Anti-Inflammatory Effects of Ethyl Acetate Fraction from Cnidium officinale Makino on LPS-Stimulated RAW 264.7 and THP-1 Cells
9
0
0
전체 글
(2)
(3)
(4)
(5)
(6)
(7)
(8)
(9)
수치

관련 문서
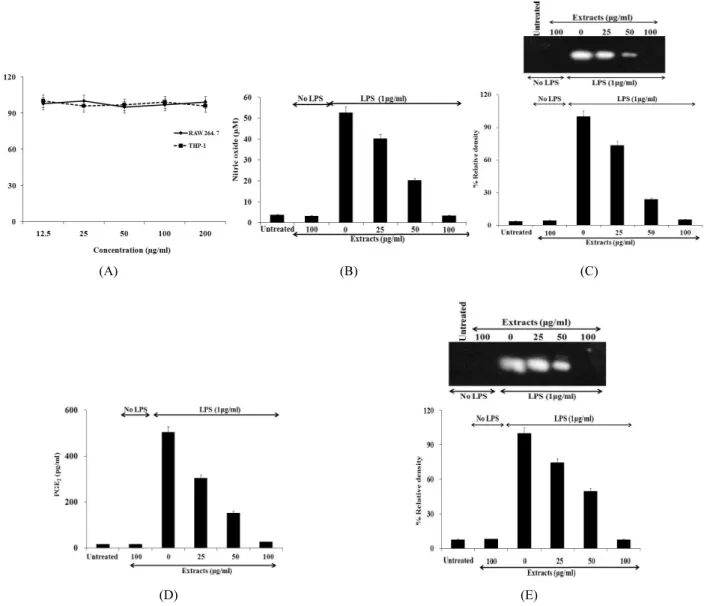
And Western Blotting was used to see the effects of Cnidium officinale MAKINO extracts on inducible Nitric Oxide Synthase (iNOS) and cyclooxygenase-2 (COX-2) expression..
Pro-allergic cytokines were important mediators of allergic inflammation, cell recruitment and allergenic response decided to further investigate the
This study researched the possibility of using natural products for hair growth agents and hair loss prevention materials based on the test for the antioxidant and
TABLE 1.ANTI PROLI FERATI VE EFFECT OF EEAD I N C6CELLS ---7.. Caspase-3/-7 activity
tricuspidata on the production of proinflammatory cytokines in TNFα+IFNγ-stimulated HaCaT cells ...15 Fig.5: The cell viability of sub-fractions from 70% EtOH
HPLC instrument conditions of methanol and ethyl acetate extraction HPLC Prominence LC-20A (Shimadzu, Kyoto, Japan) Detector Diode array detector (DAD, Shimadzu, Kyoto,
포에서 We s t e r nbl ot 을 통해 HJ ME가 i NOS의 단백질 발현을 농도의존적으로 저해 하는 것을 확인하였다( Fi g.6) .그러므로 환삼덩굴은 i NOS의 발현을
It is the purpose of this study to investigate the anti microbacterial Activity and anti inflammatory Effects of water extract Rhododendron brachycarpum
