The development of di/trimeric biosimilar Humira single-chain Fv antibody fused to leucine zipper or isoleucine zipper§
13
0
0
전체 글
(2) 194. ∙ Kim et al.. VH), 경사슬 가변영역(light chain variable region: VL)으로 표. and Plückthun, 1988; Reiter et al., 1995; Li et al., 2015). 시된다(Supplementary data Fig. S1). 반면에 항원과 직접적으. (Supplementary data Fig. S1). 개발된 ScFv 항체 단편들은 Fc. 로 결합에 관계하지 않는 정상 영역은 scaffold로 작용하여 가. 에 의한 비특이적 결합을 제거하였으며 자연 항체의 1/6인 크. 변 영역의 구조를 안정화시켜 가변 영역의 항원 결합력을 극대. 기인 약 25 kDa의 작은 크기이므로(Supplementary data Fig.. 화시킬 뿐만 아니라 정상 영역 중 Fc는 보체 활성화(complement. S1) 높은 표적 침투성을 나타내었다. ScFv 항체 단편들은 대장. activation) 및 식세포 작용(phagocytosis) 등의 반응기(effector). 균을 이용하여 발현시키기 때문에 단시간 내에 대량으로 저가. 기능을 담당한다(Supplementary data Fig. S1).. 생산이 가능할 뿐만 아니라 다양한 유전자 조작이 용이하여. 항원 특이성을 갖는 항체는 표적 치료가 가능하여 비특이. 다른 기능을 갖는 단백질 및 도메인(domain)을 항체 단편의 아. 적 반응을 최소화할 수 있으므로 치료용 항체로 활용될 수 있. 미노 말단 또는 카르복시 말단에 융합시킨 다기능성 ScFv. 어 암, 자가 면역 질환 및 다양한 염증 질환의 표적 치료제로 개. (multifunctional ScFv)를 제조할 수 있게 되었다. 예를 들어. 발되었다(Brekke and Løset, 2003; Pavlou and Belsey, 2005;. ScFv는 monomer이므로 1개의 항원 결합 부위를 구성하여 2. Li et al., 2018). 특히 종양괴사인자-알파(Tumor necrosis factor-. 개의 결합 부위를 갖는 자연 항체(Supplementary data Fig. S1). α: TNF-α)의 과 발현에 의해 발생하는 류마티스 관절염(Rheu-. 보다 항원 결합력이 감소될 수 있으므로 di/trimerization시킬. matoid arthritis)을 치료하기 위한 항체치료제가 개발되었다. 수 있는 도메인을 추가하여 2개 이상의 항원 결합 부위를 갖는. (Bondeson and Maini, 2001; Rau, 2002; Carmona et al., 2005;. di/trimeric ScFv를 제조하였다(Pack et al., 1993; De Kruif and. Tracey et al., 2008). 즉 TNF-α는 염증반응에 관계하는 사이토. Logtenberg, 1996; Shiraishi et al., 2004). 또한 효소 및 독소 등. 카인(cytokine)으로 관절 및 연골조직 등에 풍부하게 존재하. 을 항체 단편에 융합하여 발현시키게 되면 융합된 효소 및 독. 는 혈관 내피 세포(endothelial cell) 및 섬유아세포(fibroblast). 소의 기능과 항체 단편의 항원 결합 기능을 동시에 갖는 융합. 표면에 있는 TNF-α 수용체와 결합하여 염증을 일으킨다(Rau,. 항체(bifunctional ScFv)도 제조할 수 있게 되었다(Brinkmann. 2002). 그러므로 TNF-α에 결합하여 TNF-α 수용체와 결합을. et al., 1993; Goshorn et al., 1993; Han and Kim, 2016).. 억제하는 항체들이 개발되었으며(Dalum et al., 1999) 그 중 휴. 본 연구에서는 TNF-α 항원과 휴미라 항체의 결합 메커니즘. 미라가 매출 세계 1위를 차지하는 블록버스터급 치료용 항체. 과 휴미라의 VH와 VL이 유래된 germline gene family를 결정. 이다(Bong and Lee, 2020). 휴미라는 2018년 10월 유럽 특허. 하기 위해 바이오인포메틱스 연구를 수행하였으며 분자생물. 가 만료되면서 ‘바이오시밀러(biosimilar)’라는 새로운 용어. 학적 연구로, 휴미라 ScFv (Humira)와 leucine zipper (ScZip) 또. 가 등장하였는데 바이오시밀러는 이미 허가된 오리지널 치료. 는 isoleucine zipper (ILZ)가 융합된 휴미라 ScFv (HumiraScZip. 용 항체와 치료 효과가 거의 유사한 복제 항체이다. 이와 같이. 또는 HumiraILZ)들이 대장균에서 수용성 단백질로 발현되는. 치료 효과가 검증되고 특허 기간이 만료되어 국내외적으로 많. 지를 분석하였으며 특히 HumiraScZip의 dimerization과 Hu-. 은 바이오 회사들이 바이오시밀러 휴미라 생산에 주력하고 있. miraILZ의 trimerization 여부를 확인하였다. 최종적으로 발현. 다. 그러나 항체 치료제는 자연 항체이므로 여러 가지 동물 세. 된 세 종류의 휴미라 ScFvs들의 항원 결합력을 측정하여 각. 포를 숙주 세포로 사용하여 생산해야 하므로 항체 생산량에. ScFv들의 항원 결합력을 Kd값으로 수치화하여 di/trimerization. 있어 제한적이고 저가로 대량 생산하기에는 설비 및 비용적인. 에의 휴미라 ScFv의 항원결합력 증가효과를 결정하였다.. 면에서 제약을 받는다(Verma et al., 1998). 또한 자연항체는 Fc를 포함하므로 면역 세포, 상피 세포 및 섬유아세포 등 다양 한 세포들의 표면에 존재하는 Fc 수용체와 결합하여 항체의 항원특이적 결합과 관계없는 비특이적 결합을 발생시킬 수 있 으므로(Kuonen et al., 2010; Andersen et al., 2016) Fc가 제거. 재료 및 방법 TNF-α 항원 및 휴미라 항체 발현벡터 구축. 된 항체 단편들이 제조되었다. 즉 항체 유전자 클로닝 기술을. 종양괴사인자-알파(TNF-α) 항원은 합성된 유전자를 주형. 활용하여 항원 결합 부위인 VH와 VL만을 갖는 Fv 항체단편. 으로 하여 TNF-α 특이적 프라이머(Supplementary data Table. 또는 Fv보다 더 안정된 구조를 갖기 위해 Fv 항체단편을. S1A)를 이용하여 중합효소연쇄반응(polymerase chain reaction:. 15~18개 아미노산으로 구성된 펩타이드 링커(linker)로 연결. PCR)을 수행하여 유전자를 증폭하였고 유전자증폭을 위한 중. 한 single-chain variable domain fragment (ScFv) 항체 단편이. 합효소로는 GoTaq 효소(Promega)를 이용하였으며 SfiI-NotI. 주로 개발되었다(Bird et al., 1988; Huston et al., 1988; Skerra. 제한효소(New England Biolabs)를 이용하여 pET-22b 발현벡. 미생물학회지 제56권 제3호.
(3) Development of di/trimeric Humira ScFv ∙. 터(Invitrogen)로 클로닝 하였다(Min et al., 2001; Park et al., 2005). 한편 휴미라 항체유전자는 IMGT database와 Protein data bank (PDB)를 검색하여 얻어진 휴미라의 VH와 VL 유전 자 배열 정보를 기초로 휴미라 ScFv 유전자를 합성하였으며 (Bionix) 합성된 휴미라 ScFv유전자를 주형으로 하여 각각 특 이적인 프라이머(Supplementary data Table S1B and C)를 이 용하여 Humira, HumiraScZip, 및 HumiraILZ 유전자들을 클 로닝 하였으며 특히 HumiraScZip과 HumiraILZ의 경우 leucine zipper (ScZip) 및 isoleucine zipper (ILZ) 유전자를 포함하는 pUC119를 자체 제작하여 발현 벡터로 사용하였다. 즉, ScZip 또는 ILZ 유전자의 upstream에 휴미라 ScFv 항체 유전자를 SfiI과 NotI의 제한효소를 이용하여 클로닝 하면 자동적으로 ScZip과 ILZ가 융합된 ScFv 항체발현벡터가 구축되도록 제 작하였다.. 휴미라 VH 및 VL 분석. 195. 항원과 융합항체의 발현 및 정제 TNF-α 항원 및 휴미라 ScFv 항체 유전자를 포함한 발현 벡 터를 대장균 BL21star (DE3) (Invitrogen)에 형질전환 하였 다. 형질 전환된 BL21star (DE3)를 사용하여 이전 논문에서 보고된 방법으로 발현을 시도하였다(Park et al., 2011; Kim et al., 2015, 2018; Han and Kim, 2016). 즉 ampicillin (SigmaAldrich)이 함유된 2XTY 액체배지를 사용하여 25°C에서 5.5 h 동안 진탕 배양하여 TNF-α 항원 및 휴미라 ScFv 항체 단백 질 발현을 시도하였으며, 발현이 끝난 세포들을 삼투 현상 (osmosis)을 유도하는 완충 용액을 처리하고 초음파 파쇄 (sonication)를 시도하여 세포를 완전히 용해(lysis) 시켰다. 용 해된 세포를 원심 분리(10,000 rpm)하여 수거된 상등액을 Ni+NTA-아가로스 칼럼(Incospharm)에 결합시키고 250 mM의 imidazole (Daejung)을 처리하여 발현된 단백질들을 정제하 였다. 특히 TNF-α 항원 단백질은 소수성(hydrophobicity)이 높아 초음파 파쇄 후 8 M urea (Samjun)를 추가로 처리하여 수. 휴미라의 VH 및 VL 유전자가 유래되어진 germline origin. 용화(solubilization) 시킨 후 정제하였다. 정제된 단백질들은. 을 결정하기 위해 IMGT database (http//www.imgt.org)를 이. SDS-PAGE에 전기 영동하고 Coomassie 용액으로 염색하여. 용하여 휴미라의 germline 유전자를 구성하는 V, D, J segment. 발현 유무를 확인하였다. 또한 Bradford protein assay (Thermo. origin을 결정하였으며 항원과 직접적으로 결합하는 초 가변. Fisher Scientific)를 이용해 정제된 단백질들의 농도를 결정하. 부위(complementarity determining region: CDR)의 loop class. 였다.. 결정은 Chothia에 의해 제안되어진 방법(Chothia and Lesk, 1987)으로 결정해주는 web site (http//www.bioinf.org.uk)를 이용하여 결정하였다.. 항원-항체 complex의 결합분석. Size-exclusion chromatography 정제된 Humira, HumiraScZip 및 HumiraILZ들은 phosphatebuffered saline (PBS)로 균일화(equilibration)된 Superdex 200 increase10/300 GL column (GE Healthcare)을 이용하여 0.75. Protein Data Bank (http://www.rcsb.org/pdb/)를 검색하여. ml/min의 flow rate로 각각의 단백질들을 용출(elution)하여. crystallography에 의해 결정된 TNF-α 항원과 휴미라 항체. chromatogram을 얻었다. 각 단백질의 분자량 결정을 위해 단. complex의 3차 구조(PDB ID: 3WD5)를 찾아내고 이 구조를. 백질마커(Bio-Rad)를 같은 조건으로 용출하여 얻어진 chro-. Discovery Studio 4.0 visualizer (Accelrys) 프로그램을 이용. matogram과 비교하여 정제된 각 단백질의 분자량을 결정하. 하여 형상화하여(visualize) 휴미라 항체의 항원결합부위인. 였다.. VH와 VL의 3차 구조를 분석하였다. 즉 TNF-α 항원과 직접적 으로 결합하는 휴미라 항체의 3개의 중사슬 초 가변 부위 (heavy chain complementarity determining region: HCDR1, 2, 3)와 3개의 경사슬 초 가변부위(light chain complementarity determining region: LCDR 1, 2, 3)에 존재하는 아미노산 잔기 들을 결정하고 그 아미노산 잔기들이 관여하는 결합 종류 및 개수 등에 관해서도 분석하여 항원-항체 결합에 가장 중요한 CDR을 결정하였고 이 data를 종합하여 VH와 VL중 어느 도메 인이 항원-항체 결합에 더 관여하는지를 분석하였다.. Enzyme-linked immunosorbent assay (ELISA) 냉장온도(4°C)에서 TNF-α 항원을 PBS에 녹여 96 well plate (Nunc)에 밤새 고정(immobilized)시키고 고정된 plate는 PBST (PBS + 0.1% tween)로 세척하고, PBS-TS (PBS + 0.1% tween + 2% skim milk)로 blocking하였다. Blocking 완충용액에 1차 항체로 각 ScFv 항체(Humira, HumiraScFv 및 HumiraILZ)를 처리하여 항원-항체반응을 시도하였고, 다시 PBS-TS로 세척 한 후 horseradish peroxidase (HRPO)가 연결된 anti-FLAG 항 체(Sigma-Aldrich)를 2차 항체로 처리하였으며 최종적으로 Korean Journal of Microbiology, Vol. 56, No. 3.
(4) 196. ∙ Kim et al.. PBS-TS로 세척을 한 다음 ABTS [2,2'-Azino-bis (3-ethylben-. 수(koff)를 측정하고 최종적으로 항원-항체 반응의 결합력을 나. zothiazoline-6-sulfonic acid) diammonium salt] (Sigma-Aldrich). 타내는 해리 평형 상수(Kd)를 측정하였다.. 발색기질을 첨가하여 ELISA plate reader (Biochrome)로 파장. HumiraScZip과 HumiraILZ의 3차 구조 모델링. 450 nm에서 흡광도를 측정하여 항원-항체 반응을 측정하였다.. HumiraScZip 및 HumiraScZip 3차 구조 모델링을 위해서. Western blot. Protein Data Bank (PDB)에서 leucine zipper (ScZip, PDB ID:. 또 다른 항원-항체반응 측정 방법으로 TNF-α 항원을 전기 영. 3BAS)와 isoleucine zipper (ILZ, PDB ID: 1GCM)의 3차 구조. 동 방법으로 SDS-PAGE하고 nitrocellulose membrane (Milli-. 를 검색하였다(Harbury et al., 1994). Discovery Studio 2.5. pore)으로 transfer하였다. Transfer된 membrane은 PBS-TS로. (Accelrys)라는 단백질 3차 구조 모델링 프로그램을 이용하여. blocking한 다음 PBS-T로 세척하였다. 세척된 membrane은. 검색된 ScZip과 ILZ의 3차원적 X, Y, Z 좌표와 이미 결정된. Humira, HumiraScZip, 및 HumiraILZ가 함유된 PBS-TS를 1. Humira의 3차 구조(PDB ID: 3WD5) 좌표 사이에 mIgG3. 차 항체로 사용하여 membrane에 흡착된 TNF-α와 항원-항체. hinge에서 유래된 linker의 3차 구조 좌표를 삽입하여 두 단백. 반응을 시키고 PBS-T로 세척한 다음 2차 항체로 horseradish. 질을 융합시킨 다음 dimeric HumiraScZip와 trimeric Humira. peroxidase (HRPO)가 연결된 anti-FLAG 항체(Sigma-Aldrich). ILZ 융합항체의 3차 구조를 모델링하였다. 모델링 된 3차 구. 을 첨가하여 반응시킨 후 PBS-T로 세척하였다. 기질로 3,3'-. 조는 Ramachandran plot, energy minimization 등의 검증. Diaminobenzidine (DAB) (Sigma-Aldrich)을 처리하여 발색. (verification)단계를 수행하여 모델링 된 3차 구조의 정확도를. 을 시켜 Humira, HumiraScZip, 및 HumiraILZ 항체의 TNF-α. 검증하고 rotamer library를 이용한 3차 구조를 구성하는 아미. 항원에 대한 결합력을 확인하였다.. 노산 side chain을 교정하는 refinement 단계를 수행하여 3차 구조 모델링을 최종결정하고 Discovery Studio 4.0 visualizer. BLI를 이용한 Humira, HumiraScZip, 및 HumiraILZ의 binding kinetics 측정. (Accelrys) 프로그램을 이용하여 형상화하였다.. 세 종류 휴미라 ScFv (Humira, HumiraScZip, 및 HumiraILZ). 결 과. 의 TNF-α에 대한 binding kinetic curve를 결정하기 위해 BLI system (ForteBio) software인 BLItz Pro 1.2을 사용하였다. 즉. 단백질 발현 벡터 구축. 정제된 TNF-α를 10 μg/ml 농도로 PBS에 녹여 상온에서 Anti-. 항원으로 사용될 TNF-α, Humira, HumiraScZip, 및 Humira. Penta-HIS (HIS1K) biosensor (ForteBio)에 결합시키고 PBS. ILZ ScFv 유전자는 각각 특이적인 프라이머(Supplementary. 로 세척하여 biosensor 표면을 saturation시켜 비특이적 결합. data Table S1)를 이용하여 PCR을 수행한 결과 각각 471, 792,. 을 최소화하여 baseline을 안정화시켰다. PBS로 투석되고 0.1. 930, 930 bp 크기로 증폭되었으며(Table 1 and Fig. 1) 증폭된. mg/ml의 농도로 맞춘 Humira, HumiraScZip 및 HumiraILZ를. PCR산물을 SfiI-NotI 제한효소로 절단하여 TNF-α는 pET-22b. 각각 처리하여 HIS1K biosensor에 결합된 TNF-α에 결합을 시. 발현 벡터로 클로닝 하였고(Table 1 and Fig. 2) 세 종류의 휴미. 도하였다. 이때 항원-항체결합을 촉진하기 위해 1,000 rpm으. 라 ScFv는 pUC119 발현 벡터로 클로닝 하였다(Table 1 and. 로 진탕 반응을 시도하였다. 발생된 항원-항체결합은 분광학. Fig. 4). 특히 HumiraScZip pUC119 발현벡터는 ScZip을 포함. 적으로 실시간 탐지되어 일정한 형태의 kinetic curve를 나타. 하도록 하여 휴미라 ScFv의 dimerization을 유도하도록 하였. 내게 되고 이를 이용하여 결합 속도 상수(kon)와 해리 속도 상. Table 1. Biochemical properties of TNF-α, Humira, HumiraScZip, and HumiraILZ Gene size (bp). Amino acid number. Molecular weight (kDa). Isoelectric point (pI). Charge at pH 7.00. Multimer. TNF-α. 471. 157. 17.35. 7.37. 0.6. Monomer. Humira. 792. 264. 28.59. 7.83. 1.43. Monomer. HumiraScZip. 930. 310. 33.70. 8.53. 3.53. Dimer. HumiraILZ. 930. 310. 33.74. 8.53. 3.53. Trimer. 미생물학회지 제56권 제3호.
(5) Development of di/trimeric Humira ScFv ∙. 197. pET-22b 발현 벡터와 pUC119 발현 벡터는 공통적으로 IPTG에 의한 단백질 발현을 촉진하였고 아미노 말단에 pelB leader (Lei et al., 1987)를 삽입하여 TNF-α 및 세 종류의 휴미 라 ScFv (Humira, HumiraScZip, HumiraILZ)들을 대장균의 막 주변 공간(periplasmic space)으로 이동 시켜 수용성 단백질 로 분비를 촉진하였다(Figs. 2 and 4). 단백질 발현에 사용된 BL21sta (DE3)는 lacZ promoter 외에 bacteriophage T7 polymerase 유전자가 대장균의 염색체에 삽입되어 IPTG에 의해 T7 promoter를 활성화시켜 전사(transcription)를 촉진할 수 있 으며 lon 및 외막(OmpT) proteases 등의 단백질분해효소 유전 자가 결핍되어 외부 단백질의 발현을 증강시킬 수 있으며 또 한 RNA 분해효소 즉 RNaseE 유전자 돌연변이로 외부단백질 발현 시 mRNA 변성을 줄이고 mRNA transcripts 안정성을 높 여 외부 단백질의 발현을 극대화시킬 수 있는 이점을 가지고 있으므로 본 연구에서 발현용 대장균으로 사용되었다. 세 종 Fig. 1. DNA gel analysis of TNF-α, Humira, HumiraScZip, and Humira ILZ. TNF-α genes amplified by PCR was cloned into pET-22b expression vector by SfiI-NotI digestion. Three different ScFv genes amplified by PCR were cloned into pUC 119 expression vector by SfiI-NotI digestion.. 류 휴미라의 카르복시말단에는 FLAG 태그(DYKDDDDKG) 를 삽입하여 ELISA 및 western blot에서 항원 결합력을 탐지 하는데 이용되었고 발현되는 모든 단백질의 카르복시말단에 는 공통적으로 헥사히스티딘(HHHHHH) 태그를 삽입하여. 으며 HumiraILZ pUC119 발현 벡터는 ILZ를 포함하도록 하. Ni+-NTA-agarose affinity chromatography를 이용하여 발현. 여 휴미라 ScFv의 trimerization을 유도하도록 하였다(Table 1. 된 단백질을 정제할 수 있도록 제작하였다(Figs. 2 and 4). 정제. and Fig. 4). 휴미라 ScFv를 구성하는 VH와 VL 사이에 linker. 단계에서 소수성 단백질인 TNF-α는 초음파 파쇄 후 다시 Urea. 1 (GSTSGSGKPGSGEGSTKG)을 사용하였고 ScFv와 ScZip/. 처리를 하여야만 정제 가능하였으나 세 종류의 Humira ScFv. ILZ 사이에 linker 2 (PKPSTPPGSS)를 삽입하여 각 도메인들. 는 pH 7.0에서 모두 친수성을 나타내어 초음파 파쇄만으로도. 의 독립적인 3차 구조형성(folding)을 용이하게 하였다(Fig.. 정제가 가능하였다(Figs. 2 and 5).. 4). 특히 TNF-α는 등전점(pI)이 7.37으로 pH 7.0에서 소수성. 한편 발현되는 단백질들의 생화학적 특성을 알아보기 위해. 을 나타내므로 대장균에서 발현 시 비수용성(insolubility)을. 클로닝된 TNF-α 및 휴미라 ScFv의 유전자배열을 vectorNTI. 나타내어 정제과정에서 손실이 발생할 수 있으므로 pUC119. 프로그램(Invitrogen)에 입력시켜 아미노산 배열을 결정하였. 발현벡터의 lacZ promoter보다 활성이 더 큰 T7 promoter를. 으며 이 data를 이용하여 각 단백질들의 분자량, 등전점(pI) 및. 사용하는 pET-22b 발현벡터로 클로닝 하였다(Fig. 2).. 전하(charge) 등을 결정하였다(Table 1).. (A). (B). Fig. 2. Schematic outline for the construction of TNF-α in pET-22b expression vector panel (A) and SDS-PAGE analysis of purified TNF-α panel (B). TNF-α gene was assembled by gene synthesis and digested with SfiI and NotI and cloned into pET-22b expression vector. T7 promoter is activated by IPTG for overexpression. PelB leader transports expressed proteins into periplasmic space for secretion. His6 is hexahistidine tag for detection and purification of expressed proteins. pET-22b expression vector contains both ampicillin-resistance gene as a selection marker and ColE1 as a replication origin.. Korean Journal of Microbiology, Vol. 56, No. 3.
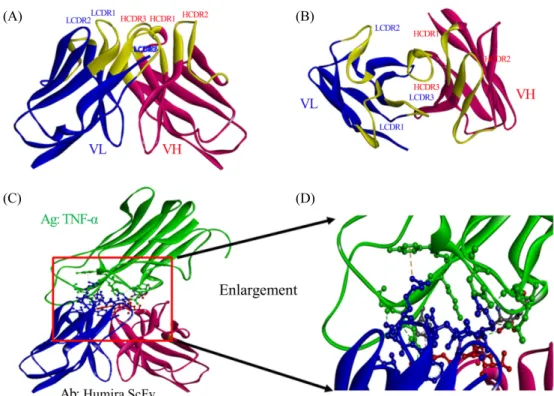
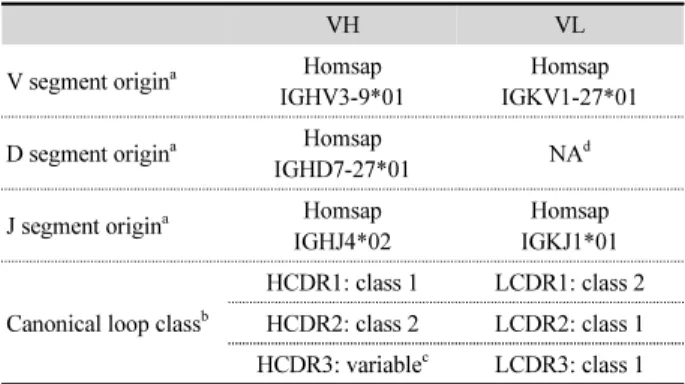
(6) 198. ∙ Kim et al.. (A). (B). (C). (D). Fig. 3. Three dimensional structure of Humira Fv bound to TNF-α. The crystal structure of Humira Fv was extracted from Protein Data Bank (PDB). Side view and top view are shown in panel (A) and (B) individually. The crystal structure of Humira Fv–TNF-α complex is shown in panel (C). Enlargement of panel (C) is shown in panel (D) to analyze binding properties of complex. Each VH and VL is colored in red and blue. CDR loop regions in VH and VL are colored in yellow.. Fig. 4. Construction of Humira, HumiraScZip, and HumiraILZ. The dimeric HumiraScZip and trimeric HumiraILZ ScFvs were constructed from monomeric Humira ScFv expression vector derived from pUC119 containing a 5'-pelB leader sequence, leucine zipper, or isoleucine zipper motif and 3'-hexahistidine tag at C-terminus of ScFv. The murine IgG3 hinge domain was inserted for the connection of ScFv to each zipper domain. In addition, the FLAG tag was added to determine antigen binding activity of each ScFv in ELISA and western blot.. 휴미라 항체 VH 및 VL 유전자 분석 IMGT database (http://www.imgt.org)를 이용하여 휴미라 VH와 VL의 origin germline gene segment를 추정한 결과 VH 유전자는 인간 germline VH 유전자 중에서 V segment는 IGHV39*01에서,D segment는 IGHD7-27*01에서, J segment는 미생물학회지 제56권 제3호. IGHJ4*02에서 유래되었음을 확인하였다. 한편 VL 유전자는 인간 germline VL 유전자 중에서 V segment는 IGKV1-27*01 에서, J segment는 IGKJ1*01에서 유래되었음을 확인할 수 있었다(Table 3). CDR loop 3차 구조는 일정한 loop 구조를 갖 는 group으로 분류 가능한데 이를 canonical loop class라 한다.
(7) Development of di/trimeric Humira ScFv ∙. 199. Table 2. Analysis of VH and VL sequences of Humira ScFv VH. VL. V segment origina. Homsap IGHV3-9*01. Homsap IGKV1-27*01. D segment origina. Homsap IGHD7-27*01. NAd. J segment origina. Homsap IGHJ4*02. Homsap IGKJ1*01. HCDR1: class 1. LCDR1: class 2. Canonical loop classb. HCDR2: class 2. LCDR2: class 1 c. HCDR3: variable. Fig. 5. SDS-PAGE analysis of Humira, HumiraScZip, and HumiraILZ expressed in Escherichia coli. The Humira (lanes 1~5), HumiraScZip (lanes 6~10), and HumiraILZ (lanes 11~15) expressed in E. coli were purified in Ni+-NTA-agarose column and analyzed in 15% polyacrylamide gel under heated reducing condition.. LCDR3: class 1. a. V segment origin, D segment origin and J segment origin were determined based on IMGT database (http://www.imgt.org). b Canonical loop class was determined by Andrew C.R. Martin’s Bioinformatics Group (http://www.bioinf.org.uk). c HCDR3 loop cannot be classified due to its high diversity. d Not available. 가 결합에 주로 관여함을 알 수 있었다. VL의 항원 결합에 관 여한 결합은 수소 결합과 정전기적 결합이었으며 LCDR1이. (Chothia and Lesk, 1987). 따라서 canonical loop class를 결정. LCDR3보다 항원과 더 많은 수소결합을 형성하는 특징을 나. 해 주는 website (http://www.bioinf.org.uk)를 활용하여 휴미. 타내었다(Table 3 and Fig. 3).. 라 VH와 VL CDR의 canonical loop class를 결정하였다(Table 2 and Supplementary data Fig. S2). 그러나 VH의 HCDR3는 V,. 항원과 융합항체의 발현 및 정제. D, J segment들의 recombination에 의해 VH를 구성하는 부위 이므로 유전자들의 다양한 변화를 예상할 수 있으며 또한 결. SDS-PAGE에서 각 단백질들의 발현유무를 확인하고 분자. 합력 향상(affinity maturation)을 위해 mutation도 활발하게 일. 량을 분석해 본 결과 TNF-α는 17.35 kDa, Humira는 28.59. 어나는 지역이므로 CDR 중에서도 가장 가변성이 높아(high. kDa, HumiraScZip와 HumiraILZ는 ScZip과 ILZ가 융합되어. variability) canonical loop class를 특정할 수 없었다(Table 2).. 있으므로 각각 33.70 kDa, 33.74 kDa으로 분석되어 대장균에 서 모두 수용성 단백질로 성공적으로 발현 및 정제됨을 확인 하였다(Fig. 5).. 항원-항체 complex의 결합분석 TNF-α 항원과 휴미라 항체 complex의 3차 구조(PDB ID:. Size-exclusion chromatography (SEC). 3WD5)를 형상화하여 휴미라의 TNF-α 항원에 직접적으로 결 합하는 아미노산 잔기들을 알아내고 결합메커니즘을 분석하. 비공유결합성 di/trimer 형성을 확인하기 위해 SEC을 수행. 였다(Table 3 and Fig. 3). 휴미라 VH를 구성하는 HCDR 중에. 하여, Humira, HumiraScZip, 및 HumiraILZ의 분자량을 결정. 서 HCDR1, 2는 항원 결합에 관여하지 않았으며 HCDR3만 결. 하였다. 즉 크로마토그램의 각 단백질 피크(peak)를 동종 조건. 합에 관여하였으며 관여하는 결합의 종류로 수소 결합과 소수. 에서 첨가하여 얻어진 단백질 분자량 마커의 peak와 비교하여. 성 결합이 관측되었다. 반면에 휴미라 VL은 LCDR1, LCDR2,. Humira, HumiraScZip, 및 HumiraILZ의 분자량을 결정한 결. LCDR3 모두 항원 결합에 관여하였고 특히 LCDR1 및 LCDR3. 과 각각 28.59 kDa, 67.4 kDa, 101.2 kDa으로 결정되었다(Fig.. Table 3. TNF-α binding site analysis of VH and VL sequences of Humira Fv VH HCDR1. HCDR2. VL HCDR3. LCDR1. LCDR2. LCDR3. Total. Hydrogen bond. 0. 0. 3. 7. 0. 4. Salt bridge. 0. 0. 0. 0. 0. 0. 14 0. Electrostatic interaction. 0. 0. 0. 1. 1. 1. 3. Hydrophobic interaction. 0. 0. 2. 0. 0. 0. 2. Total. 0. 0. 5. 8. 1. 5. 19. Korean Journal of Microbiology, Vol. 56, No. 3.
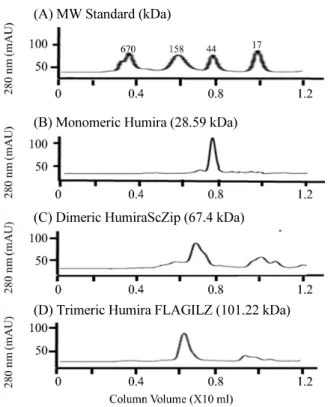
(8) 200. ∙ Kim et al.. (A) MW Standard (kDa). (A). (B). (B) Monomeric Humira (28.59 kDa). (C) Dimeric HumiraScZip (67.4 kDa). (D) Trimeric Humira FLAGILZ (101.22 kDa). Fig. 6. Size exclusion chromatographic profiles of Humira, HumiraScZip, and HumiraILZ on a Superdex 200 increase 10/300 GL column. Each chromatographic profiles were run in PBS at 0.75 ml/min as a flow rate.. Fig. 7. ELISA analysis and western blot to determine TNF-α antigen binding activity of Humira, HumiraScZip, and HumiraILZ. The TNF-α antigen expressed in E. coli was coated onto either 96-well plate for ELISA or transferred onto nitrocellulose membrane for western blot. In ELISA panel (A), each ScFvs and PBS were added as a primary antibody. Color development with ABTS substrate was carried out using anti-FLAG antibody conjugated with horseradish peroxidase (HRPO) as a secondary antibody. PBS was used as a negative control. In westerm blot panel (B), Humira (lane 1), HumiraScZip (lane 2), and HumiraILZ (lane 3) were added as a primary antibody into nitrocellulose membrane blotted with TNF-α antigen. Color development with DAB substrate was carried out using anti-FLAG antibody conjugated with horseradish peroxidase (HRPO) as a secondary antibody.. (A). 6). 이 결과와 Fig. 5를 비교하여 HumiraScZip과 HumiraILZ에 서 ScZip과 ILZ 도메인이 비공유결합성 di/trimer를 형성한다 는 결론을 내릴 수 있었다(Fig. 6).. Humira, HumiraScZip 및 HumiraILZ의 항원 결합력 측정 발현된 Humira, HumiraScZip, 및 HumiraILZ의 항원 결합 력을 분석하기 위해 ELISA를 수행한 결과 세 종류 휴미라 ScFv 모두 negative control과 비교해볼 때 TNF-α 항원에 결합 력을 나타내었다(Fig. 7A). 또한 western blot을 이용한 항원 결 합력 분석에서도 HumiraScZip과 HumiraILZ은 TNF-α 항원 에 대한 항원 결합력을 나타내었으나 Humira의 경우에는 항 원 결합력을 관찰할 수 없었다(Fig. 7B).. BLI를 이용한 Humira, HumiraScZip 및 HumiraILZ의 binding kinetics 측정 Humira, HumiraScZip, 및 HumiraILZ의 TNF-α 항원에 대한 binding kinetics 분석을 위해 bio-layer interferometry (BLI)를. (B) koff (S-1). Kd (M). Humira. 3. 2.24 × 10. 4.73 × 10-2. 2.11 × 10-5. HumiraScZip. 1.51 × 103. 2.12 × 10-4. 1.40 × 10-7. 3. -5. 1.53 × 10-8. ScFc variants. HumiraILZ. kon (M-1S-1). 3.80 × 10. 5.80 × 10. Fig. 8. TNF-α antigen binding constant (Kd) determination of Humira, HumiraScZip, and HumiraILZ by bio-layer interferometry (BLI). In panel (A) TNF-α antigen (10 μg/ml) was captured by His1K biosensor until saturation and followed by association of monomeric Humira, dimeric HumiraScZip and trimeric HumiraILZ ScFvs and dissociation. After collection of interaction curve, The kon and koff values were obtained by fitting to achieve the fitting parameter R2 > 0.99 and The final Kd value was determined. Panel (B) showed kinetic binding parameters for the interactions of Humira, HumiraScZip, and HumiraILZ with TNF-α determined by panel (A)’s kinetic curve.. 수행하여 관찰된 kinetic curves를 분석한 결과, 세 종류 휴미라 ScFv 항체 모두 HIS1K biosensor에 고정된 TNF-α 항원에 결. 합을 나타내지 않았다(Fig. 8A). 관찰된 kinetic curves로부터. 합력을 나타내었으나 대조군인 PBS는 TNF-α biosensor에 결. 휴미라 ScFv들의 결합속도상수(kon)를 결정한 결과 Humira. 미생물학회지 제56권 제3호.
(9) Development of di/trimeric Humira ScFv ∙. (A). (B). (C). (D). (E). (F). 201. Fig. 9. Homology modeling of Humira, HumiraScZip, and HumiraILZ. Three dimensional structures of Humira are shown in both panel (A and D) as a monomer. As shown in both panel (B and E), HumiraScZip 3D structures are visualized as a dimer due to non-covalent association induced by leucine zipper. In panel (C and F), HumiraILZ are shown as a trimer due to non-covalent association induced by leucine zipper. Front and top view are shown in panel (A, B, C and D, E, F) individually. Both HCDR and LCDR are colored as blue.. ScZip 및 HumiraILZ가 Humira보다 각각 0.7배, 1.7배 높은 값. 들의 등전점(pI)에 의해 결정되는 pH 7.0 에서 전하(charge)가. 을 나타내었고 해리 속도 상수(koff)값에서는 각각 223배, 815. 발현되는 단백질들의 수용성(solubility)에 미치는 영향을 분. 배 낮은 값을 나타내었으며 실질적인 결합력을 의미하는 해리. 석하였다. 즉 TNF-α는 pH 7.0에서 거의 전하를 띄지 않으므. 평형 상수(Kd)값은 HumiraScZip 및 HumiraILZ가 Humira보. 로 가장 높은 소수성을 나타내어 정제 시 소수성 단백질들을. 다 150배, 1,380배 낮게 측정되었다(Fig. 8B).. 수용화시키는 Urea처리가 필요한 것으로 추정된다. 또한 HumiraScZip과 HumiraILZ는 Humira에 비해 ScZip와 ILZ가. HumiraScZip과 HumiraILZ의 3차 구조 모델링 컴퓨터를 이용한 homology modeling에 의해 형상화된 HumiraScZip과 HumiraILZ의 3차 구조를 보면 2개의 ScZip 이 비공유결합을 이루어 dimer를 형성하거나 3개의 ILZ가 비 공유결합되어 trimer를 형성함을 추정할 수 있었다. 따라서 monomeric Humira에 ScZip과 ILZ가 융합되어 비공유결합성 di/trimer를 형성하여 그 결과 2개 또는 3개의 항원 결합 부위를 갖게 되어 항원 결합력을 증가시키는 효과를 추정할 수 있었 다(Fig. 9).. 융합되어 수용성이 향상된 효과가 관찰되었는데 이는 ScZip 및 ILZ 도메인이 염기성을 나타내어 융합된 휴미라 ScFv들의 양전하를 증가시켜 HumiraScZip과 HumiraILZ가 Humira에 비해 수용성이 증가되어 발현율이 더 향상된 것으로 추정 가 능하다. 다기능성 융합단백질(bifunctional fusion protein)을 발현 하는 경우 융합된 단백질의 접힘(folding)이 독립적으로 일 어나도록 유연성(flexibility)을 제공하는 linker가 필수적이 다. 본 연구에서는 휴미라 ScFv 내의 VH와 VL 도메인을 연결 하는 linker 1 (GSTSGSGKPGSGEGSTKG)과 휴미라 ScFv와 ScZip 또는 ILZ 도메인을 연결하는 linker 2 (PKPSTPPGSS). 고 찰. 를 사용하였는데 Linker 1과 linker 2를 구성하는 아미노산 잔 기들 대부분은 glycine (G)과 serine (S)인데 glycine은 R-group. TNF-α 항원과 세 종류의 휴미라 ScFv 항체 단백질들은 대. 이 hydrogen으로 구성되어 단백질의 전하에 영향을 미치지 않. 장균에서 성공적으로 발현 및 정제 되었으며 특히 각 단백질. 고 가장 작은 분자량을 나타내어 3차 구조 형성에 중요한 steric. Korean Journal of Microbiology, Vol. 56, No. 3.
(10) 202. ∙ Kim et al.. hindrance를 발생시키지 않아 융합시키는 두 단백질의 2차 구. coiled coil α helix를 형성하도록 하여 주변의 동일한 α helix간. 조 및 3차 구조의 folding에 방해를 주지 않는 이점이 있으며 극. 에 비공유결합인 정전기적 결합 및 소수성 결합에 의한 안정. 성을 띠는 serine (S)은 반복된 glycine (G)의 소수성을 상쇄시. 된 di/trimerization형성을 가능케 한다(Vinson et al., 2006). 그. 켜 발현하고자 하는 단백질의 수용화를 촉진시키므로 linker. 러므로 ScZip 또는 ILZ 도메인을 융합시키면 ScFv 항체의. 를 구성하는 주 아미노산들이다(Whitlow et al., 1993; Solar. di/trimerization이 유도되어 ScFv 항체의 항원 결합 부위 개수. and Gershoni, 1995; Le Gall et al., 2004). 그러나 보고되어진. 를 증가되어 항원 결합력을 증가시키는 방법으로 활용되었다. 일반적인 linker는 glycine과 serine으로만 구성되어있어 낮은. (Landschulz et al., 1988; O'Shea et al., 1989; De Kruif and. 친수성에 의한 발현율을 감소시킬 수 있으므로 본 연구에서. Logtenberg, 1996; Vinson et al., 2006). 특히 이 방법은 원래 낮. 사용된 linker 1에서는 threonine (T), lysine (K), glutamate (E). 은 항원 결합 친화력(low antigen binding affinity)을 갖는 자연. 등의 친수성 아미노산들을 첨가하여 친수성을 증가시켰으며. 항체의 항원 결합력을 유전자 변이(mutation) 없이 쉽게 증가. proline (P)을 비롯한 비교적 분자량이 큰 아미노산들로 linker. 시킬 수 있을 뿐만 아니라 핵산, 다당류(polysaccharide) 같은. 1을 구성하여 휴미라 ScFv들이 안정적으로 발현될 수 있도록. 비교적 분자량이 큰 구조를 갖는 중합체(polymer) 또는 세포. 고안되었다. 생쥐의 IgG3 hinge 펩타이드에서 유래된 linker 2. 표면에 존재하는 항원인 경우 다수의 항원-항체 결합을 필요. 는 비교적 긴 upper hinge로서 자연항체의 Fab와 Fc간에 유연. 로 하므로 아미노산 잔기들을 변화시켜 근본적인 항원 결합. 성(flexibility)을 제공한다(Dangl et al., 1988; Tan et al., 1990).. 친화력(antigen binding affinity)을 증가시키기보다는 항체의. 생쥐의 IgG2a와 IgG2b upper hinge가 더 flexibility를 제공하. 항원 결합 부위 개수를 증가시켜 항원 결합력을 증가시키는. 나 IgG3의 hinge가 더 친수성이므로 융합항체를 대장균의. 방법이 매우 효과적이다 또한 활용되는 ScZip 또는 ILZ 도메. periplasmic space로 이동하여 분비를 촉진시키므로 HumiraScZip. 인은 분자량이 작고 양극성 나선(amphipathic helices)형태이. 과 HumiraILZ의 linker 2로 활용되었다(Pack and Plüeckthun,. 므로 양극성을 갖는 세포막을 잘 통과하여 대장균에서 발현된. 1992).. ScFv가 쉽게 분비될 수 있도록 하여 대장균에서 낮은 발현율. 휴미라 VH와 VL의 3차 구조를 분석해보면 VH와 VL을 구 성하는 3개씩의 HCDR과 LCDR은 각각 3차원적으로 특징적. 을 나타내는 단백질의 발현을 향상시키는 목적으로도 활용되 었다.. 인 canonical loop class (HCDR3 제외)를 갖게 되고 6개의 loop. 발현된 세 종류의 휴미라 ScFv의 TNF-α 항원에 대한 항원. 구조가 모여 항원과 상보적으로 결합할 수 있는 독특한 hydro-. 특이적 결합을 측정하기 위해 ELISA를 수행한 결과 TNF-α 항. phobic pocket을 형성함을 확인할 수 있었고. TNF-α 항원과 휴. 원에 대한 결합을 나타내었으나 western blot에서 monomeric. 미라 항체의 결합을 분석해보면 첫째, 항체의 항원 결합에는. Humira는 항원에 대한 결합을 나타내지 않았는데 주요 원인. 일반적으로 HCDR이 LCDR보다 더 많이 관여하는 데 반해서. 으로는 첫째, ScFv는 정상 부위(constant region: C)가 결핍된. 휴미라의 경우에는 VL을 구성하는 LCDR이 VH의 HCDR보. 구조이므로 정상부위를 갖춘 Y 형태의 자연 항체보다 구조적. 다 TNF-α 항원과 더 많이 결합하는 특징을 나타내었고 둘째,. 안정도가 떨어지므로 VH 및 VL의 loop 3차 구조에 영향을 미쳐. VL에서 LCDR3가 항원 결합에 주로 관여하는 일반 항체와 달. 항원 결합력이 감소될 수 있다. 둘째, 일반적으로 항체 단백질은. 리 LCDR1이 LCDR3보다 TNF-α 항원과 주도적으로 결합하. 진핵세포인 B임파구에서 생산되는 진핵 세포 당 단백질이므로. 는 특징이 관찰되었다.. 원핵 세포인 대장균에서 발현하는 동안 misfolding되어 항원. SDS-PAGE에서 측정된 Humira, HumiraScZip, 및 Humira. 결합력을 상실되거나 감소되어 질 수 있다(Verma et al., 1998).. ILZ의 분자량은 각각 28.59 kDa, 33.70 kDa, 33.74 kDa으로 모. 특히 대장균에서 발현되는 항체들은 비당화(aglycosylation). 두 monomer를 형성하므로 비공유결합성 di/trimerizaion을 결. 로 인한 misfolding의 가능성이 높으나 당화되는 도메인은. 정할 수 없어 size exclusion chromatography를 수행한 결과 세. 주로 항체의 중사슬의 정상부위 특히 CH2 도메인이므로 Fc. 종류의 휴미라 ScFv의 분자량은 각각 28.59 kDa, 67.4 kDa,. 의 반응기(effector) 기능에만 관계하여 VH와 VL로만 구성된. 101.22 kDa으로 결정되었다. 이 두 결과를 근거로 Humira. 세 종류의 비당화 휴미라 ScFv는 정상적인 항원 결합력을 갖. ScZip과 HumiraILZ는 융합된 ScZip과 ILZ의 비공유결합에. 는 것으로 추정된다(Verma et al., 1998). 셋째, monomeric. 의해 di/trimerization됨을 확인하였다. 한편 ScZip 및 ILZ 도메. Humira는 1개의 항원 결합 부위를 가지므로 2개 이상의 항원. 인은 yeast의 transcriptional activator인 GCN4로부터 유래되. 결합 부위를 갖는 HumiraScZip 또는 HumiraILZ보다 낮은 결. 었으며 구조적으로 7개의 펩타이드가 반복적으로 나열된. 합력을 나타내어 western blot에서 충분한 시그널을 나타내지. 미생물학회지 제56권 제3호.
(11) Development of di/trimeric Humira ScFv ∙. 못한 것으로 추정할 수 있다. 이러한 추정을 확인하기 위해 Humira, HumiraScZip, 및 HumiraILZ의 TNF-α 항원에 대한 해리평형상수(Kd)를 결정하였는데 HumiraScZip과 Humira ILZ이 Humira보다 각각 150배, 1,380배 높은 항원 결합력을 나타내었으며 또한 HumiraILZ는 HumiraScZip보다 9배 높은 항원결합력을 나타내었다. 이 결과는 HumiraScZip과 Humira ILZ가 di/trimerization에 의해 2개 또는 3개의 항원 결합 부위 를 형성하여 1개의 항원 결합 부위를 갖는 monomeric Humira 보다 낮은 해리 속도 상수(koff)를 갖게 되어 항원-항체 결합체 를 안정화시키는 효과를 나타내므로 높은 항원 결합력을 갖는 것으로 결정할 수 있었다. 특히 trimeric HumiraILZ는 dimeric HumiraScZip에 비해서도 낮은 해리 속도 상수(koff)값에 의한 더 높은 결합력을 나타내어 ScZip및 ILZ를 융합된 Humira ScZip및 HumiraILZ은 보고되어진 논문들에서와 같이 비공 유결합에 의한 di/trimer를 형성하여 항원 결합력을 증가시키 는 효과를 얻을 수 있었다(Pack and Plüeckthun, 1992; Pack et al., 1993). 결론적으로 본 연구에서는 생물정보학적 연구로 TNF-α와 휴미라 복합체의 3차 구조를 분석하여 결합에 관여하는 아미 노산 잔기 및 결합 종류 등을 분석하였으며 휴미라의 VH와 VL 이 유래된 germline gene family를 결정하였다. 또한 Homology modeling 방법으로 Humira, HumiraScZip, 및 HumiraILZ의 3 차 구조를 추정하였다. 분자생물학적 연구에서는 monomeric 휴미라 ScFv (Humira) 발현 벡터를 구축하고 이후 Humira의 카르복시말단에 leucine zipper (ScZip) 또는 isoleucine zipper (ILZ)를 융합시킨 휴미라 ScFv (HumiraScZip 또는 Humira ILZ)의 발현 벡터를 구축하여 대장균에서 발현 정제하였다. Humira, HumiraScZip, 및 HumiraILZ는 수용성 단백질로 정 제됨을 확인하였고 특히HumiraScZip과 HumiraILZ는 ScZip 과 ILZ의 비공유결합에 의해 각각 dimer, trimer를 형성하였 다. TNF-α 항원에 대한 결합력을 확인한 결과 HumiraILZ, HumiraScZip 및 Humira 순서로 항원 결합력이 높게 나타났으 며, 항원 결합력을 수치화하여 HumiraScZip과 HumiraILZ의 di/trimer 형성은 항원 결합 개수의 증가로 인해 monomeric. 203. 적 요 면역 반응에 필요하며 면역을 조절하는 사이토카인인 종양 괴사인자-알파(TNF-α)는 종양괴사인자-알파 수용체와 상호 작용이 필수적이다. 휴미라와 같은 바이오시밀러 중화항체를 이용하여 종양괴사인자-알파(TNF-α)와 종양괴사인자-알파 수용체(TNF-α receptor) 사이의 상호 작용을 억제하는 방법이 자가 면역 질환 치료를 위하여 개발되었다. 우리는 단백질 데 이터 은행과 homology modeling을 활용하는 생물정보학적 연구를 통해 휴미라 항체의 종양괴사인자-알파와 결합하는 성질을 분석하였다. 또한 우리는 단백질 데이터 은행에서 휴 미라 중사슬 가변 부위(VH)와 경사슬 가변 부위(VL) 배열을 알아내고 18개의 아미노산 링커로 연결시킨 Humira single chain variable domain 항체(ScFv: Humira)를 제조하였다. 1개 의 항원 결합가로 인한 Humira의 낮은 항원 결합가는 항원 결 합가를 높이기 위해 Humira의 multimerization을 필요로 한다. 그러므로 우리는 dimer를 형성시키는 leucine zipper (ScZip) 와 trimer를 형성시키는 isoleucine zipper (ILZ)를 연결시킴으 로써 dimeric 또는 trimeric Humira ScFv를 구축하였다. Mono/ di/trimeric Humira들은 대장균에서 발현시키고 Ni+-NTA-아 가로스 칼럼으로 정제하였다. 세 종류의 정제된 ScFv들은 SDS-PAGE에서 각각 28.59 kDa, 33.70 kDa, 33.74 kDa으로 탐지되었으나 size-exclusion chromatography에서는 leucine zipper는 비공유결합에 의한 dimer (ScZip)형성을, isoleucine zipper는 비공유결합에 의한 trimer (ILZ) 형성을 유도함을 암시하는 28.59 kDa, 67.4 kDa, 101.22 kDa를 나타내었다. Dimeric 휴미라 ScFv (HumiraScZip)와 trimeric 휴미라 ScFv (HumiraILZ)의 종양괴사인자-알파 항원에 대한 결합력은 ELISA 및 western blot에 의해 관찰되었다. Bio-layer interferometry 분석은 di/trimeric 휴미라 ScFv가 monomer보다 150~ 1,380배의 더 높은 결합력을 보여주었고 이는 leucine zipper 와 isoleucine zipper helix의 비공유결합인 소수성 결합에 의 해 di/trimerization되어 직접적으로 항원 결합력 증가를 유도 할 수 있음을 제시해주고 있다.. Humira의 낮은 항원 결합력을 약 150~1,380배 증가시키는 결 과를 얻었다. 결론적으로 본 연구에서 활용한 항원 결합 부위를 증가시 켜 항원 결합력을 증가시키는 항체 공학 기술은 기존의 항체 치료제 또는 개발되고 있는 항체 치료제의 항원 결합력을 향 상시키는데 활용 가능하며 따라서 치료 효과가 향상된 치료용. 감사의 말 이 논문은 2019~2020년도 창원대학교 자율연구과제 연구 비 지원에 의해 수행되었으며 이에 감사드립니다.. 항체 개발이 가능할 것이다.. Korean Journal of Microbiology, Vol. 56, No. 3.
(12) 204. ∙ Kim et al.. References Andersen MN, Al-Karradi SNH, Kragstrup TW, and Hokland M. 2016. Elimination of erroneous results in flow cytometry caused by antibody binding to Fc receptors on human monocytes and macrophages. Cytometry A 89, 1001–1009. Bird RE, Hardman KD, Jacobson JW, Johnson S, Kaufman BM, Lee SM, Lee T, Pope SH, Riordan GS, and Whitlow M. 1988. Single-chain antigen-binding proteins. Science 242, 423–426. Bondeson J and Maini RN. 2001. Tumour necrosis factor as a therapeutic target in rheumatoid arthritis and other chronic inflammatory diseases: The clinical experience with infliximab (REMICADE). Int. J. Clin. Pract. 55, 211–216. Bong KT and Lee H. 2020. Analysis and estimation for market share of biologics based on Google trends big data. J. Soc. Korea Ind. Syst. Eng. 43, 14–24. Brekke O and Løset GÅ. 2003. New technologies in therapeutic antibody development. Curr. Opin. Pharmacol. 3, 544–550. Brinkmann U, Reiter Y, Jung SH, Lee B, and Pastan I. 1993. A recombinant immunotoxin containing a disulfide-stabilized Fv fragment. Proc. Natl. Acad. Sci. USA 90, 7538–7542. Carmona L, Gómez‐Reino JJ, Rodríguez‐Valverde V, Montero D, Pascual‐Gómez E, Mola EM, Carreño L, Figueroa M, and BIOBADASER Group. 2005. Effectiveness of recommendations to prevent reactivation of latent tuberculosis infection in patients treated with tumor necrosis factor antagonists. Arthritis Rheum. 52, 1766–1772. Chothia C and Lesk AM. 1987. Canonical structures for the hypervariable regions of immunoglobulins. J. Mol. Biol. 196, 901– 917. Dalum I, Butler DM, Jensen MR, Hindersson P, Steinaa L, Waterston AM, Grell SN, Feldmann M, Elsner HI, and Mouritsen S. 1999. Therapeutic antibodies elicited by immunization against TNF-α. Nat. Biotechnol. 17, 666–669. Dangl JL, Wensel TG, Morrison SL, Stryer L, Herzenberg LA, and Oi VT. 1988. Segmental flexibility and complement fixation of genetically engineered chimeric human, rabbit and mouse antibodies. EMBO J. 7, 1989–1994. De Kruif J and Logtenberg T. 1996. Leucine zipper dimerized bivalent and bispecific scFv antibodies from a semi-synthetic antibody phage display library. J. Biol. Chem. 271, 7630–7634. Goshorn SC, Svensson HP, Kerr DE, Somerville JE, Senter PD, and Fell HP. 1993. Genetic construction, expression, and characterization of a single chain anti-carcinoma antibody fused to βlactamase. Cancer Res. 53, 2123–2127. Han SH and Kim JK. 2016. The development of anti-DR4 single-chain Fv (scFv) antibody fused to Escherichia coli alkaline phosphatase. Korean J. Microbiol. 52, 10–17. Harbury PB, Kim PS, and Alber T. 1994. Crystal structure of an isoleucine-zipper trimer. Nature 371, 80–83. Huston JS, Levinson D, Mudgett-Hunter M, Tai MS, Novotný J,. 미생물학회지 제56권 제3호. Margolies MN, Ridge RJ, Bruccoleri RE, Haber E, Crea R, et al. 1988. Protein engineering of antibody binding sites: Recovery of specific activity in an anti-digoxin single-chain Fv analogue produced in Escherichia coli. Proc. Natl. Acad. Sci. USA 85, 5879–5883. Kim CH, Han SH, Kim HM, Han JY, Lim MW, and Kim JK. 2015. The development of murine recombinant single-chain variable domain fragment (scFv) specific to acute non-lymphocytic leukemia (ANLL) cell line HL60. Korean J. Microbiol. 51, 115–125. Kim SW, Wu S, and Kim JK. 2018. The development of anti-DR4 single-chain Fv (scFv) antibody fused to Streptavidin. Korean J. Microbiol. 54, 330–342. Kuonen F, Touvrey C, Laurent J, and Ruegg C. 2010. Fc block treatment, dead cells exclusion, and cell aggregates discrimination concur to prevent phenotypical artifacts in the analysis of subpopulations of tumor‐infiltrating CD11b+ myelomonocytic cells. Cytometry A 77, 1082–1090. Landschulz WH, Johnson PF, and McKnight SL. 1988. The leucine zipper: a hypothetical structure common to a new class of DNA binding proteins. Science 240, 1759–1764. Le Gall F, Reusch U, Little M, and Kipriyanov SM. 2004. Effect of linker sequences between the antibody variable domains on the formation, stability and biological activity of a bispecific tandem diabody. Protein Eng. Des. Sel. 17, 357–366. Lei SP, Lin HC, Wang SS, Callaway J, and Wilcox G. 1987. Characterization of the Erwinia carotovora pelB gene and its product pectate lyase. J. Bacteriol. 169, 4379–4383. Li Y, Carpenito C, Wang G, Surguladze D, Forest A, Malabunga M, Murphy M, Zhang Y, Sonyi A, Chin D, et al. 2018. Discovery and preclinical characterization of the antagonist anti-PD-L1 monoclonal antibody LY3300054. J. Immunother. Cancer 6, 31. Li K, Zettlitz KA, Lipianskaya J, Zhou Y, Marks JD, Mallick P, Reiter RE, and Wu AM. 2015. A fully human scFv phage display library for rapid antibody fragment reformatting. Protein Eng. Des. Sel. 28, 307–316. Min W, Kim JK, Lillehoj HS, Sohn EJ, Han JY, Song KD, and Lillehoj EP. 2001. Characterization of recombinant scFv antibody reactive with an apical antigen of Eimeria acervulina. Biotechnol. Lett. 23, 949–955. O'Shea EK, Rutkowski R, and Kim PS. 1989. Evidence that the leucine zipper is a coiled coil. Science 243, 538–542. Pack P, Kujau M, Schroeckh V, Knüpfer U, Wenderoth R, Riesenberg D, and Plückthun A. 1993. Improved bivalent miniantibodies, with identical avidity as whole antibodies, produced by high cell density fermentation of Escherichia coli. Biotechnology 11, 1271–1277. Pack P and Plückthun A. 1992. Miniantibodies: use of amphipathic helixes to produce functional, flexibly linked dimeric Fv fragments with high avidity in Escherichia coli. Biochemistry 31, 1579–1584. Park DW, Kim ED, Kim SH, Han JY, and Kim JK. 2011. The development of dimerized chicken recombinant single-chain Fv.
(13) Development of di/trimeric Humira ScFv ∙. (scFv) antibody using leucine zipper motif. Korean J. Microbiol. 47, 328–334. Park KJ, Park DW, Kim CH, Han BK, Park TS, Han JY, Lillehoj HS, and Kim JK. 2005. Development and characterization of a recombinant chicken single-chain Fv antibody detecting Eimeria acervulina sporozoite antigen. Biotechnol. Lett. 27, 289–295. Pavlou AK and Belsey MJ. 2005. The therapeutic antibodies market to 2008. Eur. J. Pharm. Biopharm. 59, 389–396. Rau R. 2002. Adalimumab (a fully human anti-tumour necrosis factor α monoclonal antibody) in the treatment of active rheumatoid arthritis: the initial results of five trials. Ann. Rheum. Dis. 61, ii70 –ii73. Reiter Y, Brinkmann U, Jung SH, Pastan I, and Lee B. 1995. Disulfide stabilization of antibody Fv: computer predictions and experimental evaluation. Protein Eng. 8, 1323–1331. Shiraishi T, Suzuyama K, Okamoto H, Mineta T, Tabuchi K, Nakayama K, Shimizu Y, Tohma J, Ogihara T, Naba H, et al. 2004. Increased cytotoxicity of soluble Fas ligand by fusing isoleucine zipper motif. Biochem. Biophys. Res. Commun. 322, 197–202. Skerra A and Plückthun A. 1988. Assembly of a functional immunoglobulin Fv fragment in Escherichia coli. Science 240, 1038–. 205. 1041. Solar I and Gershoni JM. 1995. Linker modification introduces useful molecular instability in a single chain antibody. Protein Eng. Des. Sel. 8, 717–723. Tan LK, Shopes RJ, Oi VT, and Morrison SL. 1990. Influence of the hinge region on complement activation, C1q binding, and segmental flexibility in chimeric human immunoglobulins. Proc. Natl. Acad. Sci. USA 87, 162–166. Tracey D, Klareskog L, Sasso EH, Salfeld JG, and Tak PP. 2008. Tumor necrosis factor antagonist mechanisms of action: a comprehensive review. Pharmacol. Ther. 117, 244–279. Verma R, Boleti E, and George A. 1998. Antibody engineering: comparison of bacterial, yeast, insect and mammalian expression systems. J. Immunol. Methods 216, 165–181. Vinson C, Acharya A, and Taparowsky EJ. 2006. Deciphering B-ZIP transcription factor interactions in vitro and in vivo. Biochim. Biophys. Acta 1759, 4–12. Whitlow M, Bell BA, Feng SL, Filpula D, Hardman KD, Hubert SL, Rollence ML, Wood JF, Schott ME, et al. 1993. An improved linker for single-chain Fv with reduced aggregation and enhanced proteolytic stability. Protein Eng. 6, 989–995.. Korean Journal of Microbiology, Vol. 56, No. 3.
(14)
수치


+3

관련 문서
6%) had a fami l y hi story ofgastri ccancer.Themi croscopi cfi ndi ngsshowed 10 poorl y di fferenti ated adenocarci nomas(45. 0%).Di sease-speci fi c 5 yearsurvi valrate
• Only the single statement (simple or compound) is part of the loop.. • The statements runs from while to ; or }
나프탈렌 등 비선형 단위체, trifluoromethyl 그룹 및 에테르 연결기, di-imide 그룹 을 갖는 di-acid와 3,3’-dihydroxybenzidine을 축합중합 하여 poly(hydroxyamide
본 실험에서는 자가 부식 접착제를 적용하기 전에 인산으로 전처리를 시행한 군 에서 인산 전처리를 시행하지 않은 군보다 미세전단 결합강도가 더 높게 나타났으
DNA 프로브법은 기존의 세균배양법, 생화학검사법 및 DNA-DNA hybri di zati on법 등에 비해 재현성,신속성 및 경제적 측면에서 뛰어나다.그러나 중합
비조사 시료에서 확인된 것처럼 방사선 조사 시료에서도 di methyltri sul fi de,( Z )- propenylmethyltri sul fi de와 di methyldi sul fi de가 주요 화합물로
di d often grow notor very sl owl y under BBL anaerobi c gas system.Authorhas therefore devel oped a new di sposabl e anaerobi c gas system bei ng abl e to create the absol
Assumption: an active customer type i only binding incentive compatibility constraint is the one for type j=i -1.. t* is lowest type of
