학 술 논 문
168
유체전단응력에 의하여 3T3-L1 지방세포가 받는 영향
이정근 · 이영훈 · 진희원 · 이서현 · 김지현
연세대학교 보건과학대학 의공학부
Effects of Fluid Shear Stress on 3T3-L1 Preadipocytes
Jeongkun Lee, Yeong Hun Lee, Heewon Jin, Seohyun Lee and Chi Hyun Kim
Department of Biomedical Engineering, Yonsei University, Korea
(Manuscript received 18 June 2018 ; revised 12 August 2018 ; accepted 24 August 2018)
Abstract: Adipocytes affect obesity through the regulation of lipid metabolism. Physical loading is an important reg- ulator of fat tissue. There are ongoing in vitro studies inducing mechanotransduction on 3T3-L1 preadipocytes with mechanical stimulus in order to treat obesity by inhibiting adipogenesis and provoking cell death. In this study, our goal was to suggest a new therapy for obesity by investigating whether fluid shear stress (FSS) changes transcription factors on 3T3-L1 related with adipogenesis and cell death. FSS loading was applied to 3T3-L1 preadipocytes at 1Pa and 1Hz. After loading, bright field images were taken and an immunofluorescence assay was conducted to observe actin stress fiber formation. Western blot analysis was conducted to identify the activation of the ERK pathway as well as the adipogenic factors, which including C/EBPs and PPAR γ. The expression of osteopontin, a protein related to inflammation in adipose tissue, and cell death related factors, Bax, Bcl-2, and Beclin, were also measured. Results showed that FSS stimulated the formation of actin stress fibers in 3T3-L1 and also that the activation of C/EBPs decreased significantly when compared with the control group. PPAR γ activation in the 2 hour FSS group was lower than the 1 hour FSS group, which implied that the results were time dependent. Additionally, there were no dif- ferences in the expression of cell death factors after FSS loading. In summary, similar to other fibroblasts, the for- mation of actin stress fibers induced by mechanotransduction may affect the differentiation of 3T3-L1, leading to inhibition of adipogenesis and inflammation.
Key words: Fluid Shear Stress(FSS), Mechanotransduction, Adipocyte, Adipogenesis, Cell death
I. 서 론
현대에 널리 유행하는 대사 질환 중의 하나인 비만은, 에 너지 대사의 균형에 이상이 생겨 지방조직이 과도하게 증식 하고 인슐린 민감성이 저하되는 등 수 많은 부정적인 증상 을 야기한다[1]. 또한 이러한 증상들은 각종 심혈관계 질환, 당뇨병 등 다른 성인병의 직간접적인 원인이 된다[2]. 비만 의 증상은 지방세포의 수, 크기, 표현형과 밀접한 관련이 있
는데, 예를 들어 지방세포가 분화하여 직경이 커질수록 지 방이 축적되고 세포의 인슐린 민감성이 감소하여 증상이 악 화된다[3].
비만을 직접적으로 완화하기 위해 지방세포에 다양한 자극 을 가하여 지방생성(adipogenesis)을 억제하려는 연구가 계속 진행 중이다[4-6]. 지방생성은 지방전구세포가 분화하면서 지 방조직이 발달하는 현상을 말하며, 세포 내 전사 인자인 CCAAT/enhancer-binding proteins(C/EBPs) 와 peroxisome proliferator-activated receptor gamma(PPAR γ)가 이를 촉 진한다[7, 8]. 또한 apoptosis는 세포예정사(programmed cell death) 라는 방법으로, 원하지 않는 조직을 흔적 없이 제거하는 방법으로 연구되고 있다[9]. Apoptosis를 유발하 는 경로로는 세포막의 수용체를 경유하는 것과 미토콘드리 아에 의해 유도되는 것 두 가지가 존재하는데, 미토콘드리 Corresponding Author : Chi Hyun Kim
Department of Biomedical Engineering, College of Health Sciences, Yonsei University, Wonju, Gangwon-do, 26493, Korea TEL: 033 760 2785
E-mail: [email protected]
이 연구는 교육부의 재원으로 한국연구재단의 지원을 받아 수행된
기초연구사업임(NRF-2015R1D1A1A01060699).
169 아 경로는 B-cell lymphoma 2(Bcl-2)와 Bcl-2-associated
X protein(Bax) 에 의해 조절된다[10].
비만 치료를 목적으로 지방세포의 지방생성과 apoptosis 를 조절하기 위한 연구로 물리적 자극을 이용한 mechano- transduction 이 있다. Mechanotransduction이란 세포에 가해진 물리적 자극이 세포의 수용체를 통해 생화학적 반응 으로 변환되어, 결과적으로 세포의 전사(transcription) 활 동에 영향을 주는 현상을 말하며, 이 세포에는 지방세포 또 한 포함된다[11]. 세포에 mechanotransduction이 일어났을 때 관찰할 수 있는 변화 중의 하나로 섬유아세포(fibroblast) 와 같은 부착 세포 내 액틴(actin)으로 이루어진 stress fiber 의 생성을 들 수 있다[12]. Stress fiber는 세포를 이 동시키고 다른 세포 혹은 기질에 잘 부착할 수 있도록 도울 뿐만 아니라 mechanotransduction을 촉진시키는 역할도 수 행한다[13]. 또한 변환된 생화학적 반응은 세포에 가해진 물 리적 자극의 형태에 따라 각기 다른 신호전달경로(signaling pathway) 를 경유하며, 이에 따라 나타나는 변화 또한 달라 진다[11]. 예를 들어, 지방세포에 압축 하중(compression loading) 이 가해졌을 때 하중의 크기가 일정하게 유지되면 (static) 지방생성이 증가하지만[14], 하중의 크기가 변화하 면(dynamic) 지방생성이 감소한다[15,16]. 따라서 물리적 자극에 대한 세포의 반응을 이해하기 위해서 다양한 방면의 연구가 필요하다.
생체 내 mechanotransduction을 일으키는 물리적 자극 중의 하나로 유체전단응력(fluid shear stress, FSS)이 있 다. 유체에는 점성(viscosity)이 있어 마찰력처럼 흐름에 저 항하려는 성질이 있는데, 이러한 성질에 의해 유체가 압력 을 받아 흐를 때 유체의 점성에 의해 벽면에 평행한 방향으 로 전단응력이 부여된다[17]. 이 현상은 유체로 가득 차 있 고 지속적으로 압력을 받는 조직에서 일어나며, 지금까지도 mechanotransduction 을 연구하기 위하여 이러한 환경을 모방한 in vitro 실험이 진행 중이다[18-20]. 신체의 하중 을 감당하는 대표적인 기관인 뼈 조직에 대해서는 유체전단 응력의 영향이 많이 연구되었으나[18] 지방세포에 대해서는 미비한 실정이며, 다만 다른 물리적 자극을 통해 그 영향을 예상할 수 있었다.
우리는 효과적인 비만 치료법을 제시하기 위하여 유체전단응 력을 3T3-L1 지방전구세포에 직접 가해주어 어떠한 변화가 일 어나는지 측정하였다. 면역형광염색법(immunofluorescence) 을 이용하여 stress fiber의 발달을 관찰하였으며 웨스턴 블롯 (western blot) 을 이용하여 지방생성과 apoptosis 관련 인자 의 변화를 살펴보았다. 또한 유체전단응력을 각각 1시간, 2 시간 동안 가해주어 자극시간에 따른 차이가 있는지 실험하 였다.
II. 실험 방법
1. 세포 배양
세포는 in vitro 환경에서 지방세포 실험을 수행하기 위 해 한국세포주은행에서 배양받은 3T3-L1 preadipocyte를 사 용하였고, confluence가 80% 이상 되었을 때 subculture를 수 행하였다. 배양 배지는 Dulbecco’s modified eagle’s medium (DMEM) 에 calf serum(CS) 5%, penicillin/streptomycin 1% 를 배합하여 사용하였다. 배지 교환은 subculture를 수 행한지 3일 후에 실시하였다. 배양 환경은 37
oC 에 이산화 탄소 5% 농도로 고정하였다.
2. 유체전단응력 실험
Slide 와 도구는 멸균하여 사용하였다. Dish에서 배양한 3T3-L1 세포를 trypsin을 이용하여 떼어준 뒤 slide에 6 × 10
5개씩 옮겨 배양하였으며, 세포들이 slide에 충분히 배양 되었을 때 유체전단응력을 부여하였다. 실험 조건은 유체전 단응력을 세포에 적용한 이전 연구들을 참고하여 진동수 1 Hz, 압력 1 Pa의 조건으로 각각 1시간, 2시간 동안 유체 전단응력을 부여하였다[21-23]. 전단응력에 사용된 배지는 세포배양에 쓰인 것과 동일한 배지를 사용하였다. 대조군을 포함한 모든 실험군은 실험 종료 후 동시에 harvest하였다.
3. Immunofluorescence
세포의 형태 분석을 위하여 immunofluorescence를 실시 하였다. 세포의 actin은 rhodamine-phalloidin으로 염색하 였고, 세포의 핵은 4’-6-Diamidini-2-phenylindole (DAPI) 로 염색하였다. 유체전단응력 자극 후, slide를 PBS로 세척 하고 상온에서 10% 포르말린으로 1시간 동안 고정하였다.
고정한 세포의 actin을 phalloidin(1:200)으로 1시간 동안 염색하였고 PBS로 3회 세척하였다. 이후 세포의 핵을 DAPI(1:1000) 로 3분 동안 염색하고 PBS로 3회 세척한 후 에 형광현미경으로 관찰하였다 (배율 ×200).
4. 단백질 추출 및 western blot
Harvest 한 세포들을 RIPA buffer와 1:1의 부피로 섞은
뒤 1시간 동안 얼음 위에 두어 단백질을 추출하였다. 추출
한 단백질들은 BCA protein assay kit을 이용하여 농도를
측정하였다. SDS-PAGE 시 gel의 농도는 running gel
9%, stacking gel 5% 로 설정하여, 80 V의 전압으로 2시간
동안 전기영동을 진행하였다. Transfer는 100 V의 전압에
서 50분 동안 실시하였다. 그 후 shaker를 이용하여, 단백
질이 transfer된 membrane을 1시간 동안 5% skim milk로
blocking 하였다. 1차 anti body는 1:1000의 비율로 1% BSA
solution 에 희석하여 사용하였고, shaker에서 overnight으로
170
배양하였다. 2차 anti body는 1:5000의 비율로 5% skim milk 에 희석하여 사용하였고, shaker에서 1시간 배양하였 다. Anti body 처리 시, 각 과정 사이에는 TBS-T 용액을 사용하여 10분씩 네 번, 총 40분 간 membrane을 shaker 로 세척하였다.
5. 통계 검정
Western blot 의 결과로서 검출된 band의 밝기 분석은, Image J 프로그램을 이용하여 측정하였다. 측정된 값은 loading control 로 normalize한 뒤, Graphpad Prism 5 프로그램을 이용하여 그래프를 산출, one-way ANOVA를 수행하였다. 결과의 값은 평균 ± 표준편차(SD)로 표현하였
고, 사후 검정은 Tukey’s test를 사용하였다. 통계적 유의 수준은 0.05로 설정하였다.
III. 결 과
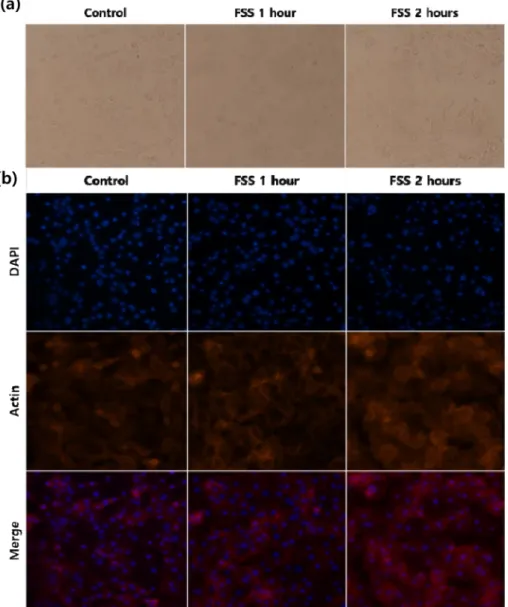
대조군과 유체전단응력을 각각 1시간, 2시간 부여해 준 3T3-L1 세포에 bright field 사진을 촬영하여 비교하였다 ( 그림 1a). 그 결과 유체전단응력을 가해줄수록 세포들의 겉 모습이 둥근 모양에서 넓적하게 퍼져있는 모습으로 변화한 것을 알 수 있었다. Immunofluorescence 사진을 비교했을 때, DAPI로 염색된 핵의 개수는 유체전단응력을 가해줄수 록 감소한 반면 붉은 색으로 염색된 actin의 수는 오히려
그림 1. 3T3-L1에 유체전단응력 실험을 실시했을 때 대조군과 각 실험군 세포의 현미경 사진. (a) Bright field 사진, (b) immunofluorescence 사진. (배율 ×200).
Fig. 1. Microscopy images of each 3T3-L1 groups treated with fluid shear stress. (a) Bright field, (b) immunofluorescence.
Using a microscope with a magnification ×200.
증가하였다(그림 1b). 171
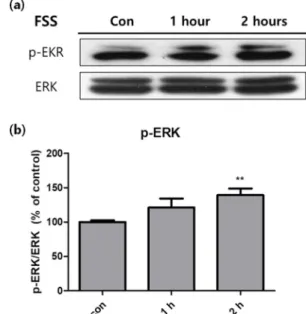
또한 western blot analysis를 이용하여 유체전단응력에 의한 세포 활성의 변화를 분석하였다. 그 결과 세포 내 신 호전달경로 중 하나인 ERK pathway가 시간에 따라 활성 화되는 것을 알 수 있었다(그림 2a). 특히 유체전단응력을 2 시간 가해주었을 때 활성도가 유의하게 증가하였다(그림 2b).
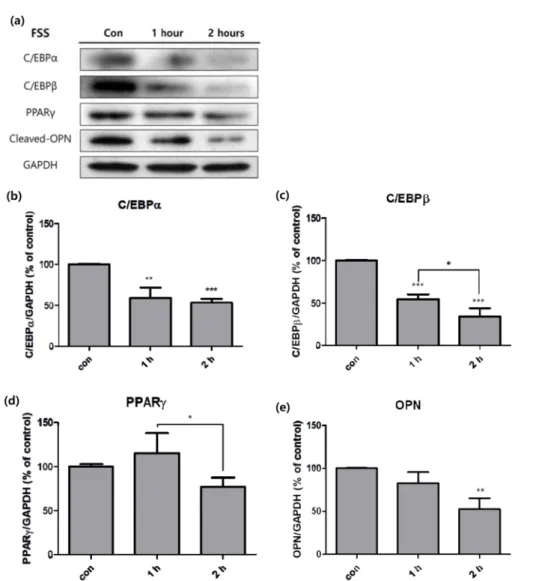
지방생성 인자의 경우, 대체로 자극에 의하여 억제되는 경 향을 보였다(그림 3a). 지방생성의 주요 인자인 C/EBPα와 C/EBPβ 모두 유체전단응력에 의해 크게 감소하였으며(그 림 3b, 3c), 특히 C/EBPβ는 1시간과 2시간 실험군 사이에 서도 유의한 변화를 보였다(그림 3c). 지방생성의 최종 인자 인 PPARγ는 두 실험군과 대조군 간에는 통계적으로 유의 한 변화를 보이지 않았으나, 1시간과 2시간 사이에는 변화가 발견되었다(그림 3d). 비만환자의 지방조직에서 cleaved되어 염증을 유발하는 osteopontin(OPN)의 경우 역시 2시간 실 험군에서 활성이 크게 감소하였다(그림 3e).
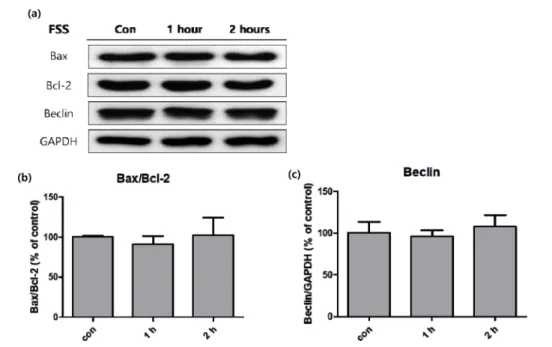
반면에 apoptosis와 관련된 인자들은 유체전단응력에 의 한 큰 변화를 보이지 않았다(그림 4a). 세포의 미토콘드리 아에 의한 apoptosis를 조절하는 Bax와 Bcl-2의 비율을 비 교해 보았을 때, 대조군과 실험군 사이에 유의한 변화가 없 었다(그림 4b). 또한 apoptosis와 구분되는 자가포식 (autophagy) 을 조절하는 Beclin 또한 유의한 변화를 보이 지 않았다(그림 4c).
IV. 고찰 및 결론
지방세포는 물리적 자극에 대한 수용체를 가진 세포로서 외부 자극에 반응하여 유연하게 그 특성을 변화시킨다[6, 11]. 본 실험은 이러한 지방세포의 특성에 집중하여, in vitro 환경에서 3T3-L1 지방전구세포에 일시적인 유체전단 응력을 부여하여 mechanotransduction을 유발, actin stress fiber 의 발달을 비교하였고, 관련 signaling pathway 의 활성과 지방생성, 세포사와 관련된 전사 인자들을 분석하 였다. 자극의 부여 시간은 fibroblast cell line을 대상으로 유 체전단응력 실험을 실시하여 mechanotransduction을 유 도한 이전 연구를 참고하여[21-23] 각각 1시간, 2시간으로 설정하였으며, 이 자극에 의한 세포의 변화를 토대로 물리 적 자극을 통한 비만 치료의 가능성을 제시하였다.
대조군과 1시간, 2시간 실험군의 세포를 bright field 사 진으로 비교해 보았을 때, 대조군에 비해 자극을 가해 준 세 포들이 더 넓은 모양으로 퍼져 있는 것을 관찰할 수 있었다 ( 그림 1a). Immunofluorescence 촬영 사진 또한 자극의 시간 이 길어질수록 actin이 더 진하게 염색되었다(그림 1b). Actin 으로 이루어진 stress fiber는 세포가 mechanotransduction이 일어날 때 발달하는 것으로 알려져 있으며, 섬유아세포와 같 은 부착세포에서 focal adhesion을 일으켜 세포가 다른 세포 나 기질에 접착할 수 있도록 돕는다[12,13]. 이 사실을 종합해 볼 때, 3T3-L1이 유체전단응력을 받아 mechanotransduction 이 발생했고, 세포가 유체의 흐름에 적응할 수 있도록 stress fiber가 focal adhesion을 제공, 넓게 퍼진 형태로 slide glass 에 접착된 것으로 보인다. 또한 이전 연구에서 섬유아 세포의 분화에 stress fiber의 활성화가 중요한 역할을 하 는 것으로 알려져 있는데[13,24,25], 이 세포가 지방세포로 분화하기 위해서 stress fiber의 생성이 억제되어야 하므로 반대로 물리적 자극에 의한 stress fiber의 생성은 지방생 성을 억제하는 작용과 연관이 있는 것으로 보인다[26,27].
실제로 3T3-L1에 피부괴사독소를 투여한 실험에서 지방생 성의 억제와 stress fiber의 활성화가 동시에 관찰된 실험 도 있었다[25].
지방세포의 mechanotransduction은 물리적 자극의 형태와 성질에 따라 경유하는 signaling pathway가 다르다[11]. 따라서 더 다양한 자극에 대한 지방세포의 mechanotransduction을 연구하기 위해 유체전단응력에 의해 활성화 되는 지방세포 의 signaling pathway를 다른 자극들과 비교해보았다.
Western blot 의 결과를 분석했을 때, 유체전단응력에 의한 물리적 자극은 ERK pathway를 통해 화학적 신호로 변환 되어 세포의 전사 활동에 영향을 준 것으로 보인다(그림 2a).
현재 알려진 지방세포의 mechanotransduction 중에서 ERK pathway 를 경유하는 자극으로는 장력의 크기와 방향
그림 2. 유체전단응력을 부여한 3T3-L1에 western blot을 실시하여 대조군 대비 세포의 ERK pathway 활성화 정도를 분석한 것.
(a) Western blot 결과, (b) p-ERK (n=3, **P<0.01).
Fig. 2. Activity of signaling pathway in 3T3-L1 cells measured with western blot analysis, after fluid shear stress was treated. (a) Bands of western blot analysis, (b) p- ERK (n=3, **P<0.01).
172
이 주기적으로 변화하는 cyclic stretching이 있다[15,16]. 따 라서 지방세포에 부여되는 유체전단응력은 cyclic stretching 과 유사한 종류의 자극으로 생각된다. 추가적인 실험으로 smad2, COX-2 등 물리적 자극을 통해 지방생성을 억제하 는 다른 signaling pathway를 분석하고 지방세포에 적용 가능한 다른 자극들과 비교하여 몸의 구조체로서 지방조직 의 기능을 추후 보다 심도있게 규명하고자한다[28,29].
또한 유체전단응력에 의해 지방세포의 전사 인자인 C/
EBPs 가 시간에 따라 크게 감소하여 지방생성이 억제된 것 을 알 수 있었는데(그림 3b, 3c), 이 또한 cyclic stretching 으로 인한 결과와 유사하다. 반면에 PPARγ는 대조군과 실 험군이 유의한 차이를 보이지 않았지만 1시간 실험군과 2 시간 실험군 사이에는 유의한 차이가 존재하였다(그림 3d).
이는 지방생성의 최종 인자인 PPARγ이 활성화 되기에는 시 간이 충분하지 않았기 때문으로 예상할 수 있다.
한편 OPN 또한 자극에 의해 큰 감소를 보였다(그림 3e).
지방조직에서 OPN은 대식세포의 활성을 조절하여 염증을 일으키는 역할을 하며[30], 비만 환자의 지방조직에서 많이 분비되는 것으로 알려져 있다[31]. 따라서 지방조직 내 유 체전단응력을 효과적으로 일으킬 수 있다면 지방조직의 비 대화와 같은 염증으로 인한 비만 환자의 증상을 크게 완화 시킬 수 있을 것이다.
반면에 미토콘드리아에 의한 apoptosis 관련 인자인 Bcl- 2 와 Bax, autophagy 전사 인자인 Beclin은 모두 유의한 변 화를 보이지 않았다(그림 4). 생화학적 자극을 이용하여 지 방세포의 지방생성 억제와 apoptosis를 동시에 유발하려는
그림 3. 유체전단응력을 부여한 3T3-L1에 western blot을 실시하여 대조군 대비 세포의 지방생성 관련 인자들의 활성화 정도를 분석한 것.(a) Western blot 결과, (b) C/EBPα, (c) C/EBPβ, (d) PPAR
γ
, (e) OPN (n=3, *P<0.05, **P<0.01, ***P<0.001).Fig. 3. Activity of adipogenic factors in 3T3-L1 cells measured with western blot analysis, after fluid shear stress was treated. (a) Bands of western blot analysis, (b) C/EBPα, (c) C/EBPβ, (d) PPAR
γ
, (e) OPN (n=3, *P<0.05, **P<0.01,***P<0.001).
실험은 이전까지 연구된 바 있지만[4,5], 물리적 자극을 이 173 용하여 apoptosis를 유도하였다는 보고는 미비하다. 신체의 항상성 유지의 측면에서 생각해 볼 때, 생체 조직에 가해지 는 물리적 자극은 생화학적 자극에 비해 운동과 같은 일상 적인 행동으로 쉽게 유발된다. 그러므로 물리적 자극에 의 한 apoptosis, autophagy 등의 세포의 직접적인 수적 변 화는 적을 것으로 보인다. 따라서 그림 1b의 형광 사진에서 DAPI 로 염색된 핵의 수가 자극에 따라 감소한 것은 세포 가 사멸한 것이 아니라, 유체전단응력에 의하여 일부 세포 가 slide에서 떨어져 나간 것으로 사료된다.
결론적으로, 유체전단응력은 mechanotransduction을 통한 3T3-L1 지방전구세포의 지방생성 억제에 탁월한 효과 를 보였으며 여기에 actin stress fiber의 생성, ERK pathway 의 활성화가 관여한 것을 알 수 있었다. 또한 유체 전단응력에 의한 영향은 시간에 따라 변화하였다. 앞으로 지 방세포의 mechanotransduction에 대한 연구가 지속되어, 이에 대한 이해를 바탕으로 유체전단응력을 포함한 물리적 자극을 지방조직에 효율적으로 일으킬 수 있다면, 비만의 주 요 증상인 지방조직의 과잉발달을 효과적으로 치료할 수 있 을 것으로 기대된다.
참고문헌
[1] B.M. Spiegelman and J.S. Flier, “Obesity and the regulation
of energy balance,” cell, vol. 104, no. 4, pp. 531-543, 2001.
[2] X. Formiguera and A. Cantón, “Obesity: epidemiology and clinical aspects,” Best Pract. Res. Clin. Gastroenterol., vol.
18, no. 6, pp. 1125-1146, 2004.
[3] C. Weyer, J.E. Foley, C. Bogardus, P.A. Tataranni, and R.E.
Pratley, “Enlarged subcutaneous abdominal adipocyte size, but not obesity itself, predicts type II diabetes independent of insulin resistance,” Diabetologia, vol. 43, no. 12, pp. 1498- 1506, 2000.
[4] Y.W. Wang and P.J.H. Jones, “Conjugated linoleic acid and obesity control: efficacy and mechanisms,” Int. J. Obes., vol.
28, no. 8, pp. 941-955, 2004.
[5] C.L. Hsu and G.C. Yen, “Effects of capsaicin on induction of apoptosis and inhibition of adipogenesis in 3T3-L1 cells,” J.
Agric. Food Chem., vol. 55, no. 5, pp. 1730-1736, 2007.
[6] D. Hwang, S. Kim, H. Lee, S. Lee, D. Seo, S. Cho, S. Chen, T. Han, and H.S. Kim, “The Effects of Whole Body Vibra- tion in the Aspect of Reducing Abdominal Adipose Tissue in High-Fat Diet Mice Model,” J. Biomed. Eng. Res., vol. 38, no. 1, pp. 49-55, 2017.
[7] M.I. Lefterova, and M.A. Lazar, “New developments in adi- pogenesis,” Trends Endocrinol. Metab., vol. 20, no. 3, pp.
107-114, 2009.
[8] U.A. White, and J.M. Stephens, “Transcriptional factors that promote formation of white adipose tissue,” Mol. Cell.
Endocrinol., vol. 318, no. 1-2, pp. 10-14, 2010.
[9] K. Vermeulen, D.R.V Bockstaele, and Z.N. Berneman,
“Apoptosis: mechanisms and relevance in cancer,” Ann.
Hematol., vol. 84, no.10, pp. 627-639, 2005.
[10] M.O. Hengartner, “The biochemistry of apoptosis,” Nature, vol. 407, pp. 770-776, 2000.
[11] N. Shoham, and A. Gefen, “Mechanotransduction in adipo- 그림 4. 유체전단응력을 부여한 3T3-L1에 western blot을 실시하여 대조군 대비 세포의 세포사 관련 인자들의 활성화 정도를 분석한 것.
(a) Western blot 결과, (b) Bax와 Bcl-2의 비율, (c) Beclin (n=3).
Fig. 4. Activity of programmed cell death factors in 3T3-L1 cells measured with western blot analysis, after fluid shear stress was treated. (a) Bands of western blot analysis, (b) ratio of Bax to Bcl-2, (c) Beclin (n=3).
174
cytes,” J. Biomech., vol. 45, no. 1, pp. 1-8, 2012.
[12] R. McBeath, D.M. Pirone, C.M. Nelson, K. Bhadriraju, and C.S. Chen, “Cell shape, cytoskeletal tension, and RhoA reg- ulate stem cell lineage commitment,” Dev. Cell, vol. 6, no. 4, pp. 483-495, 2004.
[13] S. Tojkander, G. Gateva, and P. Lappalainen, “Actin stress fibers-assembly, dynamics and biological roles,” J. Cell Sci., vol. 125, no. 8, pp. 1855-1864, 2012.
[14] Y. Hara, S. Wakino, Y. Tanabe, M. Saito, H. Tokuyama, N.
Washida, S. Tatematsu, K. Yoshioka, K. Homma, K. Hase- gawa, H. Minakuchi, K. Fujimura, K. Hosoya, K. Hayashi, K. Nakayama, and H. Itoh, "Rho and Rho-kinase activity in adipocytes contributes to a vicious cycle in obesity that may involve mechanical stretch," Sci. Signal., vol. 4, no. 157, pp.
ra3, 2011.
[15] Y. Tanabe, M. Koga, M. Saito, Y. Matsunaga, and K.
Nakayama, “Inhibition of adipocyte differentiation by mechan- ical stretching through ERK-mediated downregulation of PPARγ2,” J. Cell. Sci., vol. 117, no. 16, pp. 3605-3614, 2004.
[16] Y. Tanabe, Y. Matsunaga, M. Saito, and K. Nakayama,
“Involvement of cyclooxygenase-2 in synergistic effect of cyclic stretching and eicosapentaenoic acid on adipocyte dif- ferentiation,” J. Pharmacol. Sci., vol. 106, no. 3, pp. 478-484, 2008.
[17] M.H. Kroll, J.D. Hellums, L.y V. Mclntire, A.I. Schafer, and J.L. Moake, “Platelets and shear stress,” Blood, vol. 88, no .5, pp. 1525-1541, 1996.
[18] Y. Li, J. Yuan, Q. Wang, L. Sun, Y. Sha, Y. Li, L. Wang, and Z. Wang, “The collective influence of 1, 25-dihydroxyvita- min D3 with physiological fluid shear stress on osteoblasts,”
Steroids, vol. 129, pp. 9-16, 2016.
[19] P. Wang, P.P. Guan, C. Guo, F. Zhu, K. Konstantopoulos, and Z.Y. Wang, “Fluid shear stress-induced osteoarthritis: roles of cyclooxygenase-2 and its metabolic products in inducing the expression of proinflammatory cytokines and matrix metalloproteinases,” FASEB J., vol. 27, no. 12, pp. 4664- 4677, 2013.
[20] T.M. Maul, D.W. Chew, A. Nieponice, and D.A. Vorp,
“Mechanical stimuli differentially control stem cell behavior:
morphology, proliferation, and differentiation,” Biomech.
Model. Mechanobiol., vol. 10, no. 6, pp. 939-953, 2011.
[21] H. Huang, R.D. Kamm, and R.T. Lee, “Cell mechanics and mechanotransduction:pathways, probes, and physiology,”
Am. J. Physiol. Cell. Physiol., vol. 287, no. 1, pp. C1-C11, 2004.
[22] C.H. Kim, L. You, C.E. Yellowley, and C.R. Jacobs, “Oscil- latory fluid flow-induced shear stress decreases osteoclasto- genesis through RANKL and OPG signaling,” Bone, vol. 39, no. 5, pp. 1043-1047, 2006.
[23] C.H. Kim, and Y.M. Yoo, “Fluid shear stress and melatonin in combination activate anabolic proteins in MC3T3-E1 osteoblast cells,” J. Pineal Res., vol. 54, no. 4, pp. 453-461, 2013.
[24] K. Kumawat, T. Koopmans, M.H. Menzen, A. Prins, M.
Smit, A.J. Halayko, and R. Gosens, “Cooperative signaling by TGF-β1 and WNT-11 drives sm-α-actin expression in smooth muscle via Rho kinase-actin-MRTF-A signaling,”
Am. J. Physiol. Lung Cell. Mol. Physiol., vol. 311, no. 3, pp.
L529-L537, 2016.
[25] Y. Bannai, L.R. Aminova, M.J. Faulkner, M. Ho, and B.A.
Wilson, “Rho/ROCK-dependent inhibition of 3T3-L1 adipo- genesis by G-protein-deamidating dermonecrotic toxins: dif- ferential regulation of Notch1, Pref1/Dlk1, and β-catenin signaling,” Front. Cell. Infect. Microbiol., vol. 2, pp. 80, 2012.
[26] D.T.V. Diep, K. Hong, T. Khun, M. Zheng, A. Ul-Haq, H.S.
Jun, Y.B. Kim, and K.H. Chun, “Anti-adipogenic effects of KD025 (SLx-2119), a ROCK2-specific inhibitor, in 3T3-L1 cells,” Sci. Rep., vol. 8, no. 1, pp. 2477, 2018.
[27] T. Horii, S. Morita, M. Kimura, and I. Hatada, “Epigenetic regulation of adipocyte differentiation by a Rho guanine nucleotide exchange factor, WGEF,” PLoS One., vol. 4, no.
6, pp. e5809, 2009.
[28] N.J. Turner, H.S. Jones, J.E. Davies, and A.E. Canfield,
“Cyclic stretch-induced TGFβ1/Smad signaling inhibits adi- pogenesis in umbilical cord progenitor cells,” Biochem. Bio- phys. Res. Commun., vol. 377, no. 4, pp. 1147-1151, 2008.
[29] M.G. Hossain, T. Iwata, N. Mizusawa, S.W. Shima, T. Okutsu, K. Ishimoto, and K. Yoshimoto, “Compressive force inhibits adipogenesis hrough COX-2-mediated down-regulation of PPARgamma2 and C/EBPalpha,” J. Biosci. Bioeng., vol.
109, no. 3, pp. 297-303, 2010.
[30] Y.H. Lee, A.P. Petkova, and J.G. Granneman, “Identification of an adipogenic niche for adipose tissue remodeling and res- toration,” Cell metab., vol. 18, no. 3, pp. 355-367, 2013.
[31] M. Zeyda, K. Gollinger, J. Todoric, F.W. Kiefer, M. Keck, O.
Aszmann, G. Prager, G.J. Zlabinger, P. Petzelbauer, and T.M.
stulnig, “Osteopontin is an activator of human adipose tissue macrophages and directly affects adipocyte function,” Endo- crinology, vol. 152, no. 6, pp. 2219-2227, 2011.