Genetic Analysis of H7N7 Avian Influenza Virus Isolated From Waterfowl in South Korea in 2016
Berihun Dires1,2 and Sang Heui Seo1,2*
1Laboratory of Influenza Research, College of Veterinary Medicine, Chungnam National University, Daejeon 34134, Korea
2Institute of Influenza Virus, College of Veterinary Medicine, Chungnam National University, Daejeon 34134, Korea
Received April 23, 2018 /Revised June 8, 2018 /Accepted June 10, 2018
Type A influenza virus is circulating in wild birds and can infect wide ranges of hosts such as hu- mans, pigs, domestic birds, and other mammals. Many subtypes of avian influenza viruses are circu- lating in aquatic birds. Most avian influenza viruses found in aquatic birds are low pathogenic avian influenza viruses. Highly pathogenic avian influenza viruses have been found in waterfowls since 2005. It is known that H5 and H7 subtypes of avian influenza viruses can be mutated into highly pathogenic avian influenza viruses in domestic poultry. In this study, we isolated novel reassortant H7N7 avian influenza virus from the fecal materials of migratory birds in the Western part of South Korea in 2016, and analyzed the sequences of all its eight genes. The genetic analysis of our isolate, A/waterfowl/Korea/S017/2016 (H7N7) indicates that it was reassortant avian influenza virus contain- ing genes of both avian influenza viruses of wild birds and domestic ducks. Phylogenetic analysis showed that our isolate belongs to Eurasian lineage of avian influenza virus. Since avian influenza viruses continue to evolve, and H7-subtype avian influenza virus can mutate into the highly patho- genic avian influenza viruses, which cause the great threat to humans and animals, we closely survey the infections in both wild birds, and domestic poultry, and mammals.
Key words : H7N7, influenza A virus, South Korea, waterfowl
*Corresponding author
*Tel : +82-42-821-7819, Fax : +82-42-821-6762
*E-mail : [email protected]
This is an Open-Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/3.0) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
Journal of Life Science 2018 Vol. 28. No. 8. 962~968 DOI : https://doi.org/10.5352/JLS.2018.28.8.962
Introduction
Avian influenza viruses belong to the Orthomyxoviridae family. Among four types of influenza viruses, A, B, C and D, influenza A virus can infect a wide range of hosts com- pared to the other three influenza viruses. Both humans and animals can get infected by influenza A Viruses. The natural reservoirs of influenza A viruses are wild aquatic birds such as shorebirds, ducks and geese [4, 10, 19]. Aquatic birds in- fected with many subtypes of avian influenza viruses do not show the clear clinical signs, but they play important role in creation of reassortment of influenza A viruses [3, 13].
The genome of avian influenza viruses consists of eight negative-sense segmented genes, PB2 (RNA polymerase ba- sic subunit 2), PB1 (RNA polymerase basic subunit 1), PA (RNA polymerase acidic subunit), HA (haemagglutinin), NP
(nucleoprotein), NA (neuraminidase), M (Matrix) and NS (nonstructural). The two proteins on the surface of influenza virus, HA and NA are used to divide influenza A viruses into many subtypes. Currently, there are 18 different hemag- glutinin (H1-H18) and 11 different neuraminidase subtypes (N1-N11) in influenza A viruses [4, 19]. Influenza A viruses are also classified into low pathogenic avian influenza virus (LPAI) and highly pathogenic avian influenza virus (HPAI) based on the severity of the disease in the infected animals.
H5 and H7 subtypes of influenza A have been associated with HPAI viruses among 18 HA subtypes, and have the polybasic amino acids such as arginine (R) and lysine (K) in the cleavage sites of HA proteins [4, 20].
H7N7-subtype influenza A virus was first detected in chickens in Italy in 1902 [8]. Since then, it has been isolated from domestic poultry and wild birds around the world [2, 18]. The first human case of H7N7 influenza A virus was found in England in 1996 [12]. Three cases of human con- junctivitis in humans infected with H7N7 influenza A virus infection were reported during the 2013 outbreak of H7N7 HPAI virus in Italy [1]. H7N7 influenza A viruses were also isolated from magpie in northwestern part of South Korea [11] and live poultry markets in China [6]. Owing to their continued detection in birds and their ability to cause human
infection, H7 subtypes of influenza viruses are one of the public health threats [1, 15]. In this study, we isolated H7N7 influenza virus from the fecal samples of wild birds in South Korea in winter, 2016, and genetically characterized its genome.
Materials and Methods
Virus Isolation
About 100 fecal samples of migratory birds were collected in migratory bird site, Chunsu Man which is located in the Western part of South Korea, 2016. This area is known to attract migratory birds during winter season. Isolation me- dia containing glycerol (50%) was used to collect fecal samples. The samples were then inoculated into 10-day-old embryonated specific pathogenic free (SPF) eggs. The in- oculated eggs were incubated for 72 hr at 37℃ under humid conditions and then were chilled overnight at 4℃. Presence of influenza A virus in allantoic fluid was checked by he- magglutination assay (HA) with 0.5% turkey red blood cells in phosphate buffered solution (PBS, pH 7.4). We designated the isolate as A/waterfowl/Korea/S017/2016 (H7N7).
RNA extraction and PCR amplification
Viral RNAs from the HA-positive allantoic fluid were ex- tracted using RNeasy protect Mini kit (Qiagen, CA, USA) in accordance with the manufacturer’s instructions. Extracted viral RNAs were used to determine genetic information of the isolate. Uni-12 primer (5’-AGCAAAAGCAGG-3’) was used to reverse transcribe viral RNA to complementary DNA (cDNA). Eight viral genes of the isolate were amplified by Polymerase chain (PCR) with GoTaq® DNA polymerase and a segment specific primer set which are available up on request. The primers used for amplifying the genes were;
forward (5’TCCGAAGTTGGGGGGGAGCGAAAGCA GGTC-3’) and reverse (5’-GGGCCGCCGGGTTATTAGTAG AAACAA GGTCG-3’) for PB2, forward (5’-TCCGAAGTTGGGGGG GAGCGAAAGCAGGCA-3’) and reverse (5’-GGGCCGCCG GGTTATTAGTAGAAACAAGGCAT3’) for PB1, forward (5’
TCCGAAGTTGGGGGGGAGCGAAAGCAGGTA3’) and re- verse (5’GGGCCGCCGGGTTATTAGTAGAAACAAGGTAC3’) for PA, forward (5’TCCGAAGTTGGGGGGGAGCAAAAGC AGGGG3’) and reverse (5’GGGCCGCCGGGTTATTAGTAG AAACAAGGGTG3’) for HA, forward (5’TCCGAAGTTGGG GGGGAGCAAAAGCAGGGT3’) and reverse (5’GGGCCGC CGGGTTATTAGTAGAAACAAGGGTA3’) for NP, forward
(5’TCCGAAGTTGGGGGGGAGCAAAAGC AGGAG3’) and reverse (5’GGGCCGCCGGGTTATTAGTAGAAACAAGGA GT3’) for NA, forward (5’TCCGAAGTTGGGGGGGAGCAA AAGCAGGTA3’) and reverse (5’GGGCCGCCGGGTTATTA GTAGAAACAAGGTAG3’) for M, and forward (5’TCCGA AGTTGGGGGGGAGCAAAAGCAGGGT3’) and reverse (5’GG GCCGCCGGGTTATTAGTAGAAACAAG GG TG-3’) for NS. Gel electrophoresis was used to separate amplicons and then the bands were purified using QIAquick Gel Extraction Kit (Qiagen, CA, USA) after they were excised from the gel.
Sequencing and phylogenetic analysis
TA vector (Promega, USA) was used to clone the purified DNA. Transformation was carried out using Escherichia coli competent cell, X- gal indicator and Luria Bertani (LB) me- dium by incubating it overnight at 37℃. The white colonies were selected and grown in LB broth overnight at 37℃ in shaking incubator. Plasmid extractions were conducted and were sent for sequencing. Sequencing of genes was per- formed by Macrogen (Daejeon, South Korea). Seqman model of DNA star package was used to edit the nucleotide sequences. The nucleotide sequences were deposited into GenBank under accession numbers, MH169353-MH169360.
Phylogenetic analysis was carried out by using molecular evolutionary genetic analysis software MEGA 7 (MEGA 7.0) with the maximum-likelihood method. Bootstrap analysis using 1,000 replicates was used to evaluate the reliability of the phylogenetic tree. The input nucleotide sequence in- cluded both the isolate and published influenza virus se- quences from GenBank database.
Results and Discussion
Homology comparison of the isolate, A/Waterfowl/
Korea/S017/2016 (H7N7)
The closest related influenza viruses to our isolate, A/wa- terfowl/Korea/S017/2016(H7N7) were examined using BLAST (basic local alignment search tool) (http://blast.ncbi.nlm.
nih.gov/Blast.cgi,gisaid.org/epi3/frontend#4ead5c ) (Table 1). The data in Genbank indicated the NP, M and NS genes were closely related to those of the A/duck/Ganzhou/
G25/2015(H4N6), A/duck/Mongolia/543/2015(H4N6) and A/wild duck/Korea/SH5-26/2008(H4N6) with 99% identity respectively. PB1, PA, and HA were closely related to the A/Anseriformes/Anhui/L167/2014(H1N1), A/duck/Mon- golia/83/2015(H3N6) and A/duck/Bangladesh/26992/2015
Table 1. Nucleotide homology of A/Waterfowl/Korea/S017/2016(H7N7) with the closest related influenza viruses
Viral genes Closest related viruses Nucleotide identity (%)
PB2 PB1 PA HA NP NA M NS
A/barnacle goose/Netherlands/2/2014(H3N6), A/Anseriformes/Anhui/L167/2014(H1N1)
A/duck/Mongolia/83/2015(H3N6) A/duck/Bangladesh/26992/2015(H7N9)
A/duck/Ganzhou/G25/2015(H4N6) A/duck/Mongolia/742/2015(H10N7) A/duck/Mongolia/543/2015(H4N6) A/wild duck/Korea/SH5-26/2008(H4N6)
99 98 98 98 99 99 99 99
Table 2. Identification of amino acids of A/Waterfowl/Korea/S017/2016(H7N7) involved in binding to human-type influenza re- ceptor, enhancing antiviral drug resistance, and causing pathogenesis in mammal [6, 7, 9, 14, 17]
Viral protein Amino acid position
A/Waterfowl/Korea/
S017/2016(H7N7) Comments
HA (H3 numbering)
138 226 228
A Q G
A138S: Increased binding to human Q226L: Increased binding to human G228S: Increased binding to human NA
(N2 numbering)
152 274 292
R H R
R152K: Resistance to oseltamivir and zanamivir H274Y: Resistance to oseltamivir and zanamivir R292K: Resistance to oseltamivir and zanamivir M1
M2 NS1
215 31 42
A S S
T215A: Increased pathogenesis in mice
S31N: Resistance to amantadine and remantadine P42S: Increased pathogenesis in mice
PB2: RNA polymerase basic subunit 2; PB1: RNA polymerase basic subunit 1;HA, haemagglutinin; NA: neuraminidase; M1:
Matrix gene 1; M2: Matrix gene 2; NS1: non-structural gene 1.
A: Alanine; D: Aspartic acid E: Glutamic acid; G: Glycine; H: Histidine; K: Lysine; L: Leucine; N: Asparagine; P: Proline; Q:
Glutamine; R: Arginine; S: Serine; T: Threonine, V: Valine; Y: Tyrosine.
(H7N9) with 98% identity respectively. PB2 and NA were closely related to the A/barnacle goose/Netherlands/
2/2014(H3N6) and A/duck/Mongolia/742/2015(H10N7) with 99% identity respectively.
Genetic analysis showed that our isolate, A/Waterfowl/
Korea/S017/2016(H7N7) is a reassorted avian influenza vi- rus between the origins of domestic ducks and wild birds.
PB2 and NS genes were from the origin of wild birds, while the rest of genes, PB1, PA, HA, NP, NA, and M were from the origin of domestic ducks (Table 1).
Our genetic analysis showed that the isolate was a re- assorted avian influenza virus using viral genes of domestic ducks and wild birds. It suggests that migratory birds make frequent contacts with domestic poultry, resulting in the cre- ation of novel avian influenza virus. To prevent the out- breaks of new subtypes of influenza viruses in the domestic poultry and wild birds, it is recommended to set up a barrier to prevent the contact between wild birds and domestic poultry. Since H7-subtypes of influenza viruses can mutate
into a highly pathogenic avian influenza virus in poultry [16], the intensive surveillance in poultry is very important to minimize the economic losses in poultry industry.
Analysis of important amino acids involved in binding to human-type influenza receptor, enhancing antiviral drug resistance, and causing pathogenesis in mammal of the isolate, A/Waterfowl/Korea/S017/
2016(H7N7)
To know the characteristics of the isolate more, the se- lected regions of the A/Waterfowl/Korea/S017/2016(H7N7) genome were further analyzed (Table 2). Our isolate had ala- nine (A) at position 138, glutamine (Q) at position 226, and glycine (G) at position 228 of the HA protein, suggesting the efficient binding to avian influenza receptors [17].
Analysis of the NA gene of this isolate revealed that there was arginine (R) at positions 152 and 292, and histidine (H) at position 274, suggesting the susceptibility to the NA in- hibitors, oseltamivir and zanamivir [7]. The M1 mutation T
A/duck/Mongolia/129/2010(H7N9) A/ruddy shelduck/Mongolia/436/2010(H7N1) A/ruddy shelduck/Mongolia/1009/2010(H7N7) A/ruddy shelduck/Mongolia/1189/2010(H7N1)
A/mallard/Korea/822/2010(H7N7) A/common goldeneye/Mongolia/1271/2010(H7N1)
A/mallard/Republic of Georgia/3/2010(H7N3)) A/mallard/Republic of Georgia/3/2010(H7N3) A/duck/Thailand/CU-9744C /2010(H7N4) A/wild duck/Korea/SH19-47/2010(H7N9) A/wild duck/Korea/MHC35-25/2011(H7N9) A/ostrich/South Africa/ORD/2012(H7N1)
A/ostrich/South Africa/KVL/2012(H7N1) A/shellduck/South Africa/DLH/2012(H7N8) A/sacred ibis/South Africa/120701/2012(H7N1) A/ostrich/South Africa/FWS/2012(H7N1) A/ostrich/South Africa/MKT/2012(H7N1) A/northern shoveler/Egypt/EMC-1/2012(H7N1) A/duck/Bangladesh/27042/2015(H7N9) A/duck/Bangladesh/26992/2015(H7N9) A/Waterfowl/Korea/S017/2016(H7N7)
A/mallard/New York/A00723392/2009(H7N3) A/environment/Indiana/08OS2857/2008(H7N3)
A/mallard/MN/AI07-4315/2007(H7N3) A/mallard/MN/AI07-3302/2007(H7N8)
A/northern shoveler/Washington/44249-664/2006(H7N3) A/American green-winged teal/California/44242-906/2007(H7N3) A/green-winged teal/TX/1254/2007(H7N3)
A/blue-winged teal/TX/854/2007(H7N7) 81
99
69 81 80 100
52 99 31
93 87
97 97 63 99
99
6361 74
68
99
0.050
HA
Eurasian
North American (D)
A/duck/Mongolia/83/2015(H3N8) A/duck/Mongolia/777/2015(H3N8) A/duck/Mongolia/211/2015(H3N8) A/duck/Mongolia/167/2015(H3N8) A/duck/Mongolia/208/2015(H3N8) A/duck/Mongolia/30/2015(H3N8) A/duck/Mongolia/655/2015(H2N3) A/duck/Mongolia/742/2015(H10N7) A/duck/Mongolia/482/2015(H2N3) A/duck/Mongolia/518/2015(H10N3)
A/Waterfowl/Korea/S017/2016(H7N7) A/mallard/Republic of Georgia/13/2011(H6N2)
A/duck/Mongolia/124/2015(H3N2) A/wild goose/Dongting/C1037/2011(H12N8) A/black-tailed godwit/Bangladesh/24734/2015(H7N5)
A/duck/Bangladesh/24704/2015(H15N9) A/duck/Bangladesh/25891/2015(H4N6) A/duck/Bangladesh/24035/2014(H10N1) A/mallard duck/Netherlands/20/2012(H1N1) A/mallard duck/Netherlands/10/2012(H6N1) A/mallard duck/Netherlands/37/2011(H1N1) A/ruddy turnstone/Delaware Bay/135/1996(H7N3) A/ruddy turnstone/Delaware Bay/116/1996(mixed)
A/mallard/Alberta/167/1998(H3N8) A/chicken/NY/14714-2/1999(H7N3) A/pintail/Alberta/140/1996(H1N1) A/lesser scaup/Alberta/206/1995(H3N8))
A/mallard/Alberta/114/1997(H3N8) A/mallard/ALB/47/1998(H4N1) 84
27 15 100
21 100
100 53 61 97 61 57
40 100
56
99 71
98 99
83 75
0.0100
PA (C)
Eurasian
North America
A/duck/Mongolia/655/2015(H2N3) A/duck/Mongolia/482/2015(H2N3) A/duck/Mongolia/742/2015(H10N7) A/duck/Mongolia/200/2015(H3N8) A/common shelduck/Mongolia/2106/2011(H3N8) A/duck/Mongolia/518/2015(H10N3)
A/duck/Mongolia/572/2015(H3N8) A/duck/Vietnam/LBM533/2013(H3N6)
A/duck/Bangladesh/24697/2015(H15N9) A/duck/Bangladesh/24704/2015(H15N9) A/mallard duck/Netherlands/2/2013(H3N6) A/teal/Djankoy/4-17-11/2010(H5N2) A/mallard/Republic of Georgia/15/2011(H10N7)
A/Waterfowl/Korea/S017/2016(H7N7) A/Anseriformes/Anhui/L167/2014(H1N1) A/wild bird/Wuhan/CDHN09/2015(H6N2) A/wild bird/Wuhan/CDHN01/2015(H11N9) A/wild bird/Wuhan/CDHN22/2015(H11N9) A/wild bird/Wuhan/CDHN173/2015(H11N9) A/duck/Bangladesh/26918/2015(H3N6)
A/duck/Mongolia/179/2015(H3N8)
A/swine/North Carolina/18161/2002(H1N1) A/swine/North Carolina/47834/2000(H1N1) A/swine/Texas/4199-2/1998(H3N2)
A/swine/Minnesota/1192/2001(H1N2) A/swine/Mexico/Ver29/2010(H1N1) A/Wisconsin/10/98 (H1N1) A/swine/Mexico/Mex51/2010(H3N2) A/swine/Mexico/Mex50/2010(H3N2) 99
48 26 43 100 98
100
88 98 78 71 99 100
64 82 72
60 91 72 35
99
0.020
PB1 (B)
Eurasian
North American
A/duck/Jiangxi/33653/2013(H10N3) A/duck/Jiangxi/33629/2013(H10N3) A/duck/Jiangxi/33747/2013(H10N3) A/duck/Jiangxi/33637/2013(H10N3) A/duck/Jiangxi/33757/2013(H10N3) A/duck/Jiangxi/33775/2013(H10N3) A/duck/Jiangxi/33641/2013(H10N3) A/duck/Jiangxi/33765/2013(H10N3) A/duck/Jiangxi/33753/2013(H10N3) A/duck/Jiangxi/33695/2013(H10N3) A/duck/Jiangxi/33777/2013(H10N3) A/duck/Jiangxi/33769/2013(H10N3) A/duck/Jiangxi/33778/2013(H10N3) A/Mallard/Sweden/816/2014(H1N1) A/barnacle goose/Netherlands/2/2014(H3N6)
A/Waterfowl/Korea/S017/2016(H7N7) A/duck/Mongolia/154/2015(H1N2) A/duck/Mongolia/326/2015(H10N3) A/duck/Mongolia/245/2015(H10N3) A/mallard duck/Netherlands/2/2011(H10N7) A/mallard duck/Netherlands/47/2010(H10N7)
A/mallard/Ontario/15915/2005(H6N5) A/snow goose/Delaware/07OS201/2007(H1N8) A/mallard/Maryland/05OS693/2005(H3N2)
A/shorebird/Delaware Bay/443/2005(mixed) A/environment/Delaware/234/2005(H11N8) A/shorebird/Delaware Bay/146/2005(H11N6) A/ruddy turnstone/New Jersey/1321410/2005(H11N8) A/ruddy turnstone/New Jersey/669/2005(H11N8) 76
91 87 99 78 99
45 98
98 99 96
65 87
90 98
0.020
PB2 (A)
Eurasian
North American
A B
C D
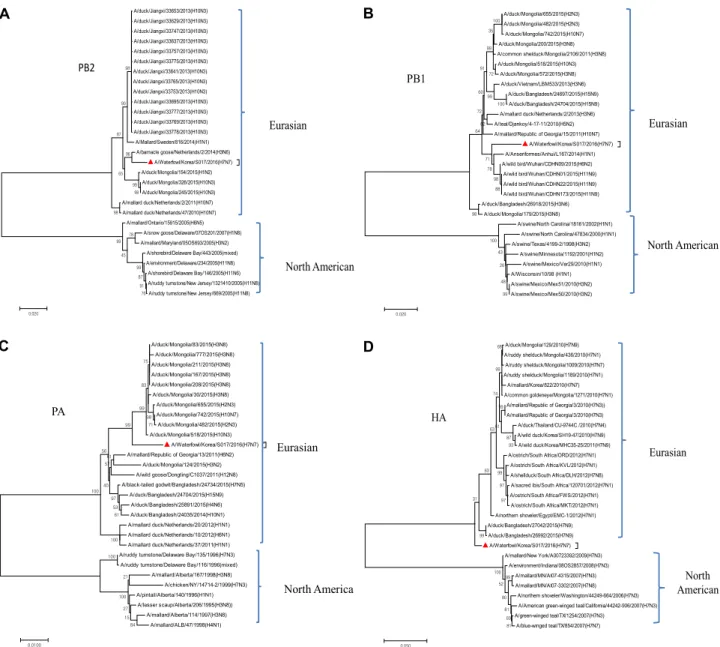
Fig. 1. Phylogenetic analysis of PB2, PB1, PA, HA, NP, NA, M and NS genes of A/Waterfowl /Korea/S017/2016(H7N7). The tree was constructed using Maximum-likelihood method in MEGA7 (www.megasoftware.net) with 1000 bootstrap replicates.
Scale bar shows nucleotide substitutions per site. A) PB2; B) PB1; C) PA; D) HA; E) NP; F) NA; G) M; H) NS. Nucleotide region used in the phylogenetic analysis includes; PB2:1-1341, PB1:1-314, PA: 1-2232, HA: 1-1708; NP: 1-1566, NA: 1-1450, M: 1-1017, NS: 1-890.
(threonine) 215A (alanine), that is known to increase viral pathogenesis in mice, was found [6]. Our isolate had serine (S) at position 31 of M2, suggesting its sensitivity to ion-channel inhibitors, such as amantadine and rimantadine [14]. NS1 had serine (S) at position 42, which is involved in the increased virulence in mice [9]. Other genes involved in causing the pathogenicity and transmission in mammals were not found in our isolate.
The amino acid analysis of our isolate showed that this virus prefers binding to avian species than humans. It sug-
gests that our isolate, H7N7 may not easily infect humans.
However, since avian influenza virus continues to mutate, we need to survey domestic pigs and poultry where avian influenza viruses can mutate by the multiple replications in these hosts. Our analysis also showed that NA protein of our isolate does not have the mutation which is responsible for the resistance against the drugs of influenza virus. It sug- gests that the infected humans, if occurred, can be treated with NA inhibitors, oseltamivir and zanamivir.
A/duck/Mongolia/245/2015(H10N3) A/duck/Mongolia/154/2015(H1N2 A/duck/Mongolia/326/2015(H10N3) A/wild bird/Wuhan/CDHN01/2015(H11N9) A/wild bird/Wuhan/CDHN22/2015(H11N9) A/wild bird/Wuhan/CDHN173/2015(H11N9) A/duck/Mongolia/996/2015(H3N8)
A/duck/Mongolia/101/2015(H4N6) A/duck/Mongolia/127/2015(H4N6) A/duck/Mongolia/543/2015(H4N6)
A/Waterfowl/Korea/S017/2016(H7N7) A/mallard duck/Netherlands/2/2015(H7N7) A/mallard duck/Netherlands/3/2015(H7N7) A/mallard duck/Netherlands/8/2014(H7N5) A/duck/Mongolia/499/2015(H10N7) A/duck/Mongolia/709/2015(H10N7) A/duck/Mongolia/173/2015(H3N8) A/duck/Mongolia/179/2015(H3N8)
A/wild bird/Wuhan/CDHN09/2015(H6N2) A/wild bird/Wuhan/CDHN15/2015(H6N2) A/duck/Wuhan/WHYF05/2014(H9N2)
A/mallard/Alberta/256/2006(mixed) A/mallard/Alberta/106/2007(mixed)
A/mallard/Alberta/246/2006(H4N6) A/mallard/Alberta/115/2007(mixed) A/American wigeon/California/HKWF295/2007(H6N5)
A/mallard/Alberta/240/2005(H1N1) A/mute swan/Rhode Island/A00325136/2008(H7N3)
A/mute swan/Rhode Island/A00325114/2008(H7N3) 63
48 94 100 99
98 98 98
68
88 95
90
82 77
31
51
65
84
0.0050
M (G)
Eurasian
North America
A/duck/Korea/J31/2009(H7N2) A/chicken/Korea/C47/2009(H9N2))
A/spot-billed duck/Xianghai/427/2011(H5N2) A/wild duck/Korea/SH5-26/2008(H4N6)
A/duck/Shanghai/408-1/2009(H4N6) A/duck/Shanghai/420-2/2009(H4N6)
A/duck/Fujian/1761/2010(H10N3) A/duck/Shanghai/MH-2/2009(H4N6) A/mallard/Korea/GH170/2007(H7N7)
A/mallard duck/Netherlands/78/2006(H6N8) A/mallard duck/Netherlands/34/2006(H3N8) A/Waterfowl/Korea/S017/2016(H7N7) A/duck/Hokkaido/W240/2014(H5N3)
A/duck/Japan/11OG1038/2011(H5N2) A/duck/Japan/11OG1083/2011(H5N2) A/chicken/Korea/C47/2009(H9N2) A/duck/Japan/11OG1084/2011(H5N2) A/duck/Guangxi/113/2012(H6N8)
A/duck/Guangxi/GXd-1/2011(H1N2) A/duck/Guangxi/GXd-4/2011(H1N2) A/wild bird/Jiangxi/P419/2016(H6N8)
A/northern shoveler/Washington/44249-783/2006(H7N3) A/mallard/Illinois/3051/2009(H11N3)
A/northern shoveler/Minnesota/Sg-00646/2008(mixed) A/mallard/Minnesota/Sg-00570/2008(H8N4) A/mallard/Alberta/106/2007(mixed) A/green winged teal/Mexico-Sonora/266/2008(H6N1) A/mallard/Washington/44338-195/2007(H12N5) A/northern shoveler/California/HKWF1325/2007(H8N4) 60
58 100
99
94 94
58
39 39 43
34 13
0.0050
NS (H)
Eurasian
North America
A/duck/Mongolia/211/2015(H3N8) A/duck/Mongolia/208/2015(H3N8) A/duck/Mongolia/83/2015(H3N8) A/duck/Mongolia/167/2015(H3N8) A/duck/Mongolia/20/2015(H3N8)
A/chicken/Wuhan/WHJF/2014(H5N2) A/duck/Ganzhou/GZ5/2015(H4N6)
A/Waterfowl/Korea/S017/2016(H7N7) A/wild bird/Jiangxi/P419/2016(H6N8) A/duck/Vietnam/LBM300/2012(H10N2)
A/duck/Quang Ninh/14/2013(H3N6) A/duck/Bangladesh/24704/2015(H15N9)
A/goose/Taiwan/01003/2015(H5N8) A/goose/Taiwan/01019/2015(H5N8) A/mallard/Xuyi/14/2015(H3N8) A/duck/Mongolia/129/2015(H3N3) A/duck/Mongolia/122/2015(H3N3) A/wild goose/Dongting/C1037/2011(H12N8)
A/duck/Mongolia/200/2015(H3N8) A/wild waterfowl/Dongting/C2383/2012(H1N2) A/mallard duck/Georgia/3/2012(H7N3)
A/mallard/Alberta/134/2007(H3N8) A/mallard/Alberta/115/2007(mixed) A/mallard/Alberta/156/2007(mixed)
A/mallard/Alberta/246/2006(H4N6) A/green winged teal/Iowa/463990-3/2006(H5N2)
A/mallard/Alberta/121/2008(H4N6) A/mallard/Alberta/106/2007(mixed)
A/mallard/Alberta/145/2007(H4N6) 79
98
86 100
30 100
100
80 91
24
29 27 51 36 41 65 87
0.0100
NP (E)
Eurasian
North America
A/mallard duck/Netherlands/2/2014(H10N7) A/mallard duck/Netherlands/1/2014(H10N7) A/mallard duck/Netherlands/13/2013(H7N7) A/harbour seal/Germany/1/2014(H10N7)
A/mallard duck/Netherlands/3/2015(H7N7) A/mallard duck/Netherlands/2/2015(H7N7) A/mallard duck/Netherlands/31/2013(H10N7) A/mallard duck/Netherlands/33/2014(H10N7) A/mallard duck/Netherlands/5/2012(H10N7)
A/mallard/Republic of Georgia/14/2011(H10N7) A/domestic duck/Republic of Georgia/1/2010(H10N7) A/domestic duck/Republic of Georgia/2/2010(H10N7) A/mallard/Republic of Georgia/1/2010(H10N7)
A/mallard duck/Netherlands/47/2010(H10N7) A/mallard duck/Netherlands/2/2011(H10N7) A/chicken/Jiangxi/10784/2014(H7N7)
A/duck/Mongolia/626/2015(H10N7) A/duck/Mongolia/709/2015(H10N7) A/duck/Mongolia/499/2015(H10N7) A/duck/Mongolia/742/2015(H10N7)
A/Waterfowl/Korea/S017/2016(H7N7) A/duck/Manitoba/1953(H10N7)
A/mallard/Alberta/553/1986(H10N7) A/mallard duck/ALB/302/1977(H10N7) A/pintail duck/ALB/303/1977(H10N7)
A/scaup/Saskatchewan/9/1981(H10N7) A/seal/Massachusetts/1/1980(H7N7)) A/seal/Massachusetts/1/1980(H7N7) A/seal/Mass/1-SC35M/1980(H7N7) 100
100 68
46 99 37
99 100 100
100 100
42 72 96 8074 40 68
99
99
42 52
0.050
NA (F)
Eurasian
North America
E F
G H
Fig. 1. Continued.
Phylogenetic analysis
The phylogenetic analysis of the isolate showed that all of the eight genes, PB2, PB1, PA, HA, NP, NA, M, and NS belonged to the Eurasian lineage of avian influenza viruses (Fig. 1A-H). We found out that all gene segments of our isolate (A/waterfowl/Korea/S017/2016(H7N7)) had 90-99%
nucleotide similarity with A/magpie/Korea/YJD174/2007 (H7N7) reported by Kim et al [11]. The nucleotide sim- ilarities between them were; 93% (PB2 gene), 92% (PB1 gene), 96% (PA gene), 90% (HA gene), 96% (NP gene), 94%
(NA gene), 97% (M gene), and 99% (NS gene). Furthermore, the Phylogenetic analysis of all H7-subtype avian influenza viruses previously isolated in South Korea were belonged
to a Eurasian lineage [11], which correlates with our findings.
However, the reassortment statuses of H7N7 viruses isolated in South Korea were not indicated on the previous study.
None of our isolate belonged to North American lineages, suggesting that migratory birds migrating in Eurasian re- gions carried our isolate. No H7N7 HPAI was isolated from the domestic ducks in Korea.
In conclusion, our study suggests that continuous surveil- lance of wild birds and domestic poultry needs to be carried out to prevent the creation of reassortant avian influenza viruses, which may pose threat to the domestic animals and humans.
Acknowledgment
This work was funded by the Basic Science Research Program through the National Research Foundation of Korea (NRF) from the Ministry of Science and Technology (2015R1A2A2A01003943).
References
1. Belser, A. J., Creager, M. H., Taronna, R. T. and Tumpey, M. T. 2017. Pathogenesis, transmissibility, and tropism of a highly pathogenic avian influenza A (H7N7) virus asso- ciated with human conjunctivitis in Italy, 2013. J. Infect. Dis.
216, 508-S511.
2. Campitelli, L., Di Martino, A., Spagnolo, D., Smith, G. J., Di Trani, L., Facchini, M., De Marco, M. A., Foni, E., Chiapponi, C., Martin, A. M., Chen, H., Guan, Y., Delogu, M. and Donatelli, I. 2008. Molecular analysis of avian H7 influenza viruses circulatingin Eurasia in 1999-2005: detection of mul- tiple reassortant virus genotypes. J. Gen. Virol. 89, 48-59.
3. Cardona, C., Yee, K. and Carpenter, T. 2009. Are live bird markets reservoirs of avian influenza? Poult Sci. 88, 856-859.
4. CDC. 2017. Types of Influenza Viruses. https://www.cdc.
gov/flu/about/viruses/types.htm. Accessed on April 04, 2018.
5. Cui, P., Deng, G., Shi, J., Kong, H., Liu, L., Guan, Y., Suzuki, Y. and Chen, H. 2016. New influenza A (H7N7) viruses de- tected in live poultry markets in China. Virology 499, 165- 169.
6. Fan, S., Deng, G., Song, J., Tian, G., Suo, Y., Jiang, Y., Guan, Y., Bu, Z., Kawaoka, Y. and Chen, H. 2009. Two amino acid residues in the matrix protein M1 contribute to the virulence difference of H5N1 avian influenza viruses in mice. Virology 384, 28-32.
7. Hay, A. J., Wolstenholme, A. J., Skehel, J. J. and Smith, M.
H. 1985. The molecular basis of the specific anti-influenza action of amantadine. EMBO J. 4, 3021-3024.
8. Horimoto, T. and Kawaoka, Y. 2001. Pandemic threat posed by avian influenza A viruses. Clin. Microbiol. Rev. 14, 129- 149.
9. Jiao, P., Tian, G., Li, Y., Deng, G., Jiang, Y., Liu, C., Liu, W., Bu, Z., Kawaoka, Y. and Chen, H. A. 2008. Single-ami- no-acid substitution in the NS1 protein changes the pathoge- nicity of H5N1 avian influenza viruses in mice. J. Virol. 82, 1146-1154.
10. Kawaoka, Y., Chambers, T. M., Sladen, W. L. and Webster,
R. G. 1988. Is the gene pool of influenza viruses in shore- birds and gulls different from that in wild ducks? Virology 163, 247-250.
11. Kim, C. M., Jeong M. O., Hyun-Mi Kang, M. H., Paek, R.
M., Kwona, S. J., Song, S. C., Kwon, K. Y., Lee, G. J., Kwona, H. J. and Lee, J. Y. 2010. Pathogenicity and transmission studies of H7N7 avian influenza virus isolated from feces of magpie origin in chickens and magpie. Vet. Microbiol. 141, 268-274.
12. Kurtz, J., Manvell, R. J. and Banks, J. 1996. Avian influenza virus isolated from a woman with conjunctivitis. Lancet 348, 901-902.
13. Liu, M., He, S., Walker, D., Zhou, N., Perez, D. R., Mo, B., Li, F., Huang, X., Webster, R. G. and Webby, R. J. 2003.
The influenza virus gene pool in a poultry market in South central china. Virology 305, 267-275.
14. McKimm-Breschkin, J. L., Sahasrabudhe, A., Blick, T. J., McDonald, M., Colman, P. M., Hart, G. J., Bethell, R. C.
and Varghesem, J. N. 1998. Mutations in a conserved resi- due in the influenza virus neuraminidase active site de- creases sensitivity to Neu5Ac2en-derived inhibitors. J. Virol.
72, 2456-2462.
15. Metreveli, G., Siamak, Z., Anneli, E. and Mikael, B. 2009.
Phylogenetic analysis of the hemagglutinin gene of low pathogenic avian influenza virus H7N7 strains in mallards in Northern Europe. Avian Dis. 54, 453-456.
16. Monne, I., Fusaro, A., Nelson, M. I., Bonfanti, L., Mulatti, P., Hughes, J., Murcia, P. R., Schivo, A., Valastro, V., Moreno, A., Holmes, E. C. and Cattoli, G. 2014. Emergence of a highly pathogenic avian influenza virus from a low-pathogenic progenitor. J Virol. 88, 4375-4388.
17. Nidom, C. A., Takano, R., Yamada, S., Sakai-Tagawa, Y., Daulay, S., Aswadi, D., Suzuki, T., Suzuki, Y., Shinya, K., Iwatsuki-Horimoto, K., Muramoto, Y. and Kawaoka, Y.
2010. Influenza A (H5N1) viruses from pigs, Indonesia.
Emerg. Infect. Dis. 16, 1515-1523.
18. Smietanka, K., Pikula, A., Minta, Z. and Meissner, W. 2011.
Evidence of persistence and multiple genetic modifications of H7N7 low-pathogenic avian influenza virus in wild mal- lards in Poland provided by phylogenetic studies. Avian Pathol. 40, 131-138.
19. Webster, R. G., Bean, W. J., Gorman, O. T., Chambers, T.
M. and Kawaoka, Y. 1992. Evolution and ecology of influen- za A viruses. Microbiol. Rev. 56, 152-179.
20. Wright, P. F. and Webster, R. G. 2001. Orthomyxoviruses, pp. 1533-1579. In: Knipe, D. M., Howley, P. M. (Eds.), Fields Virology. Lippincott, Williams and Wilkins, Philadelphia, USA.
초록:2016년 한국 야생조류에서 분리한 H7N7 조류인플루엔자 바이러스 유전자 분석
베리훈1,2․서상희1,2*
(1충남대학교 수의과대학, 2독감바이러스 연구소)
A형 인플루엔자바이러스는 야생조류에 존재하며, 사람, 돼지, 가금 및 다른 포유류등 다양한 숙주를 감염한다.
본 연구에서는, 2016년 한국 서쪽의 철새도래지에서 채취한 철새 분변에서 재조합된 새로운 H7N7 조류인플루엔 자를 분리하였으며 이 바이러스의 8개 유전자를 분석하였다. 분리된 A/waterfowl/Korea/S017/2016(H7N7) 바이 러스의 유전자 분석 결과 이 바이러스는 야생조류 및 가금 오리에서 유래한 조류인플루엔자로 구성된 재조합된 유전자를 가지고 있었다. 계통 분석결과 이 바이러스는 유럽과 아시아계에 속하였다. 조류인플루엔자 바이러스가 계속 진화를 하고 H7 형 조류인플루엔자는 고 병원성 조류인플루엔자로 변하여 사람과 동물에게 커다란 위협이 될 수 있기에 계속 된 역학조사가 필요하다.