홍조류로부터 신규 한천분해미생물 Alteromonas macleodii subsp.
GNUM08120의 분리 및 동정
지원재1, 임주현1, 박다연1, 김무찬2, 김창준3, 장용근4, 홍순광1*
1명지대학교생명과학정보학부
2경상대학교해양환경공학과및해양산업연구소
3경상대학교생명화학공학과및공학연구원
4한국과학기술원생명화학공학과
Received : August 1, 2012 / Revised : September 29, 2012 / Accepted : October 2, 2012
서 론
한천은홍조류세포벽의구성성분으로두종류의
polysac- charide
인agarose
와agaropectin
의복합체이다. Agarose
는3-O-linked
β-
D-galactopyranose
와4-O-linked
α-
L-galac- topyranose unit
가반복적으로연결되어있는구조를갖는 데, agaropectin
은sulfoxy, methoxy, pyruvate
잔기 등에 의해서수식된3,6-anhydro-
α-
L-galactose
가존재한다는점 에서agarose
와구별된다[21]. Agarase
는이러한agarose
를 분해하는효소로,
그작용 기작에따라서 α-agarase
와 β-
agarase
로나뉜다.
α-agarase
는α-1,3
결합을분해하여환 원말단에3,6-anhydro-
α-
L-galactose
를 갖는agarooligo- saccharide
를생산하고,
β-agarase
는β-1,4
결합을분해하여 환원말단에D-galactose
를갖는neoagarooligosaccharide
를 생산한다.
한천으로부터생산되는oligosaccharide
는다양 한생물학적기능이규명되어식품,
화장품,
의약분야 등 의많은산업 분야에적용될 수있을것으로 기대되고있 다[15].
해조류로부터의바이오에너지생산은
starch
또는ligno- cellulose
기반의바이오매스를재료로한경우보다많은장 점이있다.
우선,
해조류는 해양에서태양에너지만으로쉽 게자라기때문에경작을위한비용이절감되고,
경작지가 필요치않으며,
화학비료가사용되지않기때문에자연친화 적이다.
또한해조류유래의다당류에는리그닌이존재하지Isolation and Characterization of a Novel Agar Degrading Bacterium, Alteromonas macleodii subsp. GNUM08120, from Red Macroalgae. Chi, Won-Jae
1, Ju-Hyeon Lim
1, Da Yeon Park
1, Mu-Chan Kim
2, Chang-Joon Kim
3, Yong-Keun Chang
4, and Soon-Kwang Hong
1*.
1Department of Biological Science, Myongji University, Yongin, Gyeonggido 449-728, Korea,
2Department of Marine Environmental Engineering and
3Department of Chemical and Biological Engineering, Gyeong- sang National University, Gyeongnam 650-160, Korea,
4Department of Chemical and Biomolecular Engineering, Korea Advanced Institute of Science and Technology, Daejeon 305-701, Korea
An agar-hydrolyzing marine bacterium, strain GNUM08120, was isolated from Sargassum fulvellum collected from Yeongil bay of East Sea of Korea. The isolate was Gram-negative, aerobic, motile with single polar flagellum, and grew at 1-10% NaCl, pH 5.0-8.0, and 15-37
oC. G+C content and the predominant respiratory quinone were 46.13 mol% and Q-8, respectively. The major cellular fatty acids were Summed feature 3 (24.5%), C
16:0(21.7%), and C
18:1ω7c (12.5%). Based on 16S rRNA gene sequence similarity and DNA-DNA hybridization analyses, strain GNUM08120 was identified as a novel subspecies of Altero- monas macleodii, designated Alteromonas macleodii subsp. GNUM08120. Production of agarase by strain GNUM08120 was likely repressed by the effect of carbon catabolite repression caused by glucose. The crude agarase prepared from 12-h culture broth of strain GNUM08120 exhibited an optimum pH and temperature for agarase activity at 7.0 and 40
oC, respectively. The crude enzyme produced (neo)agarobiose, (neo)agarotetraose, and (neo)agarohexaose as the hydrolyzed product of agarose.
Keywords: Agarase, Alteromonas macleodii, red macroalgae, phylogenetic tree
*Corresponding author
Tel: +82-31-330-6198, Fax: +82-31-335-8249
E-mail: [email protected]
않기때문에높은수율의에너지전환율을기대할수있다
.
전통적으로한천분해는화학적분해법을통하여수행되었는 데,
최근에자연친화적인방법인효소적분해법이주목받고 있다.
이러한효소적분해법은원하는길이의oligosaccharide
를특이적으로생산할수있다는장점이있어많은연구가 진행중이다[29, 7].
이에본연구에서는,
해양으로부터해조류
agar
의분해능력이있는새로운미생물을동정하고이로부터신규의
agarase
효소를분리하여그특성을규명함 으로써산업적으로유용한효소를탐색하고자하였다.
재료 및 방법
미생물 배양조건 및 시약
홍조류로부터미생물을분리하고배양하기위해서사용된 배지는
Artificial Sea Water (ASW)
배지(MgSO
412.3 g, KCl 0.74 g, (NH
4)
2HPO
40.13 g, NaCl 17.5 g, CaCl
20.14 g, Tris base 6.2 g per 1 L, pH 7.2) [6]
에Bacto-Peptone 10 g
과Yeast extract 3 g
을첨가하여ASW-YP
로명명하여사용 하였다.
평판배지는한천을1.5%
가되도록첨가하였고,
액체 배지는한천을0.3%
가되도록첨가하여사용하였다.
미생물 배양은28
oC
에서수행하였으며,
액체배지의경우에는200 rpm
으로진탕배양하였다.
배지및효소활성측정을위해사용 된시약들은모두Sigma
사로부터구입하였고16S rRNA
유 전자의PCR
증폭을위해사용된효소등은Takara (Japan)
로 부터 구입하였다. PCR
에 사용된primer
는Genotech
사(Daejeon, Korea)
에의뢰하여합성하였으며, genomic DNA
와agarose gel
로부터DNA fragment
의추출은각각DyneBio
사(Seongnam, Korea)
의Genomic DNA extraction kit
와Gene extraction kit
를사용하여수행하였다. API 20NE Kit (Biomérieux, France)
와API ZYM Kit (Biomérieux, France)
를사용하여
GNUM08120
균주의대사특성과효소활성을측정하였다
. Cellulase, amylase, caseinase
등의효소활성측정 시,
효소반응기질은AZCL-HE-cellulose (Megazyme, Ireland), starch azure (Sigma, USA), azocasein (Sigma, USA)
을 각각 사용하였다.
Agarase 생산균주의 순수 분리
Agarase
를생산하는미생물을분리하기위해서동해영일만의해안에서식하는홍조류
Sargassum fulvellum
을채취 하였다. S. fulvellum
을멸균해수로2
회씻어준후잘게잘 라1 ml
의 멸균수에 현탁하였다.
현탁액은10
-2~10
-5배로serial dilution
하여0.1 ml
를ASW
평판배지[6]
에도말하여28
oC
에서5
일간 배양하였다. Lugol’s iodine
용액(25 g/L Iodine, 50 g/L Potassium Iodine)
으로염색하여콜로니주변에한천분해환을형성하는각각의콜로니를
agarase
생산주로서선별하였다
.
이후의모든배양에서는박테리아의 빠른성장을위하여ASW-YP
배지를사용하였다.
선별된콜로니는
ASW-YP
평판배지에도말하여같은조건에서배양하였다
.
생성된콜로니를다시새로운배지에계대하는작업 을반복하여순수한균주를확보하였다.
16S rRNA 유전자 염기서열 분석과 계통수(phylogenetic tree) 구축
선별된
Agarase
생산균주를ASW-YP
액체배지에서2
일 간진탕배양후15,000 rpm
에서10
분간원심분리하여균체 만을회수하였다.
회수된균체로부터genomic DNA
를추출하여 이를 주형으로사용하여
PCR
반응을 실시하여16S
rRNA
를코드하는 유전자단편을증폭하였다[19]. PCR
반 응을 위한 프라이머는bacterial universal primer (27F
와1492R)
를사용하였으며,
증폭된DNA
단편은Genotech Inc.
(Daejeon, Korea)
에염기서열분석을의뢰하였다.
확보된16S rRNA
유전자의염기서열은GenBank
에JN578475
로등록 하였으며, National Center for Biotechnology Information (NCBI)
의BlastN program (http://blast.ncbi.nlm.nih.gov/
Blast.cgi)
을사용하여GenBank
의database
의정보로부터 염기서열의 상동성 검사가 수행되었다.
또한EzTaxon database (http://www.eztaxon.org/)
로부터표준균주의16S
rRNA
유전자염기서열을확보하여계통발생적연관성을분석하였다
[9].
확보된 염기서열들간의Multi alignment
는ClustalW program [28]
을사용하였고5
’과3’-end
의gap
은GeneDoc program [11]
으로 편집하였다.
진화적인 거리(evolutionary distance matrix)
는Kimura’s two-parameter evolutionary model [13]
에 의해 계산하였고Neighbor- Joining [26]
방법으로phylogenetic tree
를 작성하였다. Branch
의신뢰도(bootstrap value)
는1,000
회의재구성된자 료로부터새롭게tree
를작성하여계산하였다[11].
DNA-DNA hybridization
DNA-DNA hybridization
분석을 위해서, GNUM08120
균주와계통발생적유연관계가높은표준균주인Alteromonas
macleodii DSM6062
T(Y18228) [4], Alteromonas marina
SW-47
T(AF529060) [32], Alteromonas simiduii BCRC17572
T(DQ836766) [8]
등의균주를LB
평판배지에서배양하여균 체로부터DNA
를추출하여준비하였다. Negative standard
균주로는Staphylococcus simulans KCCM41686
T가사용 되었다.
탐침용DNA
준비와hybridization
반응은DIG
High Prime DNA Labeling and Detection Starter Kit II
(Roche Applied Science, Germany)
를사용하여주어진방 법에따라수행하였다. Hybridization signal
은Quantity One
Program (Bio-rad, USA)
으로측정하였고, GNUM08120
균주의
signal
을100%
로하여표준균주들의값을환산하였다.
각 균주들에대한hybridization
값은세번의반복적인실험을통 하여계산하였다.
화학적 계통분류
Chemotaxonomic
특징은GNUM08120
균주를ASW-YP
평판배지에서28
oC, 48
시간배양후균체를회수하여분석 하였다.
주요respiratory quinones
은reverse phase HPLC
로분석하였고[16],
균체의지방산은Miller and Berger [23]
의방법에준하여
Methyl ester
화시켰으며,
전처리된fatty acid methyl esters (FAME) mixture
의 분석은Microbial Identification system (MIDI)
의지침에따라gas chroma- tography
분석법에의해수행되었다[27]. DNA G+C content
는reverse phase HPLC
를이용한Mesbah
등[22]
의방법에 따라서분석하였다.
항생제 내성시험
GNUM08120
균주를ASW-YP
평판배지에골고루도말한 후,
각각의 항생제[thiostrepton (50
µg/
µl), kanamycin (50
µg/
µl), neomycin (30
µg/
µl), ampicillin (50
µg/
µl), apramycin (50
µg/
µl), chloramphenicol (25
µg/
µl), nalidixic acid (25
µg/
µl)]
를30
µl
씩함유하는paper disc
를올려놓은 후28
oC
에서24
시간배양하여paper disc
주변에형성되는 투명한성장억제환의직경을측정하였다.
GNUM08120 균주의 세포 성장 및 agarase 생산 균주의성장및
agarase
생산을관찰하기위해서, ASW- YP
액체배지에0.3% glucose, 0.3% sucrose, 0.3% starch
를 각각첨가하여28
oC
에서진탕배양하면서3
시간간격으로 샘플을 채취하였다.
균체 성장곡선은spectrophotometer (Genesys 8, Spectronic Unicam, England)
를이용하여600 nm
에서흡광도를측정하여결정하였고, agarase
활성은샘 플을원심분리(15.000 rpm, 10 min)
하여균체를제거한상 등액을 이용하여0.2% agarose
를 기질로 하는3,5-dini- trosalicylic acid (DNS)
법[25]
으로540 nm
에서측정하였다.
GNUM08120 균주가 분비 생산하는 agarase의 생화학적 특성GNUM08120
균주를0.3% agar
가포함된ASW-YP
액체 배지에서12
시간배양하여원심분리후균체를제거하고배 양액을농축하여효소액으로사용하였다[10].
효소반응을위 한 최적pH
조건을 탐색하기 위해서,
준비된 효소액을20 mM MOPS
버퍼(pH 6.0-7.0), 20 mM Tris-Cl
버퍼(pH 7.0-9.0), 20 mM glycine-NaOH
버퍼(pH 9.0-10.0)
로40
oC
에서30
분간agarose
를기질로하는효소반응을실시 한후DNS
법으로활성을측정하였다.
최적온도조건을탐색하기위해서
,
효소액을20 mM Tris-Cl
버퍼를사용하여 다양한온도조건에서30
분간반응을유도하여활성을측정 하였다. GNUM08120
균주가세포외부로분비하는agarase
에의한agarose
가수분해산물을분석하기위해서, Temuujin
등[29]
에의해서 서술된방법에 따라서silica gel 60 plate (Merck, USA)
를이용한Thin layer chromatography (TLC)
를 수행하였다.
결과 및 고찰
GNUM08120 균주의 형태학적, 생화학적 특성 분석 영일만해안에서식하는홍조류
Sargassum fulvellum
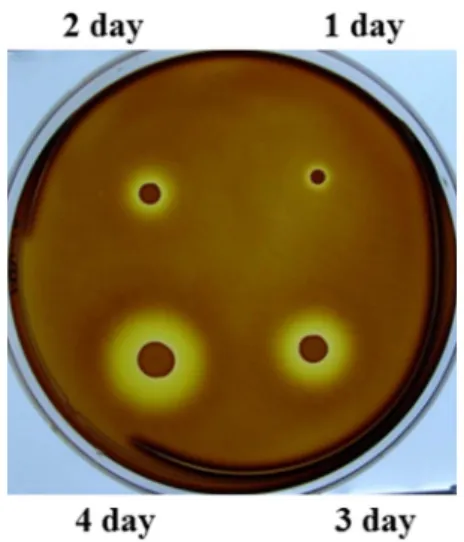
로 부터분리된균주들중에서콜로니주변에한천분해환을형 성하는균주만을일차선별하였고,
그중에서한천분해능이 뛰어난하나의균주를선별하여(Fig. 1) GNUM08120
으로명 명하였으며Korean Collection for Type Culture (KCTC)
에기탁하였다(
기탁번호; KCTC23885).
GNUM08120
균주는그람염색법을통하여그람음성균으로판명되었고
,
전자현미경관찰결과포자를형성하지않 는박테리아로확인되었다. ASW-YP
평판배지에서28
oC
로24
시간배양한콜로니는원형으로직경이약2-3 mm
정도 이고표면은부드럽고크림색으로관찰되었다.
또한ASW- YP
평판배지에서28
oC
로24
시간배양한세포를negative staining
을 통한Transmission Electron Microscopy (TEM)
관찰을실시한결과, GNUM08120
균체는주길이가약2.0- 2.5
µm
이고,
폭이약0.5-0.6
µm
이며, single polar flagella
를갖고있어운동성이있을것으로예상되었다(Fig. 2).
Fig. 1. Zymogram plate assay results of the GNUM08120 as a function of cultivation time.
The strain was cultured on ASW-YP agar plate and Lugol’s iodine
solution was overlayed to detect reducing sugars, degraded prod-
uct from agar by agarase.
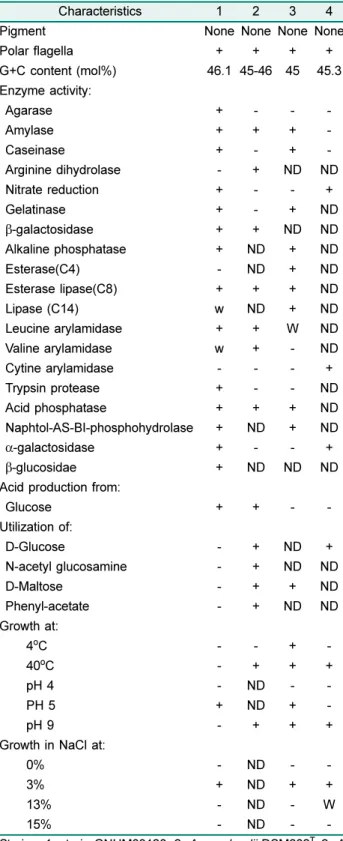
GNUM08120
균주의형태학적,
생화학적,
생리학적특성 을Table 1
에요약하였다. GNUM08120
균주는15-37
oC
에 서는성장하지만4
oC
이하및40
oC
이상에서는5
일간관찰 하여도성장하지못하였다.
또한GNUM08120
균주는pH 5.0-8.0
사이의pH
범위에서는자랄 수있고,
특히pH 7.0 -8.0
의조건에서가장빠른성장을보였다.
반면에pH 4.0
이하와
pH 9.0
이상의조건에서는장시간배양해도성장이관찰되지 않았다
. GNUM08120
균주는0%
및13%
이상의NaCl
을포함하는ASW-YP
평판배지에서는5
일이상배양 해도균의성장이관찰되지않은반면, 1-5%
의NaCl
을포함 하는배지에서는배양초기(1
일째)
에성장및agarase
활성 이관찰되었고, 10%
의NaCl
을포함하는배지에서는장시간(3
일이상)
배양할때성장및agarase
활성이관찰되었다. GNUM08120
균주는agarase, amylase, caseinase, oxidase, nitrate reductase, gelatinase,
β-galactosidase, alkaline phosphatase, esterase lipase (C8), leucine arylamidase, trypsin protease, acid phosphatase, naphtol-AS-BI-phos- phohydrolase,
α-galactosidase,
β-glucosidase
등의효소활 성이양성반응으로관찰되었고, lipase (C14)
와valine aryl- amidase
등은 약한 양성반응을 나타냈으며, arginine dihydrolase
와urease
등의효소활성은음성반응으로관찰 되었다. Glucose acidification
은 양성, indole
생산은 음성 반응으로관찰되었다.
D-glucose,
L-arabinose,
D-mannose,
D-mannitol, N-acetyl-glucosamine,
D-maltose, potassium gluconate, capric acid, adipic acid, malic acid, trisodium citrate, phenylacetic acid
등은단일탄소원으로사용되지 않는것으로관찰되었다. GNUM08120
균주에는Quinone-
Table 1. Phenotypic and chemotaxonomic characteristics of GNUM08120 and related Alteromonas species.
Characteristics 1 2 3 4
Pigment None None None None
Polar flagella + + + +
G+C content (mol%) 46.1 45-46 45 45.3 Enzyme activity:
Agarase + - - -
Amylase + + + -
Caseinase + - + -
Arginine dihydrolase - + ND ND
Nitrate reduction + - - +
Gelatinase + - + ND
β-galactosidase + + ND ND
Alkaline phosphatase + ND + ND
Esterase(C4) - ND + ND
Esterase lipase(C8) + + + ND
Lipase (C14) w ND + ND
Leucine arylamidase + + W ND
Valine arylamidase w + - ND
Cytine arylamidase - - - +
Trypsin protease + - - ND
Acid phosphatase + + + ND
Naphtol-AS-BI-phosphohydrolase + ND + ND
α-galactosidase + - - +
β-glucosidae + ND ND ND
Acid production from:
Glucose + + - -
Utilization of:
D-Glucose - + ND +
N-acetyl glucosamine - + ND ND
D-Maltose - + + ND
Phenyl-acetate - + ND ND
Growth at:
4
oC - - + -
40
oC - + + +
pH 4 - ND - -
PH 5 + ND + -
pH 9 - + + +
Growth in NaCl at:
0% - ND - -
3% + ND + +
13% - ND - W
15% - ND - -
Strains: 1, strain GNUM08120; 2, A. macleodii DSM602
T; 2, A.
marina SW47
T; 3, A. simiduii BCRC17572
T. All strains were oxi- dase positive. Symbols: +, Positive; -, negative; w, weak posi- tive; ND, no data available.
Fig. 2. Transmission electron micrograph of GNUM08120.
The bacterial cells were removed from 2 days culture on ASW-YP
plate and examined by transmission electron microscopy after neg-
ative staining with 1% phosphotungstic acid. The polar flagellum is
indicated by arrow. Bar, 0.5 µm.
8
이주요respiratory quinone
으로존재하였다. GNUM08120
균주의게놈내G+C content
는46.13 mol%
였다.
GNUM08120
균주의주요cellular fatty acid (>10%)
로서Summed feature 3 (24.5%), C
16:0(21.7%), C
18:1ω7c (12.5%)
등이측정되었다
.
그외의cellular fatty acid
는Table 2
에나 타내었다. GNUM08120
균주는thiostrepton (25
µg)
에는 감수성을보인반면, kanamycin (50
µg), neomycin (30
µg), ampicillin (50
µg), apramycin (50
µg), chloramphenicol (25
µg), nalidixic acid (25
µg)
등에는내성을가지고있는 것으로관찰되었다. GNUM08120
균주를ASW-YP
평판배 지에서배양후그람음성대표균주로서Escherichia coli
와 그람양성대표균주로서Bacillus subtilis
를overlay
하여항 균활성을측정한결과,
두지시균주모두에서어떠한성장 의저해도관찰되지않았다.
GNUM08120 균주의 계통수(phylogenetic tree) 분석
16S rRNA
유전자염기서열을토대로EzTaxon database [9]
에등록되어있는표준균주들의16S rRNA
유전자염기 서열들과의상동성조사결과, GNUM08120
균주는Alter- omonas macleodii DSM6062
T[4]
와 가장 높은98.25%
의 상동성을 보였고, Alteromonas marina SW-47
T[32]
와도98.24%
의높은상동성을보였다.
또한Alteromonas litorea TF-22
T[33], Alteromonas addita R10SW13
T[12], Altero- monas stellipolaris LMG21861
T[30], Alteromonas hispanica F-32
T[20]
과는 각 각97.82%, 97.38%, 97.11%, 97.05%
의 상동성을보였다(Table 3). PHYLIP suit program[11]
을사 용한phylogenetic tree
를제작한결과, GNUM08120
균주 는A. macleodii DSM6062
T, A. marina SW-47
T, A. simiduii BCRC17572
T등과계통발생적연관성을보이고있다(Fig.
3A).
비록Neighbour-Joining analysis
법에의한tree
상에 서신뢰도가50%
미만이지만,
이는Maximum-Likehood
프로그램에의해서그려진
tree
에서도비슷하게나타나고있다
.
계통발생적연관성이가장높은세균주와GNUM08120
균주의형태적,
생리적특성에관한비교는Table 1
과2
에 나타내었다.
Table 3. 16S rRNA gene nucleotide sequnce similarity between strain GNUM08120 (1,453 bases) and its homologous.
Bacterial species GenBank no. Similarity (%) Gene size (bases)
Alteromonas macleodii DSM6062
TY18228 98.254 1492
Alteromonas marina SW-47
TAF529060 98.241 1489
Alteromonas litorea TF-22
TAY428573 97.826 1456
Alteromonas addita R10SW13
TAY682202 97.387 1466
Alteromonas stellipolaris LMG21861
TAJ295715 97.111 1482
Alteromonas hispanica F-32
TAY926460 97.055 1479
Alteromonas genovensis LMG24078
TAM885866 96.901 1501
Alteromonas simiduii BCRC17572
TDQ836766 96.756 1391
Alteromonas tagae BCRC17571
TDQ836765 96.634 1479
Alteromonas halophita JSM073008
TEU583725 95.167 1445
Aestuariibacter halophilus JC2043
TAY207503 95.000 1434
Table 2. Cellular fatty acid profiles of strain GNUM08120 and related Alteromonas species.
Fatty acid 1 2 3 4
Straight-chain fatty acid:
C
12:01.9 2.5 2.9 2.9
C
14:02.5 2.5 2.6 5.7
C
15:01.7 2.5 2.8 -
C
16:021.7 23.8 21.2 22.7
C
16:0N alcohol 7.28 6.6 3.2 tr
C
17:02.14 2.6 3.2 2.0
Hydroxy fatty acid:
C
10:03-OH 1.1 1.5 1.3 tr
Unsaturated fatty acid:
C
15:1ω8c 1.1 1.7 1.1 1.5
C
16:1ω7c alcohol 4.0 4.3 1.9 -
C
17:1ω8c 3.7 4.3 5.6 4.5
C
18:1ω7c 12.5 9.9 11.8 11.3
Methyl fatty acid:
10-Methyl C
17:04.6 2.9 1.5 tr
Summed features:
2 (C
14:03-OH/iso-C
16:1I) 2.35 3.3 3.6 - 3 (C
16:1ω7c/iso-C
15:02-OH) 24.5 24.6 28.5 33.7 Strain: 1, strain GNUM08120; 2, A. macleodii DSM602
T; 2, A.
marina SW47
T; 3, A. simiduii BCRC17572
T. Only fatty acids that
present more than 1% of the total fatty acids are listed. Summed
features: groups of two or three fatty acids that could not be sep-
arated by GLC with the MIDI system. Symbols: tr, trace amount
detected (<1%); -, not detected/not reported.
DNA-DNA hybridization 분석
GNUM08120
균주와16S rRNA
유전자염기서열및계통 발생적연관성이높은균주들과의DNA
연관성조사결과, A.
macleodii DSM6062
T와80.06%
의hybridization
값을보였 다(Fig. 3B).
일반적으로DNA-DNA hybridization
값이70- 90%
일때새로운아종(subspecies)
로간주되며,
그값이90%
이상일때는같은종으로간주되고
, 70%
이하일때는다른종으로 간주된다
[30].
이 기준으로부터80.06%
라는DNA hybridization
값은GNUM08120
균주가A. macleodii DSM6062
T와같은종(species)
으로분류될뿐만아니라아종(subspecies)
으로분류되는값에해당된다. A. marina SW- 47
T 및A. simiduii BCRC17572
T에 대해서는DNA-DNA hybridization
값이 각각74.18%
와35.29%
로 측정되었다.
이값들은다른종으로구분할수있는70%
의cut-off
값과유사하거나매우낮은값으로이들두균주들은
GNUM08120
균주와 다른 종으로 판단되었다
.
본연구에서수행된16S
rRNA
유전자 염기서열분석과 계통 수 및DNA-DNA
hybridization
분석의결과들로부터GNUM08120
균주는A.
macleodii DSM6062
T의아종으로분류가가능하다.
따라서 본논문에서는GNUM08120
균주를Alteromonas macleodii
subsp. GNUM08120
으로명명하였다.
세포성장 및 agarase 생산 특성GNUM08120
균주의성장은탄소원으로한천뿐만아니라
glucose
또는starch
가제공되었을때성장이증가된반 면, sucrose
가제공되었을때는별다른성장의증가효과가 관찰되지않았다(Fig. 4A). GNUM08120
균주가세포외부로생산하는
agarase
는한천만제공된배지에서가장높은활성이관찰되었는데
,
균주성장곡선의stationary phase
에 진입하는시기에생산되어mid-stationary phase
부터급격 히생산량이줄어들었다(Fig. 4B).
또한한천이외의배지 내다른탄소원이제공된배지에서는agarase
활성이약하 게관찰되었는데,
이는 배지내glucose (monosaccharide), sucrose (disaccharide), starch (polysaccharide)
등에의해 서carbon catabolite repression (CCR)
을받는것으로사료된 다.
일반적으로CCR
은glucose
등과같은탄소원의존재시 단백질생합성이억제되는현상을일컫는다. Streptomyces
속의균주들의경우에는효소,
항생제,
생리활성물질등의생 산이CCR
에 의해서 영향을 받는 것으로 알려져 있는데, Streptomyces coelicolor
와Streptomyces griseus
에서는Fig. 3. Phygenetic analysis of strain GNUM08120.
(A) Phylogenetic tree of strain GNUM08120 constructed from the nucleotide sequence of
16S rRNA gene using the neighbour-joining method. The scale bar indicates a genetic dis-
tance of 0.01. The number shown next to each node indicates the percentage bootstrap
value of 1,000 replicates. Accession nos. of the 16S rRNA sequences are given in paren-
theses. (B) DNA-DNA hybridization. (1) strain GNUM08120; (2) A. macleodii DSM6062
T;
(3) A. marina SW-47
T; (4) A. simiduii BCRC17572
T; (5) S. simulans KCCM41686 as the
negative standard.
glucose kinase
에의해서매개되는것으로 보고되어있다[24].
특히, S. coelicolor
가세포외부로생산하는β-agarase (DagA)
는glucose
에의해서생합성이억제된다고보고되어 있다[1].
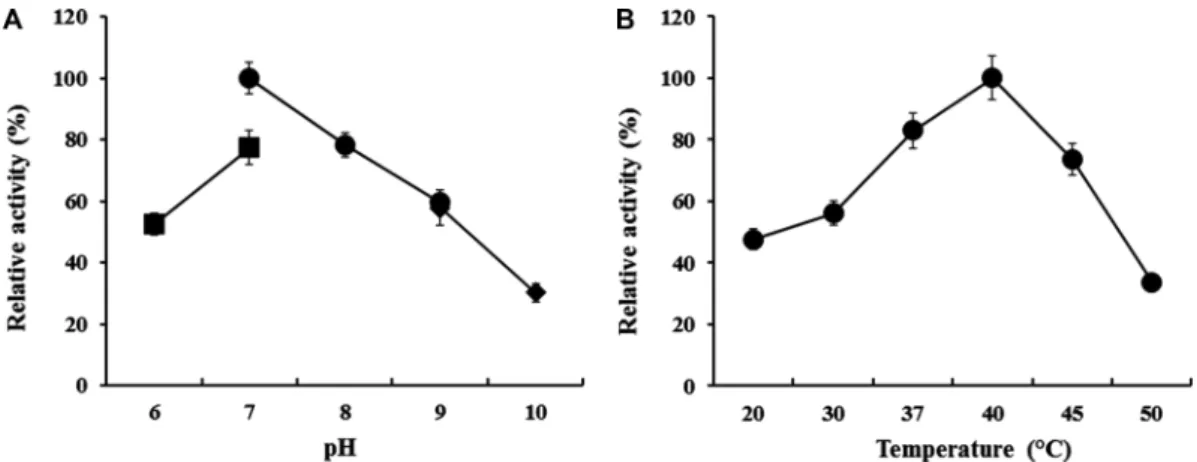
GNUM08120 균주가 생산하는 agarase의 효소적 특성
GNUM08120
균주의12
시간배양효소액을이용한최적pH
및온도조건의탐색결과, pH 7.0
과40
oC
에서최대활성이 관찰되었다(Fig. 5A, 5B).
이러한최적조건은Alteromonas
속의다른균주들로부터보고된
agarase
의특성과일치하였다[14, 18, 31].
그리고GNUM08120
균주의 효소액을 이용한agarose
분해산물의TLC
분석을통해서, (neo)agarobiose, (neo)agarotetraoase, (neo)agarohexaose
등이생산되는것 을알수있었다(Fig. 6).
현재까지Alteromonas
속의균주로 부터총4
개의agarase
가정제되어그특성이보고되었다. Alteromonas GJ1B
로부터agarotetraose
를생산하는360 kDa
의α-agarase
가보고된바있다.
또한, Alteromonas C-1
로부터neoagarotetraose
를 생산하는82 kDa
의 β-agarase [18],
Fig. 5. Enzymatic properties of agarase produced by strain GNUM08120.
(A) Effect of pH on agarase activity. The agarase assay was carried out at 40
oC at different pH conditions ranging from pH 6.0 to pH 10.0. The values obtained at 20 mM Tris-Cl, pH 7.0, were taken to be 100%.
■, 20 mM MOPS buffer;
●, 20 mM Tris-Cl buffer;
◆
, 20 mM Glycine-NaOH buffer. All data shown are mean values from at least three replicate experiments. (B) Effect of temperature on agarase activity. The agarase assay was carried out at pH 7.0 at different temperatures ranging from 20
oC to 50
oC. The values obtained at 40
oC were taken to be 100%. All data shown are mean values from at least three replicate experiments (p < 0.05).
Fig. 4. Effect of carbon source on cell growth (A) and agarase production (B).
(A) Comparison of cell growth in ASW-YP containing various carbon sources. Cell growth was measured by spectrophotometer at
600 nm. (B) Comparison of agarase activity produced in various media. The agarolytic activity was expressed in absorbance at 540 nm
per unit culture volume (ml). ASW-YP containing 0.1% agar as sole carbon source was used as the control. All data are the average
of three parallel replicates (p<0.05). -
●-, 0.3% agar only; -
■-, 0.3% agar + 0.3% glucose; -
▲-, 0.3% agar + 0.3% sucrose; -
◆-,
0.3% agar + 0.3% starch.
Alteromonas E-1
으로부터neoagarobiose
를생산하는82 kDa
의 β-agarase [14], Alteromonas SY37-12
로부터neoagarotetraose
와neoagarohexaose
를생산하는39.5 kDa
의β-agarase [31]
등 이보고되었다. (Neo)agarooligosaccharide
는항박테리아활성,
항산화활성,
항암효과,
미백효과등이있어의약산업,
식음 료산업,
화장품산업에응용이가능하다고알려져있다.
또 한다이어트보조제로서효과가뛰어나현재일본에서는식음 료제품으로출시되어판매되고있다.
따라서GNUM08120
이 세포외부로생산하는agarase
를이용하여생체활성이뛰어난(neo)agarobiose
와(neo)agarotetraose
를생산할수있으므로 여러 산업에 응용이 가능할 것으로 기대된다.
이에 앞서GNUM08120
으로부터생산되는agarase
를순수정제하여 그특성을밝히고유전자를클로닝하여대량발현시스템을 구축하는연구가선행된다면보다산업적인장점을갖출수 있을것으로판단된다.
그리고agarose
분해산물의NMR
분 석을통하여, GNUM08120
균주가생산하는agarase
가α-
형인지β-
형인지를규명할필요가있다.
결 론
GNUM08120
균주는agarase
생산균주로서영일만해안 가에서식하는홍조류로부터스크리닝되었다. GNUM08120
균주는16 rRNA
유전자염기서열및DNA-DNA hybridization
분석결과로부터Alteromonas macleodii
의아종으로분류되 었다. GNUM08120
균주가분비하는agarase
는CCR
효과 를나타내므로, agarase
생산증대를위해서는배양시에한 천이외의다른탄소원(
특히glucose)
은배제하는것이효과적일것으로판단되었다
. GNUM08120
의agarase
는40
oC
와pH 7.0
의조건에서효소반응을유도한다면높은효소활성을기대할수있다
.
그리고GNUM08120
의배양액을이용하여(neo)agarobiose
와(neo)agarotetraose
및(neo)agarohexaose
등의유용한(neo)agarooligosaccharide
를생산할수있을것 으로기대된다.
균주기탁
:
본연구에사용된Alteromonas macleodii subsp.
GNUM08120
균주는Korean Collection for Type Culture (KCTC)
에기탁번호KCTC23885
로기탁되었다.
염기서열등록
: GNUM08120
균주의16S rRNA
유전자 의염기서열은GenBank
에등록번호JN578475
로등록되 었다.
Acknowledgments
This work was supported by a grant from the Next-Generation Bio Green 21 Program, Rural Development Administration, Repub- lic of Korea.
References