133
목 적 :탈인산화 효소 protein tyrosine phosphatases (PTPases)를 억제시키는 orthovanadate (OVA)를 사용하여 단백질 탈인산화 억제가 MC3T3-E1 골 아세포에서 collagen 및 fibronectin (FN)의 합성과 alkaline phosphatase (ALP)의 활성도 및 골 결절 형성에 미치는 영향을 알아보고자 하였다.
재료 및 방법 :MC3T3-E1 골아세포에 OVA를 농도별, 시간별 처리한 후 collagen, noncollagen 단백질, 상대적 collagen 합성을 [3H] proline 편입 정도로, FN의 합성은 100 M OVA를 24시간 처리한 후 [35S] methionine을 편입시켜 면역침강법으로, collagen과 FN mRNA발현은 Northern blot 방법으로, ALP 활성도 측정은 분광광도계 측정법으로, 골 결절 형성은 염색법으로 분석하여 OVA를 처리하지 않은 군과 비교하였다.
결 과 :OVA는 농도 의존적으로, 단기 배양(6-30시간) 중 노출 12시간까지 collagen과 noncollagen 단백질의 합성을 증가시켰고, 상대적 교원질 합성비는 6시간에 유의하게 증가했으나 그 이후는 감소되었다. 장기 실험(5-29일)에서 OVA에 의한 collagen과 noncollagen 단백질의 합성은 증가되었으나, 상대적 collagen 합성비는 100 M농도에서 대조군과 유의한 차이는 없었고, 전 기간에 걸쳐 감소되었다. FN의 합성은 OVA에 의하여 배양액으로 분비된 FN 양에 는 변화 없이 세포층의 FN 생성을 농도 의존적으로 증가시켜 전체적으로 증가되었다. 각 단백질들의 유전자 발현은 OVA에 의하여 농도 의존적으로 colla- gen mRNA는 감소되었고, FN mRNA는 증가되었다. ALP 활성도는 OVA의 농도가 10 및 100 M에서, 단기 및 장기 세포 배양 동안 대조군에 비하여 감소 되었다. 골 결절 형성도 OVA에 의하여 현저히 감소되었다.
결 론 :OVA에 의한 PTPases의 억제는 골 기질 단백질들인 collagen과 FN의 합성을 증가시켰으나, ALP 활성도는 감소시켰으며 그 결과 골 결절 형성도 억제되었다.
색인 단어 :단백질 인산화, Orthovanadate, 골 형성
133
Orthovanadate가 MC3T3-E1 골아세포의 Collagen 및 Fibronectin 합성과 Alkaline Phosphatase 활성도에 미치는 영향
김신윤∙인주철∙최제용*∙안종철�
경북대학교 의과대학 정형외과학교실, 생화학교실*, 영남대학교 의과대학 정형외과학교실�
133 133 통신저자 : 김 신 윤
대구광역시 중구 삼덕동 2가 50 경북대학교병원 정형외과
TEL: 053-420-5635∙FAX: 053-422-6605 E-mail: [email protected]
Effect of Orthovanadate on Collagen and Fibronectin Synthesis, and Alkaline Phosphatase Activity in MC3E3-E1 Osteoblast cells
Shin-Yoon Kim, M.D., Joo Chul Ihn, M.D., Je Yong Choi, D.D.S., PhD*., and Jong Chul Ahn, M.D.�
Department of Orthopedic Surgery, Department of Biochemistry*, School of Medicine, Kyungpook National University, Daegu;
Department of Orthopedic Surgery, School of Medicine, Yeungnam National University�, Daegu, Korea
Purpose :The purpose:of this study was to know the effect of inhibition of protein dephosphorylation on the synthesis of collagen and fibronectin (FN), alkaline phosphatase (ALP) activity, and the formation of bone nodule in MC3T3-E1 osteoblasts using orthovanadate (OVA) which is a potent protein tyrosine phosphatase (PTPases) inhibitor.
Materilas and Methods :The synthesis of collagen, noncollagenous protein (NCP), and percent collagen in MC3T3-E1osteoblasts with or without OVA treatment according to concentration and time sequence was determined by incorporation of [3H]-proline, synthesis of FN by [35S] methionine incorproated immunoprecipitation after treatment with 100 M OVA for 24 hours, mRNA expression of collagen and FN by Northern blotting, activity of ALP by spectrophotometric method, and formation of bone nodule by staining method.
Results :OVA increased collagen and NCP synthesis concentration dependently, until 12 hours in short-time culture, and time depen- dently through the differentiation until 29 days, however, there was no significant effect on the percent collagen production. OVA increased percent collagen synthesis significantly at 6 hours, and decreased in a long time culture. Total FN synthesis and FN synthesis in cell layer were increased by OVA, however, FN synthesis in medium was not changed. OVA decreased collagen mRNA level dose-dependently and increased the steady-state level of FN mRNA. OVA inhibited activity of ALP in both short and long-time culture. OVA inhibited bone nodule formation in MC3T3-E1 osteoblasts.
Conclusion :These results indicate that the inhibition of PTPase by OVA increased the synthesis of collagen, FN, and decreased ALP activity and it resulted in the inhibition of bone formation in MC3T3-E1 osteoblast cells.
Key Words :Protein phosphorylation, Orthovanadate, Bone formation
Address reprint requests to Shin-Yoon Kim, M.D.
Department of Orthopaedics Surgery, Kyoung Buk University Hospital 50 2ga Samduk-dong, Chung-gu, Daegu 700-721, Korea
Tel : +82.53-420-5635, Fax : +82.53-422-6605 E-mail : [email protected]
골조직의 항상성은 파괴와 재생의 균형을 통한 국소 부위의 골개조에 의하여 유지된다2). 골아세포는 증식 및 분화되는 과정 중 골 형성에 필요한 특정 단백질들을 특정 시기에 만든다44). 골아세포의 분화에 여러 가지 성장 인자 및 호르몬들이 중요한 역할을 하며7,35) 여러 가지 성장 인자들의 작용에 의한 중요 전 사 인자들의 인산화와 탈인산화가 유전자 발현에 중요한 영향을 미친다1). 골의 형성 과정에서 인산화된 단백질들이 증가하며9) bone sialoprotein, osteopontin, glycoprotein-75 및 type I col- lagen의 amino terminal propeptide들이 골 형성에 중요한 인 산화된 단백질임이 밝혀졌다16,17).
세포는 외부 신호 전달에 대한 반응 중 하나로 단백질의 인산 화 및 탈인산화를 조절한다. 단백질의 인산화는 여러 가지 pro- tein kinases22)에 의해 일어나는데 이들은 기질 특이성이 있으 며 아미노산의 서열과 삼차 구조의 공통점을 지니고 있다26). 단 백질의 탈인산화는 phosphatases에 의해 일어나며 아미노산의 서열 및 삼차 구조상 공통점이 없다.
Orthovanadate (OVA)는 탈인산화 효소인 protein-tyrosine phosphatase (PTP ase)의 억제제이고47), Na+/K+ adnosone triphosphatase (ATPase)8), alkaline phophatase (ALP)43) 및 adenylate cyclase42) 등의 작용을 조절한다. OVA는 연골세 포에서 proteoglycan24)과 collagenase 합성11) 및 쥐 두개골의 collagen 합성6) 등 세포외 기질의 합성에도 영향을 미치며 골흡 수를 억제시킨다고 한다29).
Collagen은 골조직의 유기물 중 대부분을 차지하여, 이것이 적절히 생성되지 못하는 조건에서는 골조직의 석회화가 일어나 지 않고 ALP 활성도와 osteocalcin의 생성이 매우 낮은 것으로 나타났다37). 실제로 collagen 유전자의 돌연변이는 osteogenesis imperfecta 및 osteochondrodysplasias 등을 일으키는 것으로 알려져 있다30,36).
Fibronectine (FN)은 세포의 부착에 관여하며, 마우스에서 FN 유전자를 제거한 경우 중배엽의 형성이 잘 안되는 것으로 보아 골조직의 형성에 중요한 역할을 할 것으로 추측된다15). 골 기질 유도 간엽세포의 증식 및 분화에서 FN이 없으면 증식이 일어나지 않는다49). FN의 유전자 발현은 골아세포의 분화 과정 중 collagen과 비슷한 시기에 발현되며44), 많은 성장 인자들50), 세포 내 cyclic AMP13) 및 protein kinase C30) 등이 중요한 조 절 인자임이 밝혀졌다. 그러나 FN 합성에 있어서 PTPases의 작용기전은 아직도 잘 밝혀지지 않고 있다.
ALP는 세포막에 붙어 있는 당단백질로 여러가지 isozyme 형태로 존재한다20). 석회화시에 ALP가 국소적으로 인의 농도 를 증가시켜 미네랄 침착을 증가시킨다고 하는 가설40)이 제시된 후, ALP를 억제하는 levamisole이 분화 과정에 있는 골아세포 의 ALP를 차단하면 석회화가 일어나지 않고 석회화가 시작된 후 차단하면 효과 없는 것으로 나타나므로3) ALP는 골 형성에 아주 중요한 요소임을 알게 되었다. OVA는 쥐 두개골 기관배 양에서 ALP 활성도를 억제한다고 보고되었으나6) MC3T3-E1
골아세포에서 collagen, FN, ALP의 합성 및 골결절 형성에 대 한 효과는 아직 밝혀져 있지 않다.
본 연구에서는 MC3T3-E1 골아세포에서 OVA로 PTPase 를 억제시킨 후 collagen과 FN 합성, ALP 활성도 측정 및 골 결절 형성 정도를 분광광도법, 면역침강법, Northern blot 및 조직화학적 염색법 등으로 측정 관찰하여 단백질의 탈인산화 억 제가 골형성에 미치는 영향을 규명하고자 하였다.
재료 및 방법
실험재료Alpha-minimum essential medium ( -MEM), Dulbecco's modified Eagle's medium (DMEM), fetal bovine serum (FBS)은 GibcoBRL사(Grand Island, N.Y., USA)에서 구입 하였다. Trypsin, bovine serum albumin (BSA), ascorbic acid, -glycero-phosphate, N-ethylmaleimide, orthovanadate 및 정제된 bacterial collagenase (type VII)는 Sigma사(St. Louis, USA)에서, random primed labeling kit는 Boehringer Mann- heim사(Mannheim, Germany)에서 구입하였다. Nick-column 과 ExpressHyb solution은 Pharmacia사(Uppsala, Sweden), L- [2,3-3H] proline (12.4 Ci/mmol)과 [35S] methionine (1,000 Ci/mmol) 및 [32P]dCTP (3,000 Ci/mmol)은 Amersham 사(Arlington, VA, USA)의 제품을, 세포배양용 플라스틱 용 기는 Costar사(Cambridge, MA, USA)의 제품을 사용하였고, 기타 시약은 시판되는 특급 시약을 사용하였다.
Kodama 등27,31)에 의해 마우스의 두개골에서 만들어진 MC3- T3-E1 골아세포를 Sudo 등37)의 방법으로 10% FBS가 함유된 -MEM 배지에 배양하였고, 계대 배양시 0.001% pronase/
0.01% EDTA를 처리한 후 세포수를 1:4로 희석 배양하였다.
또한 종양 바이러스로 형질 전환시킨 K-BALB (Kirsten murine sarcoma virus transformed BALB/c 3T3) 세포를 ATCC (American Tissue Culture CollectionCo., Rockville, TN, USA) 에서 구입하여 DMEM 배지에서 배양하였다. 모든 배양액은 penicillin 100 unit/mL와 streptomycin 100 g/mL을 첨가하 여 사용하였다.
DNA 양 측정은 Labarca와 Paigen 방법32)을 사용하여 측정 하였다. 시료는 collagen 합성능 측정에 사용한 세포균질액 중 에서 100 L를 취한 것이며, 표준 용액은 송아지의 흉선 DNA (1 mg/mL)를 25, 50, 100, 500, 1,000 및 2,000 ng되게 2 N NaCl이 들어 있는 인산 완충액(0.05 M Na3PO4, pH 7.4)으로 2 mL되게 한 것이다. 시료와 표준 용액에 Hoechst 33258 (Fluka Co., USA)을 2 mL 인산 완충액으로 희석하여 최종 농도가 0.2 g/mL되게 한 다음 분광형광광도계(Model MK-2, Farrand Co. NY, USA)로 excitation 356 nm, emission 458 nm의 파장에서 측정하였다.
총단백질 및 collagen 합성능의 측정은 Peterkofsky 등39)의 방법에 따라 측정하였다. 24 well 배양 plate에 한 well 당 1×
105개씩 접종한 후 밀생 상태가 되게 하였다. 세포 분화를 유도 하면서 장기간 배양시는 3일 마다 유지 배양액(10% FBS, 50 g/mL ascorbic Acid 및 -glycero-phosphate 함유된 - MEM)을 갈아주었다. 밀생 상태가 된 후 세포에 조건 배양액 (1% FBS, 50 g/mL의 ascorbic acid 및 OVA와 2 Ci의 [3H] proline 포함된 -MEM)으로 갈아주었고 일정 기간 동 안 방사능이 편입되도록 한 후 5×collagenase 완충액(0.25 M Tris, 0.025 M CaCl2와 0.0125 M N-ethylmaleimide, 100 M PMSF, pH 7.4) 250 L씩 넣어 측정할 때까지 -20℃에 보관 하였다. 24 well plate의 각 well의 세포층을 초음파 분쇄기로 (Fisher Co. Pitturbergh, PA, USA) 30초간 마쇄한 후, 1 mL씩을 취하여 5 mL의 polypropylene 관에 넣고 BSA (2 mg/mL) 200 L와 50% trichloroacetic acid (TCA)/5 mM proline 용액 300 L를 첨가하여 잘 섞었다. 4℃에 30분 방치 한 뒤 1,000×g (Sorvall RT 6000, Albertville, MN, USA)로 5분간 원침하여 상청액을 제거하였다. 침전물에 5% TCA/1 mM proline을 1 mL 첨가하고 잘 교반한 후 같은 속도로 원침 하였다(3회 반복). 침전물을 0.2 N NaOH에 녹이고, 1 M HEPES 완충액(pH 7.2) 130 L를 첨가하여 중화시킨 후 5×collage- nase 완충액 100 L를 넣었다. 이 혼합 용액을 2개의 미량원 침관에 각각 250 L씩을 넣고 15 U의 collagenase가 함유된 또는 함유되지 않은 완충액 15 L를 각각 넣었다. 각 각을 37
℃ 수조관에 90분 동안 반응시킨 뒤 0℃로 냉각시키고, 50%
TCA/2.5% tannic acid 50 L를 첨가하여 4℃에서 30분간 방치하였다. Collagen (collagenase digestible protein: CDP) 합성능은 collagenase로 처리된 관의 상청액과 5% TCA로 한 번 씻은 상청액을 합한 것으로 계산되었다. 총 단백질량은 col- lagenase 처리하지 않은 관의 침전물을 100 L의 0.2 N NaOH에 녹여 측정하였다. Collagen합성의 상대적 비율은 총단 백질 합성에 대한 백분율로 나타내었다39). Noncollagen 단백질 합성능은 총단백질 양에서 collagen 양을 뺀 값으로 하였다.
총 RNA의 추출은 세포를 1×106 cells/100 mm plate가 되 도록 접종한 후 밀생 상태가 되도록 하였다. 약 3일이 지난 후 여러 가지 농도의 OVA가 포함된 조건 배양액(1% FBS가 함 유된 -MEM)으로 24시간 처리한 후 RNA를 Chomczynski 와 Sacchi40)의 방법으로 분리하였다. 100 mm dish의 세포층을 찬 phosphate buffer saline (PBS)로 씻은 후 변성용액(GIT:
guanidinium thiocyanate)을 각 dish당 600 L씩 넣어 세포를 완전히 녹였다. GIT 용액을 2 mL의 미량원침관에 옮기고 1/10 용량의 2 M sodium acetate (pH 4.0)를 첨가한 후 같은 양의 phenol 액을 넣었다. GIT 용액의 1/20 용량의 chloroform/
isoamylalcohol (49:1)을 넣고 10초간 강하게 교반하여 15분 동 안 얼음에 방치한 후, 미세 원심분리기로 15,000 rpm (Hanil Co., Seoul, Korea)에서 20분간 원침하여 상청액 약 650 L를 다른 미량원침관에 옮기고 같은 양의 찬 isopropanol로 섞어 -20
℃에서 1시간 이상 방치하였다. 그 후 같은 조건으로 원침한 후
침전물을 찬 알코올(75%)로 씻고, 상온에서 15분 정도 방치하 여 말린 후 물을 각각 50 L씩 넣어 녹였다. RNA양은 U.V 분광광도계(Gilford사, USA)로 260/280 nm에서 흡광도를 잰 후 양과 순도를 계산하였다. 사용된 cDNA는 Hf67741)과 pFH142) 으로 Hf677은 EcoRI으로 처리하여 1.8 kb의 cDNA insert를 확인할 수 있었고, 벡터가 pAT 153인 pFH1은 EcoRI이나 BamHI으로 처리하여 2.3 kb의 cDNA insert를 확인할 수 있 었다. cDNA insert 준비는 Sambrook 등41)의 방법으로 하였다.
Northern blot hybridization
RNA 실험과 관련된 모든 기구는 0.1% diethylphrocarbon- ate (DEPC)용액에 하룻밤 동안 담궈 두었다가 완전히 멸균 건 조시킨 후에 사용하였고, 용액은 0.01% DEPC를 처리한 재증 류수로 만들어 멸균 사용하였다. 총 RNA는 10 g, 2 L의 10
×running 완충액[0.2 M sodium morpholinopropane sulfonate (MOPS, pH 7.0), 80 mM sodium acetate, 10 mM EDTA (pH 8.0)], 3.5 L의 formaldehyde, 10 L의 formamide, 그 리고 재증류수로 총 20 L가 되도록 하여 섞은 뒤 65℃에서 15분간 방치하였다. 시료를 얼음에서 식히고 2 L의 gel load- ing 완충액(50% glycerol, 1 mM EDTA pH 8, 0.25% bro- mophenol blue, 0.25% xylene cyanol FF)을 넣어 formalde- hyde agarose gel 전기영동을 Sambrook 등41)의 방법에 따라 시행하였다.
Gel을 재증류수로 세척하여 formaldehyde를 제거하고, UV transilluminator에서 관찰하고 사진을 찍었다. Gel 내 RNA의 이동을 증진시키기 위해 0.05 N NaOH에 20분간 gel을 둔 후 재증류수로 gel을 씻고 45분간 20×SSC (3 M NaCl, 0.3 M Na3Citrate-2H2O, pH 7.0) 용액 중에 두었다. Nylon membrane 을 물에 완전히 적시고, 20×SSC에 5분간 담근 후 agarose gel 위를 덮어 통상적인 방법으로 RNA를 transfer 하였다. Nylon membrane을 6×SSC에 5분 담그고 상온에서 30분간 말렸다.
3 MM (Whatman International Ltd., Maidstone, England) 종이 사이에 nylon membrane를 넣고 80℃ 건조기에서 2시간 동안 진공 건조시킨 후 hybridization을 시행할 때까지 dessica- tor에 보관하였다.
cDNA Labeling은 Feinberg 및 Vogelstein의 방법44)을 이용 한 random primed DNA labeling kit를 사용하여 다음과 같이 하였다. 500 L 용량의 미량 원침관에 변성시킨 25 ng의 c- DNA, 반응 혼합액 2 L, 5 L의[ -32P] dCTP 그리고 klenow 효소(2 units/ L) 1 L을 넣고 최종 용량이 20 L 되도록 재증류수로 채운 다음 37℃에서 30분간 반응시킨 후 0.5 M EDTA (pH 8.0) 1 L를 넣어 반응을 중지시켰다. 이 반 응 액을 Nick-column을 이용하여 gel filtration chromatogra- phy를 시행하여 label된 cDNA와 유리[ -322P] dCTP를 분리 하였다. Nylon membrane과 제조된 probe (1×109s cpm/g cDNA)를 사용하여 ExpressHyb solution으로 prehybridization
및 hybridization을 시행하였다. Nylon membrane의 세척은 200 mL의 0.1% SDS가 포함된 2×SSC 용액으로 10분씩 3회 한 후 500 mL의 0.1% SDS가 포함된 1×SSC 용액으로 65℃
에서 15분마다 nylon membrane을 뒤집어 주면서 45분 동안 세척하였다. Nylon membrane을 3 MM 종이 위에서 건조시켜 랩으로 싼 후 intensifying screen이 붙은 X-ray film cassette 에 넣고, -70℃에서 며칠 둔 후 현상하였다.
FN 합성능 측정은 세포를 5×104개로 35 mm 배양 용기에 서 3일간 배양하여 밀생 상태가 되게 하였다. Labeling은 -MEM 및 DMEM에 투석한 1% FBS이 든 배양액에 OVA를 넣고 마 지막 3시간 동안 labeling 배양액[35S] methionine이 없는 MEM 에 L-lysine (72.5 g/mL), L-leucine (152 g/mL), [35S]
methionine 30 Ci, 투석한 FBS을 넣어 1% 되게 한 것] 1 mL 로 갈아넣었다. 시료 처리는 Grover와 Adamson의 방법18)을 변 형하여 시행하였다. Labeling후 배양액은 phenylmethyl-sulfonyl fluoride (PMSF)/EDTA를 넣어 각각 1 mM 되게 한 후 -20℃ 에 보관하였다. 세포층은 얼음 위에서 1 mL lysis 완충액(0.01 M Tris-HCl, pH 7.4, 0.15 M NaCl, 0.02% NaN3, 1% Non- idet P-40, 1% sodium deoxycholate, 0.05% sodium dodesyl sulfate, 1 mM EDTA, 1 mM PMSF)을 넣어 미량원침관에 세포를 모았다. [35S] methionine이 단백질에 편입된 정도를 보 기 위하여 항생제 paper disk에 4 L 놓은 후 찬 TCA (5%) 로 5분간 담궜다. 다시 뜨거운 TCA (10%)로 5분간 담궈 me- thionine tRNA를 제거한 후 알코올, 알코올/ether (3:1) 및 ether 순으로 씻고 말린 다음 scintillation vial에 넣어 -counter 로 측정하였다. 시료가 5×106cpm 되도록 희석 완충액[0.01 M Tris-HCl, pH 7.4, 0.15 M NaCl, 0.02% NaN3, 1% Nonidet P-40, 1 mM EDTA, 1 mM PMSF]으로 희석하여 1 mL되 게 하고 여기에 goat anti-mouse FN antibodies (KPL, USA) 을 3 g 넣어 4℃에서 한 두 시간 두었다. Immune complex를 3 L의 protein A-Sepharose 4B (50% v/v slurry, Phama- cia, Uppsala, Sweden)를 넣어 1시간 동안 흔들어 주었으며, bead를 원침하여 수거한 후 희석 완충액으로 3번 씻었다. 마지 막으로 30 L SDS 시료 완충액을 넣어 3분간 끓인 후 6s%
gel에 150 V/45 min로 전기영동 하였다. 전기영동 후 순서대 로 고정액(30% methanol/10% acetic acid)에 1시간 이상, EN3HANCE (NEN, USA) 용액에 1시간, 물에 30분간 담근 후 70℃로 말려 자가 방사법을 실시하였다.
ALP 활성도 측정은 세포를 5×104 cells/cm2으로 24 well 배양용기에 심은 후 매 3일 마다 배양액(50 g/mL ascorbic acid와 10 mM -glycerophosphate 포함)을 갈아주었다. 세 포층의 ALP 활성도는 Bessay 등4)의 방법을 이용하여 측정하 였다. 조건을 준 24 well plate의 배양액을 제거하고 찬 PBS로 3번 씻은 후 측정할 때까지 -20℃에 보관하였다. 얼음 위에서 기질(PNPP: Sodium paranitrophenyl-2-phosphate)을 준비 하고 세포층에 완충액(0.02% Nonident P-40)을 1 mL 넣어
30초간 초음파 마쇄기로 마쇄한 다음 300 L을 취하여 ALP 의 활성도를 측정하였고 200 mL을 취하여 단백질의 양을 측정 하였다33). 표준용액으로는 paranitrophenol을 30 nmol까지 되 도록 넣고 37℃에서 30분간 반응시킨 뒤 1 N의 NaOH 250 L 를 넣어 반응을 중단시켰다. 이것을 410 nm에서 분광광도계 (Gilford, San Diego, CA, USA)로 흡광도를 측정하였고, 그 활성치는 U/min/mg protein 단위로 나타내었다.
골결절 형성은 세포를 5×104 cells/cm2으로 6 well 배양 용 기에 심은 후 매 3일 마다 배양액(50 g/mL ascorbic acid와 10 mM -glycerophosphate 포함)을 갈아 준 후 Dahl 방법12) 으로 염색하였다. 6 well 배양용기를 찬 PBS로 씻은 후 상온에 서 Alizarin Red S 용액(0.1%)을 1시간 동안 담근 후 light green SF용액(0.1%)으로 다시 30분간 염색하였다. 그 후 acetic acid (1%)로 즉시 씻은 후 알콜로 탈색시켜 광학 현미경으로 관찰하였다.
실험 값은 3-6개의 시료에서 얻은 성적을 평균±표준 편차로 하였다. 그리고 유의성 판정은 student's t-test로 검정하였다.
결 과
OVA의 농도에 따른 noncollagen 단백질, collagen 및 상 대적collagen 합성에 미치는 영향
OVA는 10 M 및 100 M에서 각각 7.2% 및 42% 정도 noncollagen 단백질(NCP)의 합성을 증가시켰고, collagen (CDP)의 합성을 10 M 및 100 M에서 통계적으로 유의하 게 증가시켰다. 상대적 collagen (percent collagen) 합성에는 유의한 증감이 없었으나 100 M OVA에서는 다소 감소하였 다(Fig. 1). 높은 농도의 OVA (1 M)에서는 세포에 독작용을 나타내어 noncollagen 단백질 및 collagen의 합성이 현저히 감 소되어 이 후 단일 농도 실험에서는 100 M OVA로 하였다.
Noncollagen 단백질, collagen 및 상대적 collagen 합성에 대 한 OVA의 효과가 비교적 짧은 시간 처리한 경우에 어떻게 변 하는지 알아보고자 3일간 배양한 세포들을 100 M OVA를 6, 12, 24, 30시간 각각 처리한 후 마지막 3시간 동안 [3H] proline 을 편입시켰다. OVA는 12시간까지 noncollagen 단백질와 col- lagen의 합성을 유의하게 증가시켰으나 이 후에는 증가 효과가 감소되었다. 상대적 collagen 합성은 초기 6시간 동안 유의하게 증가되었으나 그 이 후에는 차이가 없었다(Fig. 2).
MC3T3-E1 골아세포 분화 중 OVA에 의한 noncollagen 단백질, collagen 및 상대적collagen합성에 미치는 영향 세포 분화 후 5, 11, 17, 23, 29일째에 각각 100 M OVA 를 24시간 처리한 결과 OVA는 29일째 collagen을 제외하고 전 기간에 걸쳐 noncollagen 단백질과 collagen의 합성을 통계적으 로 유의하게 증가시켰다. 상대적 collagen의 합성은 전 기간에 걸쳐 OVA에 의하여 감소되었고 5일 및 29일째는 통계적으로 유의하게 감소하였다(Fig. 3).
FN합성에 대한 OVA의 효과
밀생 상태의 세포에 100 M OVA를 24시간 처리한 후 마 지막 3시간 동안 [35S] methionine을 편입시켜 면역침강법으로
FN의 합성을 측정하였다. FN의 합성은 OVA에 농도 의존적으 로 증가되었다. K-BALB 섬유아세포에서는 OVA에 대한 FN 의 합성에 변화가 없었다(Fig. 4). 이러한 사실은 OVA에 대한
*
** **
*
Fig. 3.Effect of orthovanadate (OVA) on noncollagen protein (NCP), collagenass digestible protein (CDP) and percent collagen syn- thesis during MC3T3-E1 cells differentiation. Cells were cultured in -MEM containing 1% FBS and 50 g/mL of ascorbic acid with (■) or without (□)100 M OVA for 24 hours. *p<0.05,**p<0.01 compared with control.
8 7 6 5 4 3 2 1 0
Percent collagen (%)
0 5 10 15 20 25 30 35
6 5 4 3 2 1 0
CDP (dpm/ng of DNA)
0 5 10 15 20 25 30 35
NCP (dpm/ng of DNA)
0 5 10 15 20 25 30 35 20
16
12
8
4
0
Hours Hours Hours
■ ■
■ ■
■
■ ■
■
■
■ ■
■
■** ■** ■
** **
*
*
Fig. 1.Dose response effect of orthovanadate (OVA) on noncollagen protein (NCP), collagenase digestible protein (CDP) and percent collagen synthesis in MC3T3-E1 cells. Cells were cultured in -MEM containing 1% FBS, 50 g/mL ascorbic acid and OVA for 24 hours.
*p<0.05,**p<0.01 compared with control.
A B C
7 6 5 4 3 2 1 0
NCP (dpm/ng of DNA)
0 10 100
5
4 3
2
1 0
CDP (dpm/ng of DNA)
0 10 100
Percent collagen (%)
0 10 100
18 16 14 12 10 8 6 4 2 0
Orthovanadate ( M) Orthovanadate ( M) Orthovanadate ( M)
**
** *
Fig. 2.Time-response effect of orthovanadate (OVA) on noncollagen protein (NCP), collagenass digestible protein (CDP) and per- cent collagen synthesis in MC3T3-E1 cells. Cells were cultured in -MEM containing 1% FBS, 50 g/mL ascorbic acid with (●) or without (○) 100 M OVA for each indicated time. *p<0.05, **p<0.01 compared with control.
10 8 6 4 2 0
Percent collagen (%)
0 6 12 24 30
4
3
2
1
0
CDP (dpm/ng of DNA)
0 6 12 24 30
NCP (dpm/ng of DNA)
0 6 12 24 30
8
6
4
2
0
Hours Hours Hours
**
* *
●
●
●
●○
●
●
● ●
●
●
●○ ●○
*
*
FN의 합성 조절이 세포 특이적인 현상임을 나타낸다. OVA에 의한 FN의 합성 증가가 FN의 분비에 어떤 영향을 미치는지를 관찰하기 위하여 세포층과 배양액에 유리된 부분을 나누어 측정 한 결과, OVA는 세포층의 FN합성을 증가시켰고 배양액의 FN 치는 변화가 없어 FN의 분비에는 영향을 주지 않고 세포층의 FN 생성을 증가시키는 것으로 나타났다(Fig. 5).
Procollagen 1(I)및FN mRNA의 발현에 대한OVA의 효과
1-100 M OVA를 24시간 처리한 후 총 RNA를 추출하여 Northern blot을 한 결과 Procollagen 1(I) mRNA는 OVA의 농도 의존적으로 감소되었고 반대로 FN mRNA는 증가되었다 (Fig. 6).
농도 및 시간에 따른ALP활성도에 대한 OVA의 효과 밀생상태의 세포에 1-100 M OVA를 24시간 처리 후 세포 층의 ALP 활성도를 측정한 결과 ALP 활성도는 농도가 증가 함에 따라 감소하였으며 10 M 및 100 M OVA 처리 군에 서는 처리하지 않은 군 보다 유의하게 감소하였다(Fig. 7). 분 화시기에 따른 OVA의 효과를 알아보고자 세포 배양 5, 11, 17, 23, 29일째 100 M OVA를 24시간 처리한 결과 17일째를 제 외하고 전 기간에 걸쳐 OVA는 ALP 활성도를 감소시켰다 (Fig. 8).
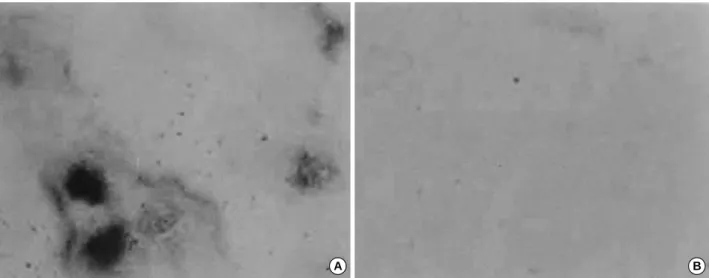
분화 과정 중OVA가 골결절 형성에 미치는 영향 매 3일 마다 배양액 교환시 100 M OVA를 24시간 처리한 후, 칼슘 침착을 알 수 있는 Alizalin Red S염색을 시행한 결과 15일 후 OVA를 처리한 군에서는 골결절이 생기지 않았으나 처리하지 않은 대조군에서는 많은 골결절이 형성되어 있었다 (Fig. 9).
고 찰
본 연구는 OVA에 의하여 MC3T3-E1 세포에서 골 형성에 중요한 collagen과 noncollagen 단백질의 합성이 증가하였고 골 결절의 형성이 억제되었다. 쥐 두개골의 기관배양6)에서도 본 연 구와 유사하게 collagen과 noncollagen 단백질의 합성이 모두 증가되었다. 그러나 이러한 단백질들의 증가가 Canalis6)의 주장 과는 다르게 골 형성의 증가로 나타나지 않았다. 이것은 이러한 단백질들의 비율이나 양적 균형이 골 형성에 중요하다는 것을 나타낸다. Collagen과 FN의 합성을 증가시키는 transforming growth factor- 를 쥐 두개골에서 일차 배양한 골아세포의 분 화 초기에 처리하면 골 형성이 억제되는 것으로 알려져5) 단순 한 collagen 및 noncollagen 단백질들의 합성능의 증가가 골 형 성의 촉진으로 연결되지 않음을 나타내는 좋은 예가 될 수 있다.
Collagen과 noncollagen 단백질의 합성이 100 M OVA에 의 하여 기관배양에서는 감소되었으나 MC3T3-E1 골아세포에서 는 모두 증가되었다. 두 system 간의 이러한 차이점에 대한 기 전은 아직 잘 모르지만 적어도 부분적으로 실험에 사용된 세포 Fig. 5. Effect of orthova- nadate (OVA) on fibr- onectin (FN) synthesis in cell and cell culture medium of MC3T3-E1 cells. Cells were grown to confluence and treat- ed with 100 M of OVA for 24 hours. After 3 ho- urs labeling samples were separate into cel- lular layer and medium.
FN
Cell
0 OVA 0 OVA
Media
250 kDa
0 1 10 100
MC3T3-E1 K-BALB
OVA ( M)
FN
0 100
Fig. 6.Effect of orthovanadate (OVA) on fibronectin (FN) and pro- collagen (Procoll) mRNA levels in MC3T3-E1 cells. Cells were grown to confluence and treated with 100 M of OVA for 24 hours.
FN
0 1 10 100 ( M)
Procoll
OVA
7.9 kb
5.7 kb 4.7 kb Fig. 4.Effect of orthovanadate (OVA) on fibronectin (FN) syn- thesis by immunoprecipitation in MC3T3-E1 cells. Cells were grown to confluence and treated with various dose (1-100 M) of OVA for 24 hours. Labeling medium was changed for the last 3 hours.
들의 차이로 인한 것이라 생각해 볼 수 있다. 즉 기관배양은 같 은 종류로 구성된 MC3T3-E1 골아세포와는 달리 여러 가지 세 포들로 구성되어 있어 OVA에 대한 작용이 다를 수 있다.
MC3T3-E1 골아세포에서 총단백질의 합성에 대한 collagen의 상대적 합성은 농도에 따라 뚜렷한 특성을 나타내지 않았다. 단 지 10 M의 OVA에서는 약간 증가하는 경향을 나타내었고 100 M의 OVA에서는 감소하는 경향을 나타내었으나 통계적 으로 유의한 차이는 없었다. OVA에 의하여 procollagen 1(I) mRNA 양은 농도-의존적으로 감소하여 단백질 수준인 상대적 collagen의 합성과 다르게 나타났다. 이러한 차이점은 현재로서 잘 알 수 없으나 OVA가 collagen 전사 단계나 mRNA의 안정 도에 기인된 결과로 추정해 볼 수 있다. Canalis6)에 의하면 organ culture에서도 OVA를 24시간 처리하면 상대적 collagen
합성이 억제된다고 하였다. 세포가 분화함에 따라 OVA가 noncollagen 단백질, collagen 및 상대적 collagen 합성에 미치 는 영향은 분화정도에 따라 큰 차이가 없었으며, 이는 세포가 분화를 하는 과정에서도 PTPases의 작용이 계속된다는 것을 암시하였다. 이는 세포 밀생 정도가 높을 수록 PTPase의 활성 도가 증가된다는 사실과 일치하였다38). 본 연구에서 고농도인 1 M의 OVA에서는 noncollagen 단백질 및 collagen의 합성이 현 저히 감소되었으나 이는 OVA의 세포 독작용 때문이라 생각되 며 그 기전은 잘 모르지만 Cantley 등8)은 Na, K-ATPase가 억제되기 때문이라고 하였다.
골기질 단백질의 합성에 PTPases가 중요한 역할을 하는 또 다른 증거로는 OVA로 PTPases를 억제하면 FN의 합성이 증 가되나 분비에는 큰 영향이 없음을 나타내어 PTPases의 활성 Alkaline phosphatase activity (nmol/min/mg of protein)
0.7 0.6 0.5 0.4 0.3 0.2 0.1 0.0
0 1 10 100
Fig. 7.Effect of orthovanadate (OVA) on ALP activity in MC3T3- E1 cells. Cells were cultured in -MEM containing 1% FBS with various dose (1-100 M) of OVA for 24 hours. *p<0.01 com- pared with control.
Orthovanadate ( M)
Alkaline phosphatase activity (nmol/min/mg of protein) 7 6 5 4 3 2 1 0
0 5 10 15 20 25 30 35
Fig. 8.Effect of orthovanadate (OVA) on activities of alkaline phosphatase during MC3T3-E1cells differentiation. Cells were cultured in -MEM containing 1% FBS and 50 g/mL of ascorbic acid with (□) or without (■) 100 M of OVA for 24 hours. *p<0.01 compared with control.
days
■
■
■
■ ■
□
□
□
□ □
*
*
A B
Fig. 9.Effect of OVA on the formation of bone nodules during the MC3T3-E1 cells differentiation. Cells were cultured in -MEM containing 10% FBS, 10 mM of -glycerolphosphate and 50 g/mL of ascorbic acid and medium was changed every 3 days with (B) or without (A) 100 M of OVA. Phase contrast micrographs (×40) by Alizarin Red S staining after 15 days.
*
*
은 collagen과는 달리 FN 합성의 억제로 작용을 한다는 것을 의미한다. 아직 정확한 기전은 잘 모르지만 FN 유전자 발현과 관계 있는 전사 인자들의 인산화 정도가 FN 유전자 발현에 어 떤 영향을 미칠 것으로 생각되며, 다른 세포에서 serine/threo- nine phosphatase (PPase)가 FN 유전자 발현을 억제하는 것 으로 알려져 이를 뒷받침한다19).
Protein phosphatase의 억제재로서 OVA는 PTPases를, okadaic acid은 PPases를 억제하여 FN 유전자 발현을 증가시 키나, OVA에 의하여 정상 세포는 암세포로 전환되고 okadaic acid19)에 의하여 암세포는 정상 세포로 형태가 변형되는 것으로 알려져 있다. 골아세포에서 OVA에 의하여 FN의 합성이 증가 되었을 때 골 형성에 어떤 영향을 미치는지 아직 잘 모르나 본 연구의 결과로 보아 이러한 비정상적인 FN합성 증가는 골결절 형성 억제와 부분적으로 연관이 있을 것으로 생각되며, 이 부분 에 대해서는 추가적인 연구가 필요하다. OVA가 MC3T3-E1 세포의 분화 정도와 상관없이 ALP의 활성도를 억제한다는 사 실은 PTPases의 활성이 또 다른 표지자인 ALP의 활성도에 영 향을 미친다는 것을 나타낸다. 이러한 OVA의 ALP 활성 억제 사실은 쥐 두개골의 기관배양6)이나 근육8)에서도 잘 알려져 있 다. 이러한 단백질들의 합성 및 활성도의 변화는 골 형성 지표 인 골결절 형성을 억제할 수 있는 또 다른 하나의 근거가 될 수 있다.
결 론
본 연구 결과 PTPases의 강력한 억제재인 OVA는 MC3T3- E1 골아세포에서 FN 유전자의 발현을 증가시키고 collagen과 noncollagen 단백질 합성의 증가 및 ALP 활성도를 감소시켰고 이러한 결과가 골 형성 지표인 골결절 형성의 억제를 초래하였 다. 이는 세포내 PTPases의 활성도를 조절함으로써 골 형성을 조절할 수 있을 가능성을 시사하는 것으로 보인다.
참고문헌
1. Abate C, Baker SJ, Lees-Miller SP, Anderson CW, Marshak DR and Curran T: Dimerization and DNA binding alter phosphorylation of fos and jun. Proc Natl Acad Sci USA, 90: 6766-6770, 1993.
2. Baron R, Vignery A and Horowitz M: Lymphocytes, macrophages and the regulation of bone remodeling, In: Bone and Mineral Research, Peck WJ, New York, Elsevier, 175-243, 1984 .
3. Bellows CG, Aubin JE and Heersche JNM: Initiation and progres- sion of mineralization of bone nodules formed in vitro: the role of alkaline phosphatase and organic phosphate. Bone Miner, 14: 27-40, 1991.
4. Bessay OA, Lowry OH and Brock MJ: A method for the rapid deter- mination of alkaline phosphatase with fibe cubic millimeters of serum. J Biol Chem, 164: 321-329, 1946.
5. Breen EC, Ignotz RA, McCabe L, Stein GS and Lian JB: TGF- alters growth and differentiation related gene expression in proliferating
osteoblasts in vitro, proeventing development of the mature bone pheno- type. J Cell Physiol, 160: 323-335, 1994.
6. Canalis E: Effect of sodium vanadate on deoxyribonucleic acid and pro- tein synthesis in cultured rat calvariae. Endocrinology, 11: 855-862, 1985.
7. Canalis E: Insulin like growth factors and the local regulation of bone for- mation. Bone Miner, 14: 273-276, 1993.
8. Cantley LC, Josephson L, Warner R, Yanagisawa M, Lechene C and Guidotti G: Vanadate is a potent (Na,K)-ATPase inhibitor found in ATP derived from muscle. J Biol Chem, 252: 7421-7423, 1977.
9. Chomczynski P and Sacchi N: Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction. Anal Biochem, 162: 156-159, 1987.
10. Chu ML, Myers JC, Bernard MP, Ding JF and Ramirez F: Cloning and characterization of five overlapping cDNAs specific for the human pro 1(I) collagen chain. Nucleic Acids Res, 10: 5925-5934, 1982.
11. Cruz TF, Mills G, Pritzker KPH and Kandel RA: Inverse correlation between tyrosine phosphorylation and collagenase production in chondro- cytes. Biochem J, 269: 717-721, 1990.
12. Dahl LK: A simple and sensitive histochemical method for calcium. Proc Soc Exp Biol Med, 80: 474-479, 1952.
13. Dean DC, Blakeley MS, Newby RF, Ghazal P, Hennighausen L and Bourgeois S: Forskolin inducibility and tissue-specific expression of the fibronectin promoter. Mol Cell Biol, 9: 1498-1506, 1989.
14. Feinberg AP and Vogelstein B: A technique for radiolabeling DNA restriction endonuclease fragments to high specific activity. Anal Biochem, 132: 6-13, 1983.
15. George EL, Georges-Labouesse EN, Patel-King RS, Rayburn H and Hynes RO: Defects in mesoderm, neural tube and vascular develop- ment in mouse embryos lacking fibronectin. Development, 119: 1079- 1091, 1993.
16. Gerstenfeld LC, Gotoh Y, Mckee MD, Nanci A, Landis WJ and Glimcher MJ: Expression and ultrastructural immunolocalization of a major 66 kDa phosphoprotein synthesized by chicken osteoblasts during mineralization in vitro. Anal Rec, 228: 93-103, 1990.
17. Glimcher MJ: Mechanism of calcification: role of collagen fibrils and col- lagen-phosphoprotein complexes in vitro and in vivo. Anal Rec, 224: 139- 153, 1989.
18. Grover A and Adamson ED: Roles of extracellular matrix components in differentiating teratocarcinoma cells. J Biol Chem, 260: 1225-1228, 1985.
19. Gupta RW, Joseph CK and Foster DA: V-src-induced transformation is inhibited by okadaic acid. Biochem Biophy Res Commun, 196: 320-327, 1993.
20. Harris H: The human alkaline phosphatases: what we know and what we don't know. Clin Chem Acta, 186: 137-150, 1989.
21. Hunter T and Karin M: The regulation of transcription by phosphory- lation. Cell, 70: 375-387, 1992.
22. Hunter T: Protein kinase classification. Method Enzymol, 200: 3-37, 1991.
23. Karin M and Smeal T: Control of transcription factors by signal trans- duction pathways: the beginning of the end. Trends Biochem Sci, 17: 418- 422, 1992.
24. Kato Y, Iwamoto M, Koike T and Suzuki F: Effect of vanadate on cartilage-matrix proteoglycan synthesis in rabbit costal chondrocyte cul- tures. J Cell Biol, 104: 301-319, 1987.
25. Klarlund JK: Transformation of cells by an inhibitor of phosphatases act- ing on phosphotyrosine in proteins. Cell, 41: 707-717, 1985.
26. Knighton D, Zheng J, Eyck L, et al: Crystal structure of the catalytic subunit of cyclic adenosine monophosphate-dependent protein kinase. Sci- ence, 253: 407-414, 1991.
27. Kodama H, Amagai Y, Sudo H, Kasai S and Yamamoto S: Estab- lishment of a clonal osteogenic cell line from newborn mouse calvaria. Jpn J Oral Biol, 23: 899-901, 1981.
28. Kornblihtt AR, Vibe-Pedersen K and Baralle FE: Isolation and characterization of c-DNA clones for human and bovine fibronectins. Proc Natl Acad Sci, 80: 3218-3222, 1983.
29. Krieger NS and Tashjian AH: Inhibition of stimulated bone resorption by vanadate. Endocrinology, 113: 324-328, 1983.
30. Kuivaniemi H, Tromp G and Prockop DJ: Mutations in collagen genes: causes of rare and some common diseases in humans. FASEB J, 5:
2052-2060, 1991.
31. Kumegawa M, Hiramatsu M, Hatakeyama K, et al: Effects of epi- dermal growth factor on osteoblastic cells in vitro. Calcif Tissue Int, 35:
542-548, 1983.
32. Labarca C and Paigen K: A simple, rapid, and sensitive DNA assay procedure. Anal Biochem, 102: 344-352, 1980.
33. Lowry OB, Rosenbrough MJ, Farr AL and Rebar RW: Protein mea- surement with folin phenol reagent. J Biol Chem, 193: 255-260, 1951.
34. Meek DW and Street AJ: Nuclear protein phosphorylation and growth control. Biochem J, 287: 1-15, 1992.
35. Mundy GR, Boyce B, Hughes D, et al: The effects of cytokines and growth factors on osteoblastic cells. Bone, 17: 71S-75S, 1995.
36. Olsen BR: Mutations in collagen genes resulting in metaphyseal and epiphyseal dysplasias. Bone, 17: 45S-49S, 1995.
37. Owen TA, Aronow M, Shahoub V, et al: Progressive development of the rat osteoblast phenotype in vitro: reciprocal relationships in expression of genes associated with osteoblast proliferation and differentiation during formation of the bone extracellular matrix. J Cell Physiol, 143: 420-430, 1990.
38. Pallen CJ and Tong PH: Elevation of membrane tyrosine phosphatase activity in density-dependent growth-arrested fibroblasts. Proc Natl Acad Sci, 88: 6996-7000, 1991.
39. Peterkofsky B, Chojkier M and Bateman J: Determination of colla- gen synthesis in tissue and cell culture systems, In: Immunochemistry of the extracellular matrix, Vol. II, CRC Press, Boca Raton, Florida, 1982, pp.19-47.
40. Robinson B: The possible significance of hexose phosphoric esters in ossi- fication. Biochem J, 17: 286-293, 1923.
41. Sambrook J, Fritsch EF and Maniatis T: Molecular cloning: a labora- tory manual, Cold spring harbor laboratory, New York, 46-52, 1989.
42. Schwabe U, Puchstein C, Hannemann H and Sochting E: Activa- tion of adenylate cyclase by vanadate. Nature, 277: 143-145, 1979.
43. Seargeant LE and Stinson RA: Inhibition of human alkaline phos- phatase by vanadate. Biochem J, 181: 247-250, 1979.
44. Stein GS, Lian JB, Bortell TA and Owen A: Relationship of cell growth to the regulation of tissue-specific gene expression during osteoblast differ- entiation. FASEB J, 4: 3111-3123, 1991.
45. Studer RK, Craven PA and DeRubertis FR: Role for protein kinase C in the mediation of increased fibronectin accumulation by mesangial cells grown in high-glucose medium. Diabetes, 42: 118-126, 1993.
46. Sudo H, Kodama H, Amagai Y, Yamamoto S and Kasai S: In vitro differentiation and calcification in a new clonal osteogenic cell line derived from newborn mouse calvaria. J Cell Biol, 96: 191-196, 1983.
47. Swarup G, Speeg KV, Jr Cohen S and Grabers DL: Phosphotyro- syl-protein phosphatase of TCRC-2 cells. J Biol Chem, 257: 7298-7301, 1982.
48. Weiss RE and Reddi AH: Role of fibronectin in collagenous matrix- induced mesenchymal cell proliferatio and differentiation in vivo. Exp Cell Res, 133: 247-254, 1981.
49. Yamada KM: Fibronectins: structure, functions and receptors. Curr Opin Cell Biol, 1: 956-963, 1989