×104 cells per ml. The cells were treated with indicated concentrations of AIMs for 24 h. (B) The cells were exposed to AIMs for 6 h. The cells (5×104 cells) were loaded on pre-coated Matrigel 24-well invasion chambers (BD Biosciences) in the presence of indicated agents (TNF-α and AIMs).
Medium containing 20% FBS was placed in the basolateral chamber to act as a chemoattractant. After 18 h, the cells on the apical side were wiped off. Next, the cells on the bottom of the filter were stained with DAPI solution and then counted. Values represent means±SD from three independent experiments. *p<0.05 versus control.
RESULTS
1. AIMs suppress the growth of Hep3B cells in a dose-dependent manner
We first examined its effects on the growth of Hep3B cells at the indicated concentration range (100∼400μg/ml). The growth of Hep3B cells was inhibited by AIMs in a dose-de- pendent manner and the inhibitory effects of AIMs at the con- centrations up to 200μg/ml were minimal (Fig. 1A).
2. AIMs suppress the invasion of Hep3B cells
Cancer cell invasion is the key events in metastasis. TNF-α is usually increased in patients with advanced cancers and the pathophysiological relevance of TNF-α-induced NF-κB acti- vation is well-known. Here, we used TNF-α, one of the most potent NF-κB stimulants to know whether it stimulate cell invasion. We found that TNF-α stimulated the invasion of Hep3B cells and AIMs inhibited Hep3B cell invasion in a dose-
dependent manner (Fig. 1B).
3. AIMs suppress the expression of TNF-α- induced NF-κB-regulated proteins which are implicated in cancer cell proliferation
Cyclin D1 is one of cell proliferation marker because it is required for cells to advance from the G1 phase to the S phase of the cell cycle. More than 30% of the cancers are known to overexpress cyclin D1.8) COX-2 has been implicated in carci- nogenic processes, and its overexpression in malignant cells has been shown to mediate cancer cell proliferation.9) c-Myc is also involved in cancer cell proliferation.10) To evaluate the in- hibitory effects on the NF-κB-regulated proteins involved in cell proliferation, we assessed the expressions of cyclin D1, COX-2, and c-Myc in Hep3B cells. Here we also used TNF-α to clearly demonstrate the AIM effects on NF-κB-regulated genes. Western blotting (Fig. 2) revealed that TNF-α stimu- lated these gene expressions, and AIMs inhibited these acti- vated gene expressions.
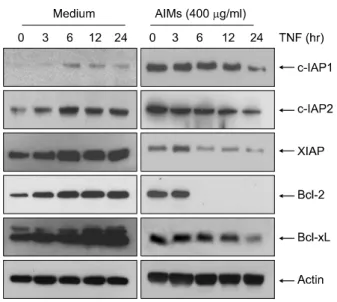
Fig. 2. AIMs suppressed TNF-α-induced NF-κB-regulated proteins involved in cancer proliferation. Hep3B cells, either left untreated or pretreated with AIMs (400 μg/ml) for 24 h and then were exposed to TNF-α (10 nM) for the indicated times.
Whole-cell extracts were prepared, and 30 μg of the whole- cell lysate was analyzed by Western blot using antibodies against COX-2, c-Myc, and cyclin D1. β-Actin was used as an internal control. The results are from one representative experiment of at least two independent experiments that showed similar patterns.
Fig. 3. AIMs suppressed TNF-α-induced NF-κB-regulated proteins involved in cancer anti-apoptosis. Hep3B cells, either left untreated or pretreated with AIMs (400 μg/ml) for 24 h and then were exposed to TNF-α (10 nM) for the indicated times. Whole-cell extracts were prepared, and 30 μg of the whole-cell lysate was analyzed by Western blot using antibodies against c-IAP1 &2, XIAP, Bcl-2, and Bcl-xL. β-Actin was used as an internal control. The results are from one representative experiment of at least two independent experi- ments that showed similar patterns.
Fig. 4. AIMs suppressed TNF-α-induced NF-κB-regulated proteins involved in cancer invasion and angiogenesis. Hep3B cells, either left untreated or pretreated with AIMs (400 μg/ml) for 24 h and then were exposed to TNF-α (10 nM) for the indicated times. Whole-cell extracts were prepared, and 30 μg of the whole-cell lysate was analyzed by Western blot using antibodies against MMP-2, MMP-9, ICAM-1, and VEGF. β- Actin was used as an internal control. The results are from one representative experiment of at least two independent experi- ments that showed similar patterns.
4. AIMs suppress the expressions of TNF-α- induced NF-κB-regulated proteins which are implicated in anti-apoptosis
NF-κB regulates the expression of several anti-apoptotic gene products, including Bcl-2, Bcl-xL, XIAP, cIAP1, and cIAP2, the overexpressions of which have been linked to cell survival, chemoresistance, and radioresistance.11,12) Moreover, Bcl-2 is one of the most important anti-apoptotic molecule, and most cancers generally overexpress Bcl-2 thereby over- coming apoptotic stimuli and escaping apoptosis. Whether AIMs modulated the TNF-induced expression of NF-κB-regu- lated anti-apoptotic gene products was examined. Western blotting (Fig. 3) revealed that TNF-α stimulated these gene expressions, and AIMs inhibited these activated gene expres- sions.
5. AIMs suppress TNF-α-induced NF-κB-re- gulated proteins which are involved in inva- sion & angiogenesis
MMPs plays a crucial role in cancer invasion and angio- genesis by mediating degradation of the extracellular matrix (ECM), and inhibition of MMP activity has been shown to sup- press lung metastasis.13,14) ICAM-1, and VEGF are involved in
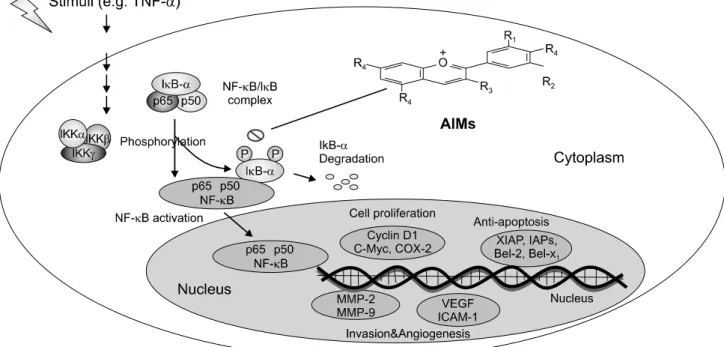
Fig. 5. Schematic representation on anticancer effects of AIMs on human hepatocellular carcinoma cells. TNF-α stimulated the NF-κB-regulated proteins involved in cancer cell proliferation (cyclin D1, c-Myc and COX-2), anti-apoptosis (XIAP, IAP1, IAP2, and Bcl-xL), invasion (MMP-2, and MMP-9), and angiogenesis (ICAM-1, and VEGF).
angiogenesis and regulated by NF-κB.
To evaluate the anti-invasive effects and anti-angiogenic ef- fects at the molecular level, we assessed the expressions of MMP-2, MMP-9, ICAM-1, and VEGF in Hep3B cells. We ob- served that TNF-α stimulated these gene expressions, and AIMs inhibited these activated gene expressions (Fig. 4).
DISCUSSION AND CONCLUSION
We have previously reported that anticancer activity of AIMs in Hep3B cell human hepatocelluar carcinoma cells. This study was designed to demonstrate the inhibitory effects on NF-κB- regulated gene products which are involved in cancer cell pro- liferation, anti-apoptosis, invasion and angiogenesis. This study suggests that AIMs have anti-cancer activities in Hep3B cells through the suppression of NF-κB-regulated proteins involved in cancer cell proliferation (cyclin D1, c-Myc, and COX-2), an- ti-apoptosis (IAP1, IAP2 XIAP, Bcl-2, and Bcl-xL), and in- vasion & angiogenesis (MMP-2, MMP-9, ICAM-1, and VEGF).
Cyclin D1 and COX-2 are overexpressed in a variety of cancers and mediates cancer cell proliferation8,9) and c-Myc is also in- volved in cancer cell proliferation.10) MMP-2, and MMP-9 are
important molecules in cancer invasion which is the first step to metastasis. ICAM-1 and VEGF is presumed to affect neo- vasculalization of cancer. All these genes are regulated by NF- κB activity.3)
It has been shown that activation of NF-κB induces resist- ance to apoptosis induced by various chemotherapeutic agents.3,11) Other studies suggest that activation of NF-κB signaling in- duces inflammation and leads to carcinogenesis.4) We pre- viously demonstrated that AIMs had inhibitory effects on NF- κB which were related to inhibition of IκB phosphorylation.7) Here we demonstrated that effective inhibition of NF-κB- regulated proteins involved in cell proliferation, anti-apoptosis, cell invasion, and angiogenesis.
We here used TNF-α, one of the NF-κB stimulants to clearly demonstrate the AIM effects on NF-κB-regulated pro- teins which are closely related to metastasis.3,15) TNF-α-induced NF-κB activation is pathophysiologically relevant to cancer in human body because TNF-α is usually increased in patients with advanced cancers16) and the advanced cancers have higher chance to metastasize.
Data presented in this study indicated that AIMs in the con- centration of 400μg/ml had the anti-anticancer effects on
Hep3B human hepatocellualr carcinoma cells. The concen- trations used in the present study are consistent with those in many other studies on the anti-tumor effect of anthocyanins on cultured cells.17,18)
In conclusion, this study showed that AIMs have inhibitory effects on TNF-α-induced NF-κB-regulated gene activation (Fig. 5). This study provides evidence that AIMs might have anticancer effects on human hepatocellular carcinoma.
ACKNOWLEDGEMENT
This study was supported by a grant of the Korea Health- care technology R&D Project, Ministry for Health, Welfare &
Family Affairs, Republic of Korea (A080164) and in part Korean Cancer Research Institute.
REFERENCES
1) Chun KH, Kosmeder JW 2nd, Sun S, Pezzuto JM, Lotan R, Hong WK, Lee HY. Effects of deguelin on the phosphati- dylinositol 3-kinase/Akt pathway and apoptosis in premalig- nant human bronchial epithelial cells. J Natl Cancer Inst 95, 291-302, 2003.
2) Hatcher H, Planalp R, Cho J, Torti FM, Torti SV. Curcumin:
from ancient medicine to current clinical trials. Cell Mol Life Sci 65, 1631-1652, 2008.
3) Aggarwal BB. Nuclear factor-kappaB: the enemy within.
Cancer Cell 6, 203-208, 2004.
4) Gilmore TD. Introduction to NF-kappaB: players, pathways, perspectives. Oncogene 25, 6680-6684, 2006.
5) Ding M, Feng R, Wang SY, Bowman L, Lu Y, Qian Y, Castranova V, Jiang BH, Shi X. Cyanidin-3-glucoside, a natural product derived from blackberry, exhibits chemopre- ventive and chemotherapeutic activity. J Biol Chem 281, 17359-17368, 2006.
6) Favot L, Martin S, Keravis T, Andriantsitohaina R, Lugnier C. Involvement of cyclin-dependent pathway in the inhibitory effect of delphinidin on angiogenesis. Cardiovasc Res 59, 479- 487, 2003.
7) Shin DY, Lee WS, Kim SH, Kim MJ, Yun JW, Lu JN, Lee SJ, Tsoy I, Kim HJ, Ryu CH, Kim GY, Kang HS, Shin SC, Choi YH. Anti-invasive activity of anthocyanins isolated from Vitis coignetiae in human hepatocarcinoma cells. J Med Food 12, 967-972, 2009.
8) Guttridge DC, Albanese C, Reuther JY, Pestell RG, Baldwin AS. NF-kappaB controls cell growth and differentiation through transcriptional regulation of cyclin D1. Mol Cell Biol 19, 5785-5799, 1999.
9) Chun KS, Surh YJ. Signal transduction pathways regulating cyclooxygenase-2 expression: potential molecular targets for chemoprevention. Biochem Pharmacol 68, 1089-1100, 2004.
10) Schmidt EV. The role of c-myc in regulation of translation initiation. Oncogene 23, 3217-3221, 2004.
11) Catz SD, Johnson JL. Transcriptional regulation of bcl-2 by nuclear factor kappaB and its significance in prostate cancer.
Oncogene 20, 7342-7351, 2001.
12) Chu ZL, McKinsey TA, Liu L, Gentry JJ, Malim MH, Ballard DW. Suppression of tumor necrosis factorinduced cell death by inhibitor of apoptosis c-IAP2 is under NF-κB control.
Proc Natl Acad Sci 94, 10057-10062, 1997.
13) Vihinen P, Kahari VM. Matrix metalloproteinases in cancer:
prognostic markers and therapeutic targets. Int J Cancer 99, 157-166, 2002.
14) Garg A, Aggarwal BB. Nuclear transcription factor- kappaB as a target for cancer drug development. Leukemia 16, 1053- 1068, 2002.
15) Bogenrieder T, Herlyn M. Axis of evil: molecular mechanisms of cancer metastasis. Oncogene 22, 6524-6536, 2003.
16) Garbisa S, Scagliotti G, Masiero L, Di Francesco C, Caenazzo C, Onisto M, Micela M, Stetler-Stevenson WG, Liotta LA.
Correlation of serum metalloproteinase levels with lung cancer metastasis and response to therapy. Cancer Res 52, 4548-4549, 1992.
17) Chen PN, Kuo WH, Chiang CL, Chiou HL, Hsieh YS, Chu SC. Black rice anthocyanins inhibit cancer cells invasion via repressions of MMPs and u-PA expression. Chem Biol Interact 163, 218-229, 2006.
18) Meiers S, Kemeny M, Weyand U, Gastpar R, von Angerer E, Marko D. The anthocyanidins cyanidin and delphinidin are potent inhibitors of the epidermal growth-factor receptor. J Agric Food Chem 49, 958-962, 2001.