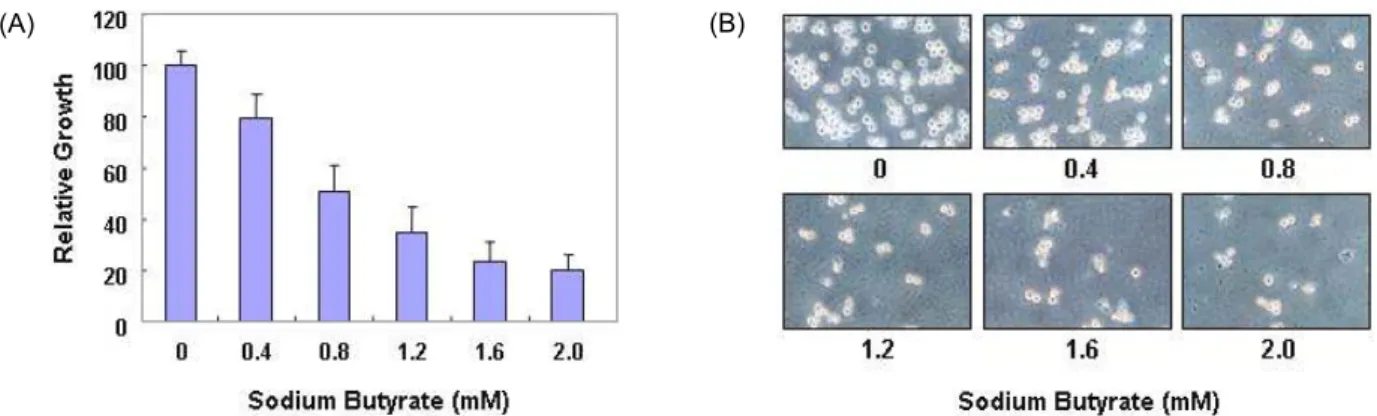
G1 Arrest of U937 Human Monocytic Leukemia Cells by Sodium Butyrate, an HDAC Inhibitor, Via Induction of Cdk Inhibitors and Down-regulation of pRB Phosphorylation
7
0
0
전체 글
(2)
(3)
(4)
(5)
(6)
(7)
수치



관련 문서
In vitro cell migration in APE or JAG1 siRNA-treated M059K cell line Fig,11 Down-regulation of JAG1 induces S phase arrest in
In this study, we investigated the effects of low-power CO 2 laser on proliferation on human gingival fibroblast cells so that determine laser
Fig 4 : Inhibitory effects of classified methanol extracts of Acalypha australis the COX-2 protein expression of the human oral cavity carcinoma KB cells....
4 > Effects of Taro on COX-2 expression and iNOS expression(hot water) in human thyroid cancer cells. The cells were pretreated for 48hours with either
PI3K inhibition decreased antioxidants/GD-induced apoptosis in A549 cells, and PI3K inhibitor LY294002 had inhibitory effect on antioxidants/GD-induced caspase-3
In the present study we investigated the proliferation effect of human oral cancer KB cell treated with pulsatilla koreana extract.. We analyzed the effects of this
In the present study, consistent to that report, up-regulation of VEGF mRNA was observed in the colon cancer cell with the acquired resistance to 5-FU and this
The aim of this study was to examine the effects and mechanisms of the new metformin derivative HL156A in human multidrug-resistant cancer cell lines (FADU/PTX,
